
Abstract
Polyelectrolyte multilayer films have been suggested as tunable substrates with flexible surface properties that can modulate cell behavior. However, these films’ biological effects on neural stem/progenitor cells have rarely been studied. Herein, biomimetic multilayer films composed of hyaluronic acid and poly-L-lysine were chosen to mimic the native extracellular matrix niche of brain tissue and were evaluated for their inductive effects, without the addition of chemical factors. Because neural stem/progenitor cells are sensitive to substrate properties, it is important that this system provides control over the surface charge, and slight stiffness variations are also possible. Both of these factors affect neural stem/progenitor cell differentiation. The results showed that neural stem/progenitor cells were induced to differentiate on the poly-L-lysine/hyaluronic acid multilayer films with 0.5–4 alternating layers. In addition, the neurite outgrowth length was regulated by the surface charge of the terminal layer but did not increase with the layer number. In contrast, the quantity of differentiated neurons was enhanced slightly as the number of layers increased but was not affected by the surface charge of the terminal layer. In sum, material pairs in the form of native poly-L-lysine/hyaluronic acid films achieved important targets for neural regenerative medicine, including enhancement of the neurite outgrowth length, regulation of neuron differentiation, and the formation of a network. These extracellular matrix–mimetic poly-L-lysine/hyaluronic acid multilayer films may provide a versatile platform that could be useful for surface modification for applications in neural engineering.
Keywords
Introduction
Neural stem/progenitor cells (NSPCs), which are present in the developing and adult mammalian central nervous systems, possess self-renewal capacity and can be induced into three neural lineages: neurons, astrocytes, and oligodendrocytes.1,2 Thus, these cells are attractive candidates for the development of therapies in neural regenerative medicine. However, undesired differentiation and the low viability of NSPCs are current limitations for their use. The stem cell niche is a complex, anisotropic environment composed of a number of biophysical and biochemical factors that control cellular processes, and the neural stem cell niche regulates neurogenesis as well as the survival and regeneration of NSPCs.3–6 Stem cells tend to differentiate into distinct lineages in vitro in response to different signaling factors supplied by artificial niches.7–9 Many studies have discussed the function of chemical reagents, such as platelet-derived growth factor-BB (PDGF-BB), basic fibroblast growth factor (b-FGF), and nerve growth factor (NGF), in stimulating neurotransmitter release, cell growth, and cell differentiation.10,11 However, the independent effects of the substrate and the effects of native structure modulation on NSPC differentiation have rarely been explored.12,13
Certain protein, polypeptide, and extracellular matrix (ECM) components are generally used to enhance cell adhesion and niche fabrication.14–17 Hyaluronic acid (HA) is a major ECM component in tissues and has been identified to have important effects on cell behaviors, such as cell differentiation, cell growth, and cell migration.18–20 In addition, HA is ubiquitous and has been identified as the main component of the ECM in brain tissue.21–23 However, most cells do not preferentially adhere to substrates coated with pure HA because of its negative charge.24,25 Previous studies have used poly-D-lysine to modify HA scaffolds for applications in tissue repair. 26 Wu et al. 27 used HA-based polyelectrolyte multilayer (PEM) films to improve biocompatibility with neural cells. Because polylysine is also one of the polypeptides that have been widely used to improve neural cell adhesion and neurite outgrowth, 28 in the current study, poly-L-lysine (PLL)/HA composite materials were chosen to fabricate native multilayer films and to determine their inductive effects on NSPCs.
The alternating adsorption of polyanions and polycations onto a charged surface usually leads to the progressive formation of films that have been defined as PEM films. This electrostatic self-assembly method was first developed by Decher et al. 29 and has been extensively used in the biomedical field to fabricate ultra-thin films containing macromolecules such as proteins, 30 hormones, 31 and drugs 32 as well as films that enhance cell activity. 18 The multilayers constitute versatile tools that offer many possibilities for tuning the film properties, such as thickness, surface roughness, composition, and hydrophilicity.33,34 In addition, the layer-by-layer assembly of PEM films from natural materials has been demonstrated to be a promising approach for creating biomaterials that mimic the native ECM. 35
Previous studies have shown that most cells adhere to and grow on PEM films with higher elasticity.35,36 However, brain tissue is one of the softest tissues in the body, and many studies have demonstrated that neurons favor soft surfaces. 37 Leach et al. 38 showed that neurite extension from PC12 cells can be modulated by tunable matrices and culture substrates, including collagen and other soft substrates. 39 HA has a high water capacity and possesses a high viscoelasticity. Thus, it is believed that the stiffness of HA-based PEM films may be lower than that of other material pairs with the same number of layers, providing niches that are similar to brain tissue.
Herein, a biochemical-free system for evaluating the effect of the surface of the niche on NSPC differentiation was fabricated. In particular, native ECM-mimetic multilayer films were chosen to facilitate NSPC differentiation. In this study, the biological properties of NSPCs cultured on sequential PLL/HA multilayer films that were evaluated included the neurite outgrowth length, the differentiation lineage, and the synapse functionality of differentiated neurons. In addition, the percentage of differentiated neurons on PLL/HA multilayer films was compared with that on PLL/poly-L-glutamine acid (PLGA) multilayer films.
Materials and methods
Monitoring of PLL/HA multilayer film growth using a quartz crystal microbalance
A quartz crystal microbalance (QCM) system employing AT-cut quartz crystals, P-chip (Au-25; ANT Technology Co. Ltd., Taiwan), was used to monitor the in situ growth of multilayers. Before use, the crystals were cleaned with an O2 plasma cleaner (PDC-001-HP; Harrick Plasma, USA). The Affinity Detection System (ADS; ANT Technology Co. Ltd., Taiwan) was used to perform and record the flow injections and continuous frequency variations. A non-continuous plug was used to exchange the liquids at a flow rate of 30 µL/min. All measurements were recorded for an average of 10 samples per second.
Preparation of PLL/HA multilayer films
The preparation of PLL/HA multilayer films followed a procedure described in the literature, with modifications. 40 The physical deposition of PLL/HA multilayer films was performed using batch and static conditions. First, all polyelectrolytes were dissolved in 10 mM Tris-HCl buffer with 0.15 M NaCl at pH 7.4. All of the polyelectrolyte solutions used here were at a concentration of 1 mg/mL and were filtered through an 8-µm Millipore membrane used in cell studies. To begin the deposition, glass cover slides were immersed in PLL solution (Sigma, St. Louis, MO), kept at room temperature for 10 min, and then rinsed with 1 mL of Tris-HCl buffer for 1 min. Second, the PLL-coated slides were immersed in HA solution (Sigma, St. Louis, MO) for 10 min and again rinsed with 1 mL of Tris-HCl buffer for 1 min. Finally, PBS was used to clean the slides and remove uncoupled polypeptides. The layered materials were designated as PLL/HA (n), where n denotes the number of polyelectrolyte pairs generated by repeating the above steps. For example, n = 0.5 refers to glass-PLL only, and n = 1 refers to PLL/HA (n = 1).
