
Abstract
The mechanism of action underlying the intriguing prominent bioactivity of urinary bladder matrix (UBM) for in situ tissue regeneration of soft tissue defects remains to be elucidated. It is speculated that the activity of UBM for cell adhesion, migration, and activation is inherent. The bioactivity of UBM for in situ tissue regeneration and its relation with the structure and intact soluble components of UBM were investigated in comparison to a collagen-based scaffold, PELNAC (PEL). We isolated the soluble component of the two materials with urea buffer, and evaluated the respective effect of these soluble components on the in vitro adhesion and migration of L929 fibroblasts. The spatiotemporal pattern of endogenous-cell ingrowth into the scaffolds and cell activation were investigated using a model of murine subcutaneous implantation. UBM is more capable of promoting the adhesion, migration, and proliferation of fibroblasts than PEL in a serum-independent manner. In vivo, as compared with PEL, UBM exhibits significantly enhanced activity for fast endogenous cell ingrowth and produces a more prominent pro-regenerative and pro-remodeling microenvironment by inducing the expression of TGF-β1, VEGF, MMP-9, and murine type I collagen. Overall, our results suggest the prominent bioactivity of UBM for in situ tissue regeneration is inherent.
Keywords
Introduction
Soft tissue defects, such as cutaneous wounds, peritoneal defects, volumetric muscle loss and so on, are ubiquitous problems in clinical medicine. Patients suffer these defects secondary to surgical or traumatic injuries, chronic diseases, and natural aging. Loss of soft tissues leads to functional impairment and aesthetic flaw that are challenging to treat. 1 Moreover, if the treatment is retarded, risks of infection and progressive injuries increases, further complicating the situation. 2 Therefore, the primary goal of dealing with soft tissue defects is reconstructing and restoring the lost tissue rapidly in order to prevent infection and alleviate the patients’ suffering.
Although autologous tissue transfer is still regarded as the gold standard in the repair and restoration of soft tissue defects, it requires lengthy surgical procedures and results in donor site morbidity. In recent decades, strategies of using bioactive materials to promote in situ tissue regeneration (ISTR), or in situ tissue engineering, have been established and developed. The core principle of ISTR is to utilize the body’s own biological resources and regenerative potential by means of appropriate cell-free bioactive materials capable of recruit endogenous progenitor cells to the sites of injury and unlock the regenerative capability of these cells. 3 Correspondingly, ISTR avoids big challenges in in vitro tissue engineering such as time-consuming, costly procedures of cell isolation and culture, and difficulty to maintain cell/tissue viability. 4
Currently, bioactive materials implemented for ISTR can be classified into two main groups, the synthetic grafts and the naturally-derived ECM grafts. The advantages of synthetic grafts, such as collagen I-based porous scaffolds, are batch-to-batch quality homogeneity and the absence of the risk of disease transmission. 5 Notwithstanding, some naturally-derived ECM scaffolds show intriguing enhanced bioactivity for ISTR. Although clinical randomized controlled studies are lacking, there are still many case reports6,7 and animal experiments8,9 that suggest the inherent pro-regenerative activity of natural ECM scaffolds.
Urinary bladder matrix (UBM) is one type of naturally-derived ECM grafts that draws much attention regarding its pro-regenerative potential. It is composed of urinary basement membrane and the upper layer of lamina propria that are decellularized by peracetic acid. 10 Its main components are structural proteins such as type I collagen and laminin, glycoproteins, and growth factors. 11 UBM has been widely used in clinical practice to repair soft tissue defects. It is worth noting that UBM can be used alone to promote the healing and dermal reconstruction of complex wounds with exposed tendon and bone, 12 while collagen I-based dermal substitutes need to be combined with vacuum sealing drainage (VSD) systems to initiate and prompt the dermal reconstruction of these complex wounds. 13 However, due to ethical issues surrounding the harvest of patient tissue, the mechanism underlying the difference between UBM and synthetic dermal substitutes in ISTR remains to be elucidated.