Isolation and culture of cortical NSPCs
Cerebral cortical NSPCs were isolated from ED 14 to 15 Wistar rat embryos using a previously described protocol. 13 The animal work in this study was performed in strict accordance with the recommendations of the Institutional Animal Care and Use Committee at Chang Gung University (IUPAC permit number CGU13-018). Briefly, rat embryonic cerebral cortices were dissected, cut into small pieces, and mechanically triturated in cold Hank’s balanced salt solution (HBSS). After dissociation, the cells were collected by centrifugation and resuspended in serum-free Dulbecco’s modified Eagle’s medium-F12 with N2 supplementation. Purification and culture of the NSPCs were performed in T25 culture flasks (Corning, NY, USA). The cells were plated at a density of 50,000 cells/cm2, and basic fibroblast growth factor (bFGF) was added to the medium. The isolated cells were cultured in an incubator and maintained at 37℃ and 5% CO2. After 1–3 days in vitro, neurospheres formed from the proliferating cells in suspension in the medium. Subsequently, the suspended neurospheres were collected by centrifugation, and the adherent cells were discarded. The neurospheres were then subcultured at the same density in fresh medium containing bFGF in a new flask. Before seeding, the substrates were placed in 24-well tissue culture polystyrene (TCPS) plates and kept under ultraviolet light overnight for sterilization. The cells were then seeded at a density of 200 spheroids per well in 1 mL of medium/cell suspension.
Lactate dehydrogenase release assay
The cytotoxicity of the sequential PLL/HA multilayer films to NSPCs was evaluated by using a Cytotoxicity Detection Kit (Roche, Mannheim, Germany) to measure lactate dehydrogenase (LDH) release during culture. The culture medium was collected and incubated with the reaction mixture from the kit according to the manufacturer’s protocol. The LDH content was estimated by an enzyme-linked immunosorbent assay (ELISA), and the results were read by a plate reader (Synergy™ HT, Bio-Tek, USA) at an absorbance of 490 nm, with a reference wavelength of 630 nm.
Immunostaining of NSPCs on different substrates
For immunocytochemical characterization, NSPCs cultured on substrates for 5 days were fixed in ice-cold 4% paraformaldehyde in PBS for 20 min and then washed three times with PBS. After fixation, the cells were incubated with the following primary antibodies: rabbit anti-microtubule-associated protein 2 (MAP-2; 1:500 dilution; Chemicon), rabbit anti-glial fibrillary acidic protein (GFAP; 1:500 dilution; Chemicon), and anti-synapsin I (1:1000 dilution; Chemicon). The primary antibodies were diluted in PBS containing 10% bovine serum albumin and 0.3% Triton X-100 and were incubated with the cells for 2 h at room temperature. The cells were then incubated with secondary antibodies for 30 min at room temperature. The secondary antibodies used here were FITC-conjugated goat anti-mouse IgG (preabsorbed with rabbit and rat serum protein; 1:100; Chemicon), FITC-conjugated donkey anti-rabbit IgG (preabsorbed with rabbit and rat serum protein; 1:100; Chemicon), and rhodamine-conjugated goat anti-mouse IgG (preabsorbed with rabbit and rat serum protein; 1:100; Chemicon). The immunostained cells were visualized by confocal microscopy (LSM 510 META, Zeiss, Germany).
Analysis of the neurite outgrowth length
Photomicrographs were taken of random fields of view for NSPC spheres cultured on sequential PLL/HA multilayer films for 3 days. The lengths of 10–15 of the longest processes on each neurosphere were measured from the edge of the neurosphere to the tips of the processes, i.e. the end-to-end distance. The process lengths were measured by tracing the processes using ImageJ software, and the process lengths of at least 10 independent neurospheres were calculated for each substrate. The mean and standard error of the mean (SEM) were calculated for each treatment.
Analysis of the percentage of neuron/astrocyte differentiation
Immunochemistry images were collocated with standard fluorescence illumination and a digital camera for at least three random fields of view for each group after 5 days of culture. The percentage of differentiated cell phenotypes was calculated, and the number of GFAP- and MAP2-positive cells was divided by the total cell number. Data were collected from three independent experiments based on three separate cultures and analyzed using ImageJ software.
Statistical analysis
The data are presented as the means ± standard error from 4 to 6 independent experiments. The results were analyzed using Student’s t-test and the significance values were further adjusted for comparisons between multiple groups using the Bonferroni correction and the number of independent comparisons made was n = 4. Statistical significance is indicated as *p < 0.0125 and **p < 0.00025.
Results and discussion
The procedures for step-by-step PLL and HA adsorption onto glass and for NSPC seeding are illustrated in Figure 1. The formation of PLL/HA multilayer films was achieved through layer-by-layer electrostatic interactions of PLL as a polycation and HA as a polyanion. After forming the PLL/HA multilayer film structures, NSPCs were seeded and cultured on the sequential PLL/HA multilayer films at a density of 200 spheroids per well per mL. After 3 days of culture, the differentiation of the NSPCs on the PLL/HA multilayer films was investigated as in the schematic illustrations.
Schematic illustrations of layer-by-layer PLL/HA adsorption on glass and of NSPC seeding, migration, and neurite outgrowth. NSPCs attached to the surfaces and differentiated into neurons and astrocytes, without the addition of chemical reagents. HA: hyaluronic acid; NSPC: neural stem/progenitor cell; PLL: poly-L-lysine.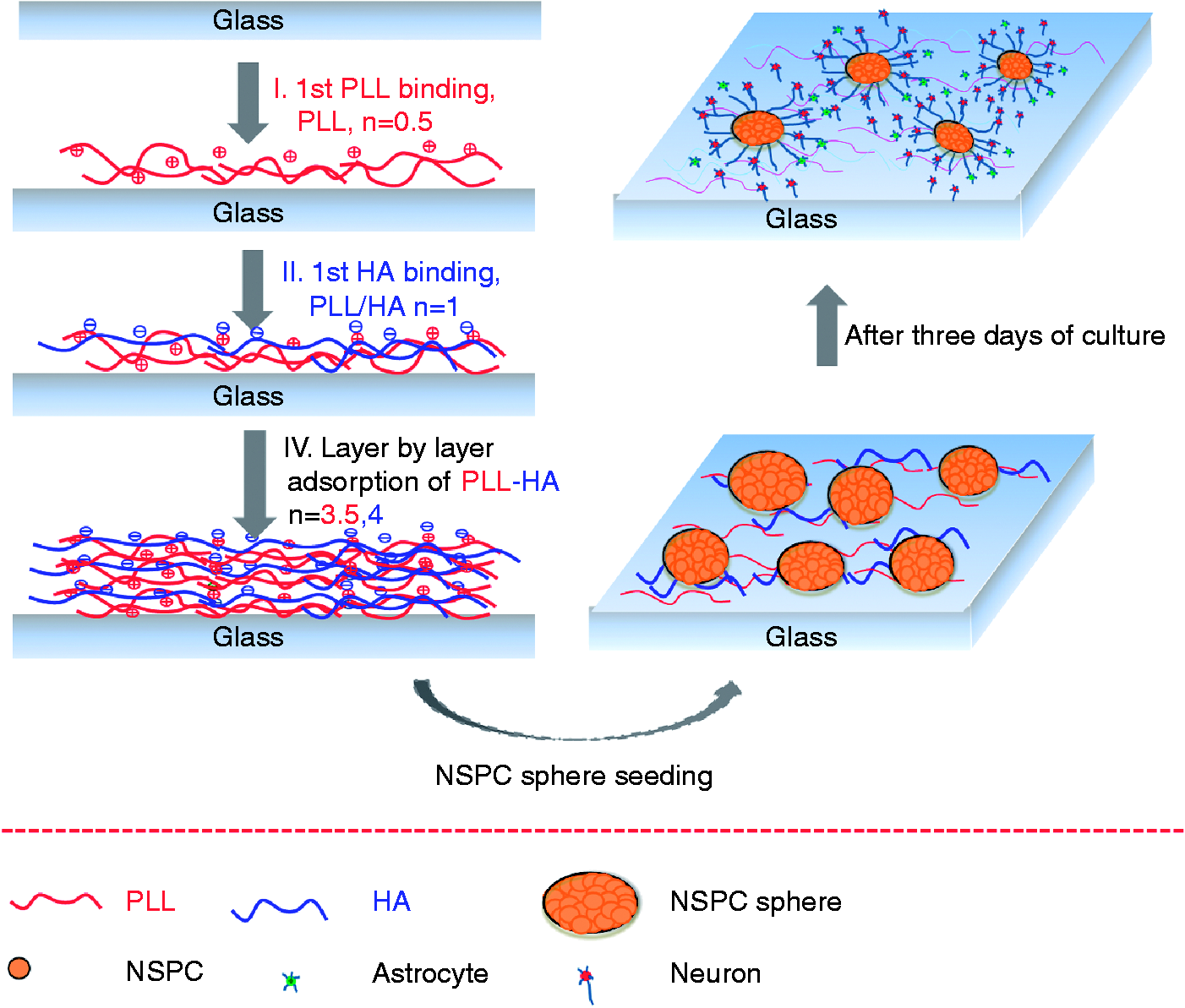
PLL/HA multilayer film growth
Figure 2 shows the QCM profiles for the formation of PLL/HA and PLL/PLGA multilayer films and illustrates the characteristics of the layer-by-layer coating process. The PEM films were built up from PLL adsorption on the Au chip treated with O2 plasma at point (I), forming a PLL film. Subsequent HA adsorption then formed a PLL/HA (n = 1) substrate at point (II). The step-by-step process shown in the QCM profile in Figure 2(a) corresponds to steps (I–II) shown in Figure 1. It is shown that the frequency decreased as the layer-by-layer deposition process progressed in both systems. However, Figure 2(a) shows that the decrease in Δf became larger as the number of layers increased, especially when the HA solution was added. In contrast, as shown in Figure 2(b), for PLGA, the decrease in Δf was smaller than for the same number of layers in the PLL/HA system. It is possible that the Δf of HA for each addition was larger than that of PLGA because of the hydration of HA, which may cause larger increases in thickness. These results are consistent with previous research showing the exponential character of the film growth.
41
Layer-by-layer PEM film growth, as monitored by QCM-D. A PLL solution was injected into the QCM chamber at point (I), as denoted by the red color, and adsorbed onto the Au chip via charge–charge interactions. Subsequently, an HA solution flowed in at point (II), as denoted by the blue color, and layer-by-layer adsorption was performed for up to four layers. (a) PLL/HA. (b) PLL/PLGA.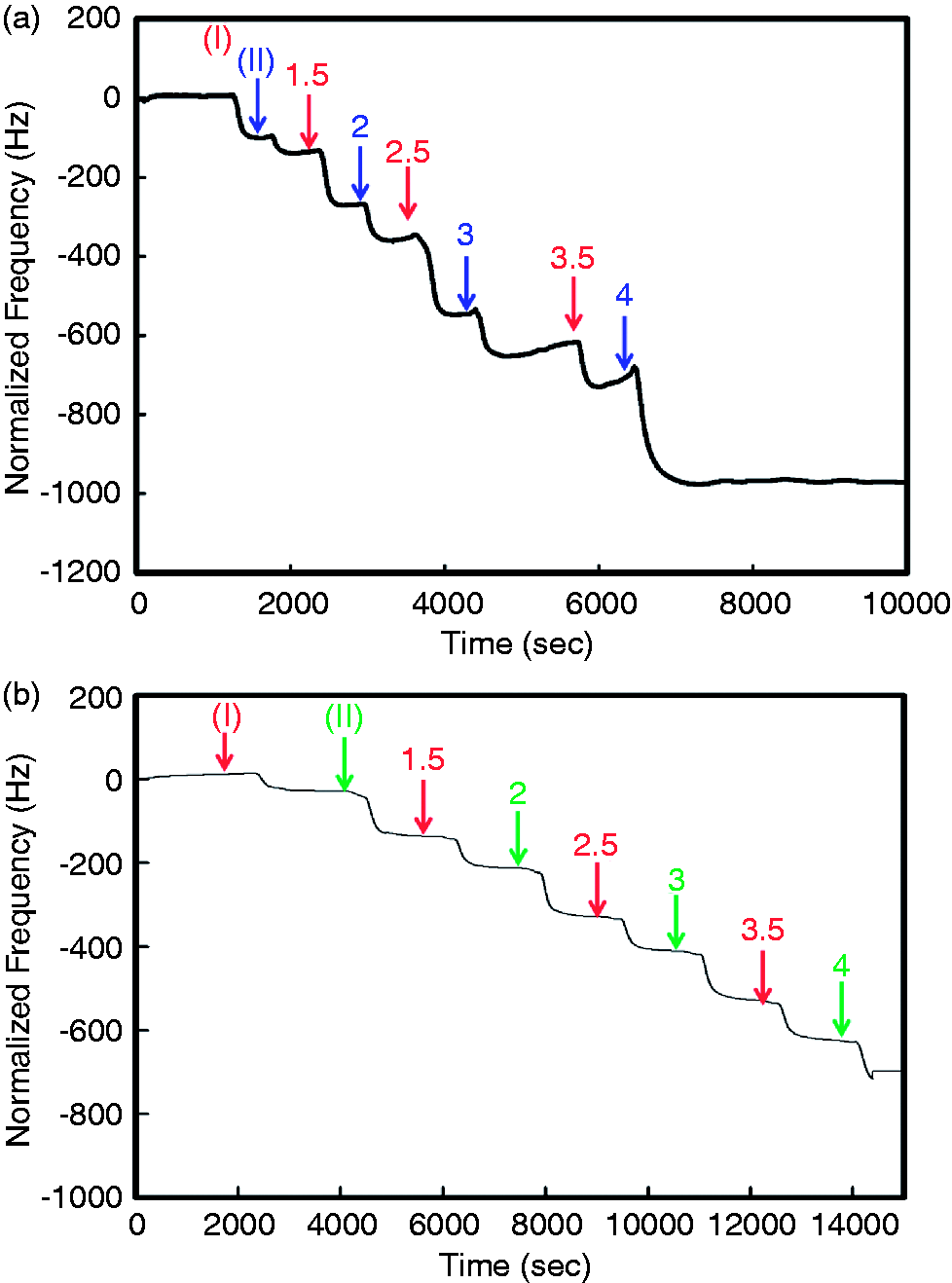
Cytotoxicity of PLL/HA multilayer films to NSPCs