Some previous studies focused on the immune-modulating effect of UBM that usher tissue reconstruction including the induction of macrophage polarization and the recruitment of T cells.14,15 For example, one study noted the role of UBM in activating adaptive immune responses, 16 and another demonstrated that UBM induces adaptive immune response to corneal epithelial defect trauma in mice, promoting corneal regeneration and visual recovery. 17 A study investigated the pro-regenerative activity of soluble and structural components of UBM digest, and it demonstrates that both the soluble and structural components can stimulate the chemotaxis of human peripheral stem cells, reduce stem cell proliferation, and form an anti-inflammatory and pre-remodeling response in THP1 human macrophages, thereby promoting tissue remodeling. 18 However, many soluble components of ECM, including stromal cell proteins and growth factors, can be combined with structural components. The two main structurally-bound growth factors, VEGF and bFGF contained in UBM, cannot be detected after they are digested by pepsin. This is probably due to the digestion by acid and pepsin that denatures the soluble components and hydrolyzes the protein, leaving only the degraded matricryptic peptide. Therefore, the structurally-bound growth factors are undetectable both before and after UBM degradation. It cannot be ruled out that the activity of UBM structural components is derived from the soluble components combined with them. Using urea supplemented buffer, another study isolated intact soluble components of ECM generated by in vitro culture mesenchymal stem cells, 19 and suggested that ECM-bound molecules rather than the structural proteins play major roles in the induction of the proliferation, attachment, migration, and differentiation of mesenchymal stem cells. However, the experiments adopted by the research institute used matrix coatings for verification, and did not directly use soluble ingredients to culture cells to evaluate their effects on cell adhesion and chemotaxis behavior.
It is speculated that UBM also has the inherent biological activity of promoting cell adhesion and movement, partly produced by non-collagen components.18,19 Therefore, this study aimed to confirm the inherent and enhanced bioactivity of UBM for ISTR as compared with a type of collagen-based scaffold, PELNAC artificial dermis (PEL). The soluble components of UBM are to be characterized for the adhesion and migration of fibroblasts, and the inherent activity that promotes cell growth in a three-dimensional environment and induces their activity. We used urea-supplemented buffer to thoroughly isolate the soluble components of UBM, and characterized the effects of these components on the adhesion and migration of L929 fibroblasts. Furthermore, cells were directly seeded on the scaffold, and fibroblast adhesion and growth are evaluated in a three-dimensional environment. Finally, the infiltration and activation of fibroblasts were evaluated in a mouse implant model. We hope that this study may provide clues for the design and screening of novel materials to promote ISTR.
Materials and methods
Materials
CytalTM wound matrix 1-layer sheets and Micromatrix® powder of UBM were supplied by ACell, Inc., and PELNAC® dermal substitutes were supplied by Gunze Limited, Inc. L-929 fibroblast cell line was purchased from Shanghai Zhongqiao Biotech. DMEM, Penicillin/Streptomycin, and fetal bovine serum were purchased from Gibco. Transwell plates (6.5 mm diameter polycarbonate membrane filters with 8.0 μm pore) was purchased from Corning. Dialysis tube was purchased from Shanghai Yuanye Bio-Technology Co., Ltd. Rabbit primary antibodies against Mouse ER-TR7,α-SMA, TGF β1, VEGF, MMP 9, and Collagen I antibodies were supplied by Abcam. Donkey anti-rabbit IgG secondary antibody was purchased from Boster Biological Technology Co., Ltd.
Extraction of soluble components
Firstly, PEL was cut into small pieces. Then, 250 mg of Micromatrix® UBM powder and the PEL pieces was respectively added into, and immersed in, 5 mL buffer solution of 150 mM NaCl and 2 M urea. The resulting solutions were incubated with mild agitation at 4°C for 2 days. Afterwards, the solutions were centrifuged at 25,000 g for 10 min, and the supernatants were dialyzed against ultrapure water for 3 days. The dialyzed solutions were diluted 10 times with DMEM, filtered through 0.22 μm membranes, and then stored at -80°C until use.
The adhesion of fibroblasts on the scaffolds
Under sterile conditions, CytalTM wound matrix UBM and PEL scaffolds were punched into discs of 6-mm diameter and placed in 96-well plate. Then, 20 μL of DMEM was added to swell the scaffold for 1 h. Prior to being seeded onto the scaffolds, L929 fibroblasts were cultured to 90% confluence, digested off the culture dish with trypsin, and resuspended in DMEM medium containing 10% FBS and 1%P/S (or DMEM medium alone). 200 μL of cell suspension (2.0×105 cells/ml) was then transferred gently to the surfaces of the incubated scaffolds by pipettes. After 6-h incubation in a 37°C, 5% CO2 incubator, the culture media were removed, and the scaffolds were rinsed with 200 μL PBS three times, 5 min for each time, and then fixed in PBS containing 2.5% glutaraldehyde overnight. The scaffolds were subjected to sequential dehydration in graded ethanol (50, 70, 80, 90, 100%). After immersion in hexamethyldisiloxane, the specimens were air dried for 3 min. All samples were stored in a dry environment and then gold sprayed and observed using scanning electron microscope.