Cell survival tests were performed by determining the quantity of LDH released into the culture medium because LDH is an intracellular enzyme that is released when cells are damaged. Cytotoxicity tests are critical for determining whether a given culture substrate is biocompatible with cells and suitable for cell survival. Therefore, LDH release from NSPCs cultured on sequential PLL/HA multilayer films was measured after 3 and 5 days, and the quantities are shown in Figure 3. It was found that the release levels were all slightly increased after 5 days of culture, which may have been due to the medium being serum-free. In addition, the release levels on the PEM films were slightly higher than those on TCPS at day 3, especially when PLL was the terminal layer. However, the release levels were the same for the PEM films and TCPS after 5 days of culture. The cytotoxicity test showed that sequential native PLL/HA multilayer films provide surfaces that are biocompatible with NSPCs.
LDH release by NSPC spheres cultured on TCPS, glass or sequential PLL/HA multilayer films after 3 and 5 days. The optical density corresponding to the LDH activity was read on an ELISA plate reader at 490 nm, with a reference wavelength of 630 nm. HA: hyaluronic acid; PLL: poly-L-lysine; TCPS: tissue culture polystyrene.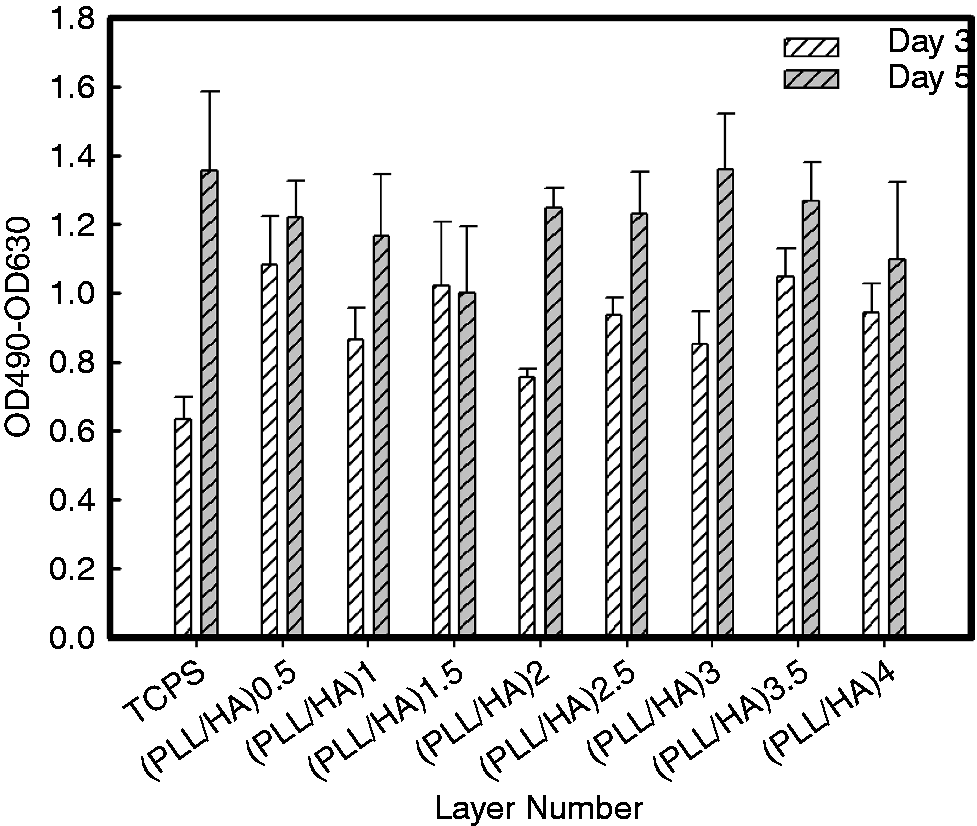
Morphologies of NSPCs on PLL/HA multilayer films
Figure 4 shows phase-contrast microscopy images of NSPCs cultured on PLL/HA multilayer films after 3 and 5 days of incubation. The NSPCs did not adhere to the glass cover slide or TCPS substrate but attached to the PLL/HA multilayer films, without requiring the addition of inductive chemical regents. After NSPC attachment, cell migration from the spheroids and neurite outgrowth were investigated. Certain regions displayed a complicated network with differentiated phenotypes. Our previous study showed that short and crooked processes were observed on PEM films with PLGA as the terminal layer, whereas the cells formed many long and straight processes on PEM films with PLL as the terminal layer (not shown here). In contrast, the neurite morphologies did not vary with the surface charge in the PLL/HA system described here.
Phase-contrast images of the differentiated cell phenotypes of NSPC spheres cultured on PLL/HA (n = 0.5), PLL/HA (n = 1), PLL/HA (n = 3.5), PLL/HA (n = 4), and glass after 3 and 5 days of culture. HA: hyaluronic acid; PLL: poly-L-lysine.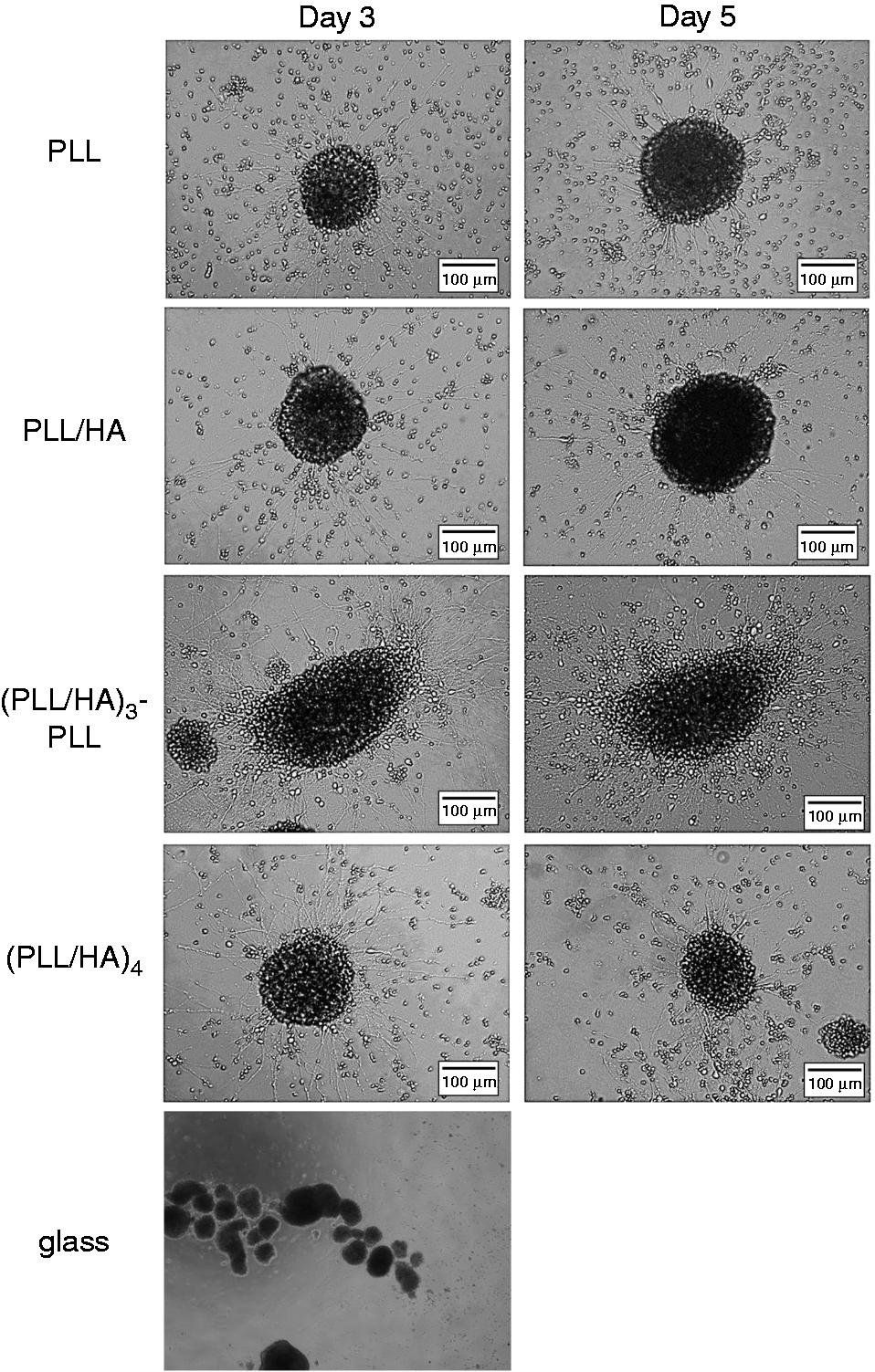
Measurement of the neurite outgrowth length
Quantitative measurements of the neurite outgrowth length were compared between PLL/HA multilayer films with different layer numbers and terminating outer layers. The results were adjusted according to the Bonferroni correction in which the p values were multiplied by the number of comparisons made as a more stringent test of significance. As shown in Figure 5(a), NSPCs displayed longer neurites on the multilayer films with PLL as the terminal layer than on PLL/HA multilayer films with HA as the terminal layer. In particular, as shown in Figure 5(a)–(c), the NSPCs displayed very long neurites and formed a well-connected neuron network. In addition, the neurite outgrowth lengths of 10 spheroids on sequential PLL/HA multilayer films were characterized, and the results are shown in Figure 5(b). It was found that most of the neurite outgrowth lengths were between 200 and 400 µm after only 3 days of culture. In addition, except the layer numbers of 0.5 and 1, the average neurite outgrowth lengths on the PLL/HA multilayer films were more than 300 µm. Furthermore, the neurite outgrowth length of NSPCs cultured on PLL/HA multilayer films with a positively charged terminal layer were significantly different from that of NSPCs cultured on PLL/HA multilayer films with HA as the terminal layer (*p < 0.0125 and **p < 0.0025). It was found that PLL/HA is a suitable material for NSPC induction that enhances neurite outgrowth. However, in contrast to the other PEM films, the effect of the layer number on neurite outgrowth was not obvious (data not shown). This result suggests that the charge of the terminal layer is a more important factor for neurite outgrowth in this system.
(A) Images showing the neurite morphologies on (a) PLL/HA (n = 0.5), (b) PLL/HA (n = 1), (c) PLL/HA (n = 3.5), and (d) PLL/HA (n = 4). The red arrows indicate the locations of neurites. The circles indicate the formation of a neural network. (B) Quantification of the process lengths of NSPC spheres on sequential PLL/HA multilayer films cultured under serum-free conditions at 250 neurospheres per cm2 for 3 days. The lengths of all of the processes per neurosphere were determined from the edge of the neurosphere to the tips of the processes when in a linear form. The values represent the means ± SEM for at least six independent neurospheres. The asterisks denote significant differences in the average process lengths on the different PLL/HA multilayer films (*p < 0.0125 and **p < 0.0025 , two-tailed t test with Bonferroni correction for multiple comparisons) as determined by Student’s t-test. HA: hyaluronic acid; PLL: poly-L-lysine.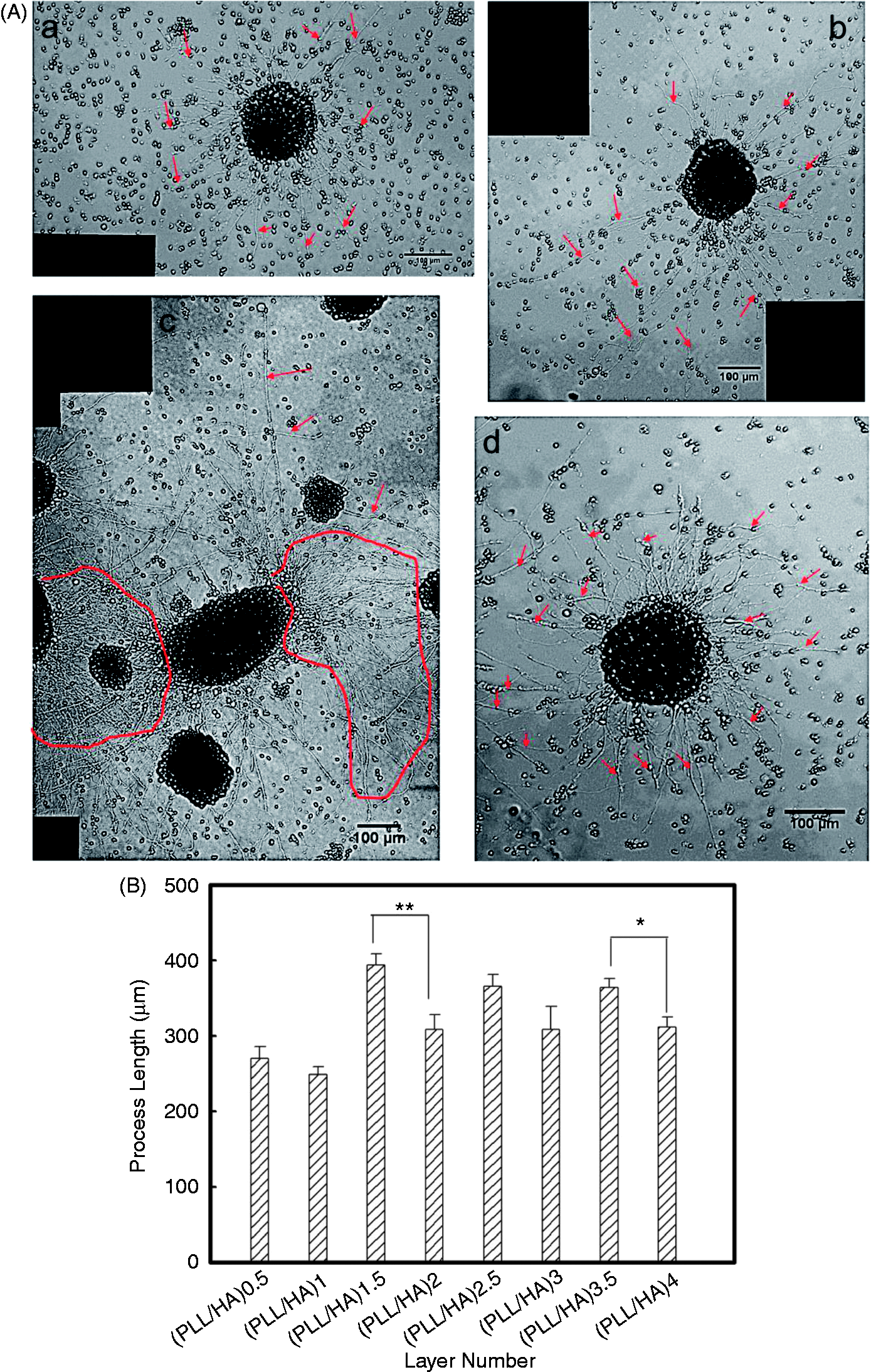
Characterization of NSPC differentiation on PLL/HA multilayer films
NSPCs have the capacity to differentiate into three lineages: neurons, astrocytes, and oligodendrocytes. However, NSPC culture under different culture conditions produces heterogeneous populations, and differentiated cells cannot be discriminated only by their morphologies. Neurons and astrocytes are the most common differentiated types of NSPCs, and immunocytochemical staining can be used to analyze these differentiated cell types. In the present study, to determine the differentiated phenotypes of NSPCs on the sequential substrates, MAP-2 was used as the neuron marker and GFAP was used as the astrocyte marker. Representative images are shown in Figure 6, and it is obvious that the NSPCs successfully differentiated into both neurons and astrocytes on the PLL/HA multilayer films, without requiring the addition of chemical factors. In contrast to previous reports in the literature, the NSPCs attached to the surfaces with HA as the outermost layer and differentiated into specific cells. The results suggest that the charge of the terminal layer is not a key factor affecting differentiation induction in this system.
Fluorescence photomicrographs showing the differentiated cell phenotypes of NSPC spheres after 3 days of culture. Anti-MAP-2 (red) and anti-GFAP (green) are immunoreactive with differentiated neurons and astrocytes, respectively. The fluorescence photomicrographs show NSPCs cultured on PLL/HA multilayer films. (a) PLL/HA (n = 0.5), (b) PLL/HA (n = 1), (c) PLL/HA (n = 1.5), (d) PLL/HA (n = 2), (e) PLL/HA (n = 2.5), (f) PLL/HA (n = 3), (g) PLL/HA (n = 3.5), and (h) PLL/HA (n = 4).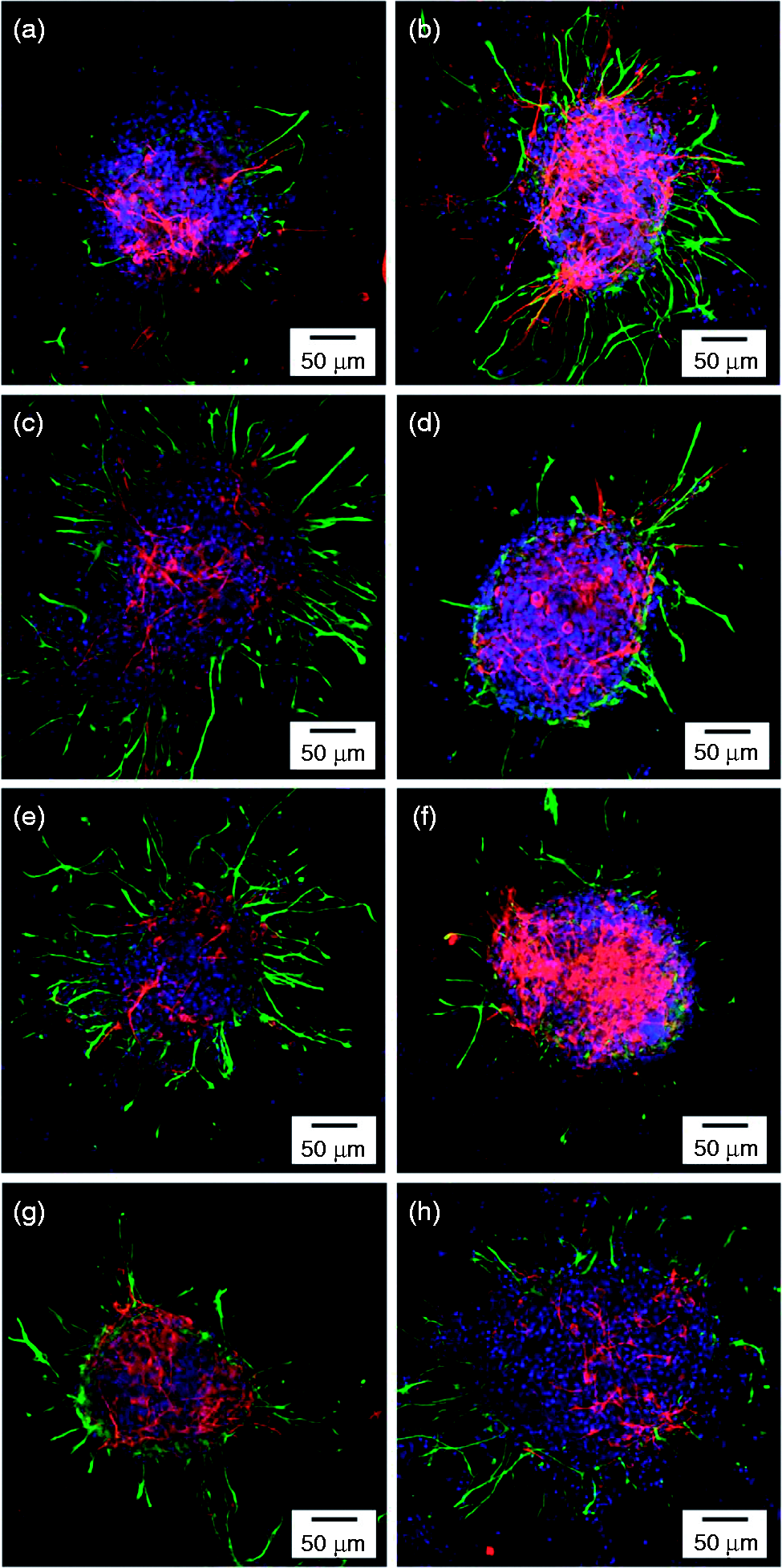
Furthermore, the results of the quantification of the differentiated cell phenotypes on the sequential PLL/HA multilayer films, which were determined by counting the numbers of MAP-2- and GFAP-positive cells, are shown in Figure 7. In addition, the percentage of differentiated NSPCs on the PLL/PLGA PEM films was examined. Besides, the significance values were further adjusted for comparisons between multiple groups using the Bonferroni correction and the number of independent comparisons made was n = 4. As shown in Figures 7(a) and (b), it was found that the percentage of differentiated neurons increased slightly as the layer number increased (*p < 0.0125 for PEM (n = 3.5)). However, the terminal charge did not show a significant effect on neuron induction. In contrast, Figure 7(c) shows that the percentage of differentiated neurons on PLL/PLGA (n = 3.5) was higher than that on PLL/PLGA (n = 4), indicating that the terminal charge is more important in this system. It is possible that the charge effect may be reduced because HA is the main ECM component of brain tissue and after composed with PLL which may reduce the surface charge. In addition, the hydration effect of HA may reduce the stiffness of PLL/HA multilayer films as the layer number is increased, which may enhance the percentage of neurons among the differentiated cells.
Quantification of (a) the percentage of cells that differentiated into neurons and astrocytes, (b) the neuron/astrocyte ratios for neurospheres on the sequential PLL/HA multilayer films, and (c) the neuron/astrocyte ratio for neurospheres on the PLL/HA and PLL/PLGA multilayer films. The cells were cultured on PLL/HA and PLL/PLGA multilayer films under serum-free conditions at 250 neurospheres per cm2 for 5 days and 3 days, respectively. The asterisks denote significant differences in the percentage of neurons on the different PLL/HA multilayer films (*p < 0.0125, two-tailed t test with Bonferroni correction for multiple comparisons) as determined by Student’s t-test. HA: hyaluronic acid; PLL: poly-L-lysine.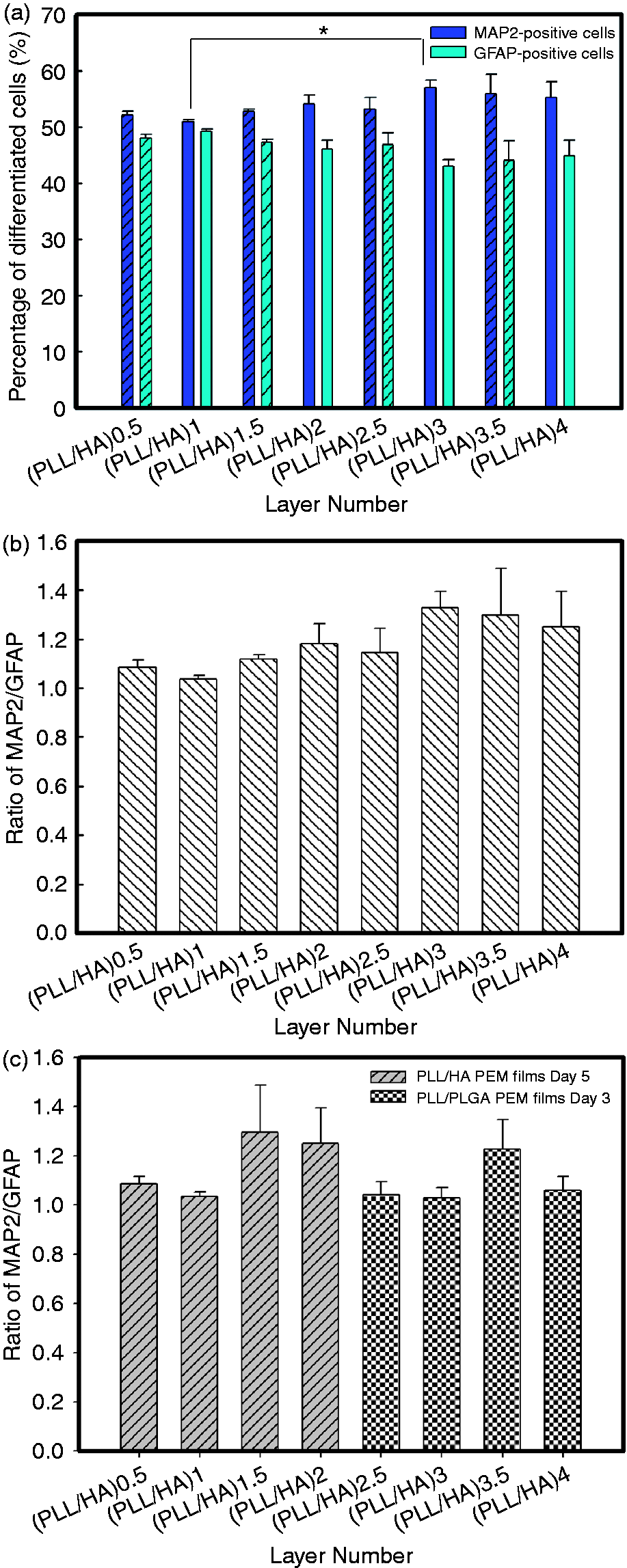
Characterization of functional neurons via immunostaining for synapsin I
Synapse formation is an important capability of functional neurons. Therefore, immunostaining for synapsin I was performed in this study to identify the functionality of the differentiated neurons on the PLL/HA multilayer films. Fluorescence photomicrographs showing synapsin I expression and indicating functional neurons on PLL/HA (n = 0.5) and PLL/HA (n = 1) after 3 days of culture are shown in Figure 8. Only the immunostaining for synapsin I shown in Figure 8(a) displayed high expression, and a dense network, with a large number of neurites and neural filaments, was investigated on both substrates. Furthermore, double staining for synapsin I and MAP2, as shown in Figure 8(b), was observed in the neural networks with interlocking axons and dendrites, confirming that the differentiated neurons were functional.
Fluorescence photomicrographs showing synapsin I expression, a marker of functional neurons, on PLL/HA multilayer films after 3 days of culture. (A) Immunostaining for synapsin I only. (B) Immunostaining for MAP2 (green) and synapsin I (red). (a) PLL/HA (n = 0.5) and (b) PLL/HA (n = 1).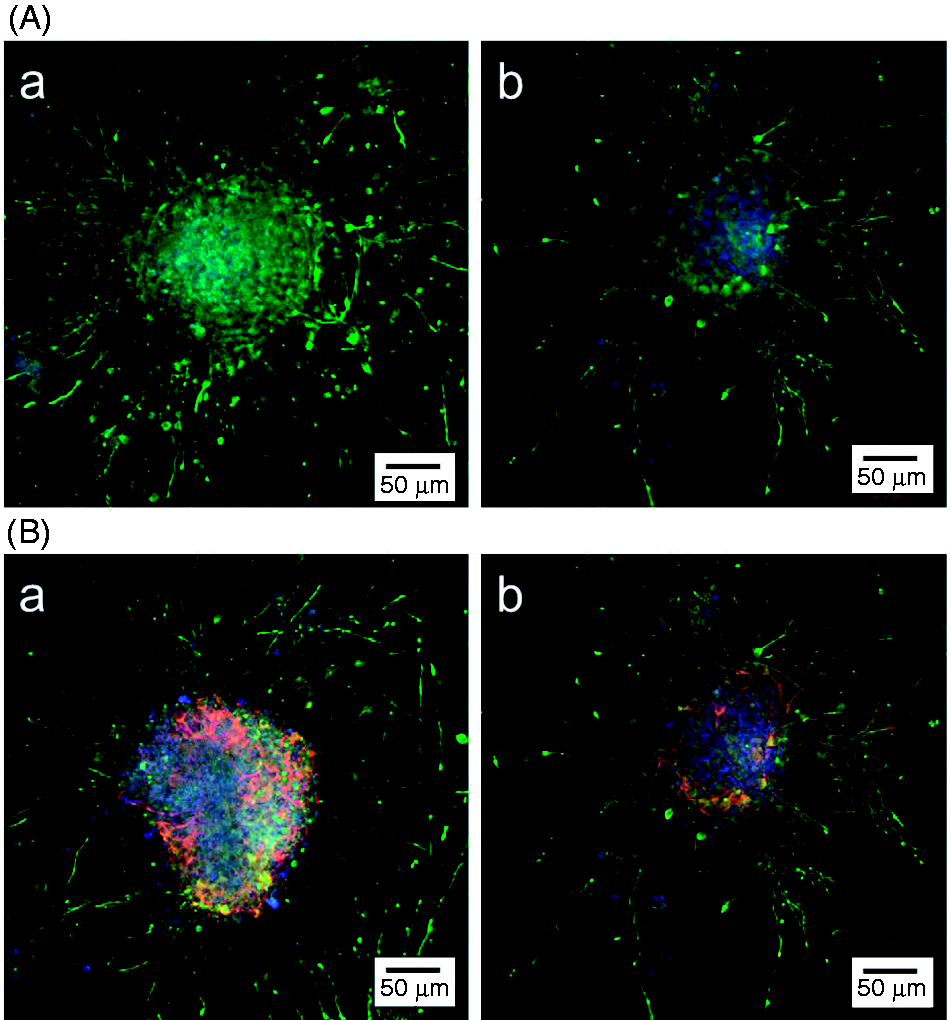
Conclusion
In conclusion, a native ECM-mimetic system was fabricated for NSPC induction and neuron culture. NSPCs could migrate from neurospheres and adhere to the sequential PLL/HA multilayer films, regardless of the type of terminal layer. In contrast, the neurite outgrowth length was longer when PLL served as the outermost layer than when HA did. However, the effect of terminal charge on the percentage of neurons in the PLL/HA system was not as strong as in the PLL/PLGA system. It has been suggested that HA is an important ECM component in brain tissue and that neuron-HA interactions are stronger than neurons’ interactions with other negatively charged polyelectrolytes. This material system provides a biomimetic platform for NSPC induction and the culture of neuron-related cells by improving NSPC adhesion and differentiation and generating large amounts of functional neurons. This material pair may be useful in neural engineering and for surface modification of neural culture systems.
Footnotes
Acknowledgment
The authors thank professor Chii-Wann Lin for QCM instrument support.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Financial support for this study was obtained from Chang Gung University and from grants from the National Science Council Taiwan. The grant numbers are NSC 101-2221-E-182-012 and NSC 102-2221-E-182-007.