The ingrowth of fibroblasts into the scaffolds
The previous steps were the same as the cell adhesion assay, in addition to being cultured in DMEM medium (10% FBS, 1% P/S) for 3 days and then the scaffolds were fixed overnight in 4% paraformaldehyde and paraffin-embedded, sectioned at 6 μm thick. The sections were baked at 60°C for 20 min, then dewaxed three times in xylene for 10 min each time, and hydrated in gradient ethanol (100%, 90%, 60%, 30%, 0%) for 5 min each time. Finally, the sections were sealed with anti-fluorescence attenuation sealer containing DAPI and then imaged under a Zeiss ortho-fluorescence microscope.
The adhesion and migration of fibroblasts induced by soluble components of the scaffolds
L929 fibroblasts were digested with trypsin and resuspended with UBM and PEL soluble components, then evenly distributed in a 6-well plate with 2 mL per well. The cells were allowed to adhere for 2.5 h in a 37°C, 5% CO2 incubator and then the medium was aspirated and the plate was washed with PBS one time. Cell adhesion on the 6-well plate observed by an inverted microscope.
The same fibroblasts digested as above were resuspended in serum-free medium after 12 h of serum withdrawal starvation. 100 μL cell suspension (2×104 cells/well) was added in the upper chamber of transwell plate, and in the lower chamber, 500 μL DMEM medium (10% FBS, 1% P/S) was added to promote cell adhesion in a 37°C, 5% CO2 incubator. After 1 h, the culture medium in the lower chamber was replaced with 500 μL UBM and PEL soluble components, and then continued to be placed in the cell culture chamber. Each group was examined for two time points, and three samples were repeated for each time. After 12 and 24 h, the chamber was taken out and the excess cells on the inner surface were gently wiped off with a cotton swab. Then the chamber was stained with 0.1% crystal violet for 20 min, and washed three times with PBS. Finally, the chamber was placed on an inverted microscope to take images, and the number of migrated cells in the UBM and PEL soluble components-treated groups was counted respectively. For both the adhesion and migration test, 3 experiments were conducted with 2 replicate samples in every experiment at each time point.
Surgical procedure
All surgical procedures were performed in accordance with the NIH Guidelines for Care and Use of Laboratory Animals and with approval from the Animal Ethics Committee of Jiangnan University and Use Committee (JN.No20171115c0600130). Twelve male C57BL/6 mice, aged 6–8 weeks, were randomly assigned to two separate groups. The mice were anesthetized by intraperitoneal injection of a mixture of ketamine (90 mg/kg) and xylazine (10 mg/kg). Under aseptic conditions, a small incision was made in the middle of the mouse's back and two pockets were formed by blunt separation to the left and right of the incision. UBM and PEL were randomly assigned to be implanted under the skin on the right and left sides of each mouse, and the incisions were closed with 6–0 absorbable sutures. UBM was implanted with the lamina propria side facing the skin.
Histological and immunohistochemical staining
Mice were euthanized at 1 and 2 weeks after implantation, and the scaffolds were excised along with the surrounding native tissue. It was then immediately fixed in 4% paraformaldehyde solution, paraffin-embedded, sectioned at 5 μm thick, and then stained by standardized techniques for H&E staining and immunohistochemical staining. H&E staining was performed in accordance to manufacturer protocol. Immunohistochemical staining was conducted with Rabbit primary antibodies against mouse ER-TR7, α-SMA, TGF-β1, VEGF, MMP-9, and Col-I overnight at 4°C, followed by donkey anti-rabbit IgG secondary antibody for 30 min. Staining was visualized with diaminobenzidine (DAB). Cell nuclei were counter-stained by hematoxylin.
Statistical analysis
The total number of positive cells was counted by randomly taking five pictures for each sample. All numerical results were expressed as mean±SD (standard deviation). Statistical analysis was performed using two-way analysis of variance followed by Graphpad Prism 8.0 analysis, where p < 0.05 indicates statistical significance.
Results
UBM promotes the adhesion and ingrowth of fibroblasts in vitro
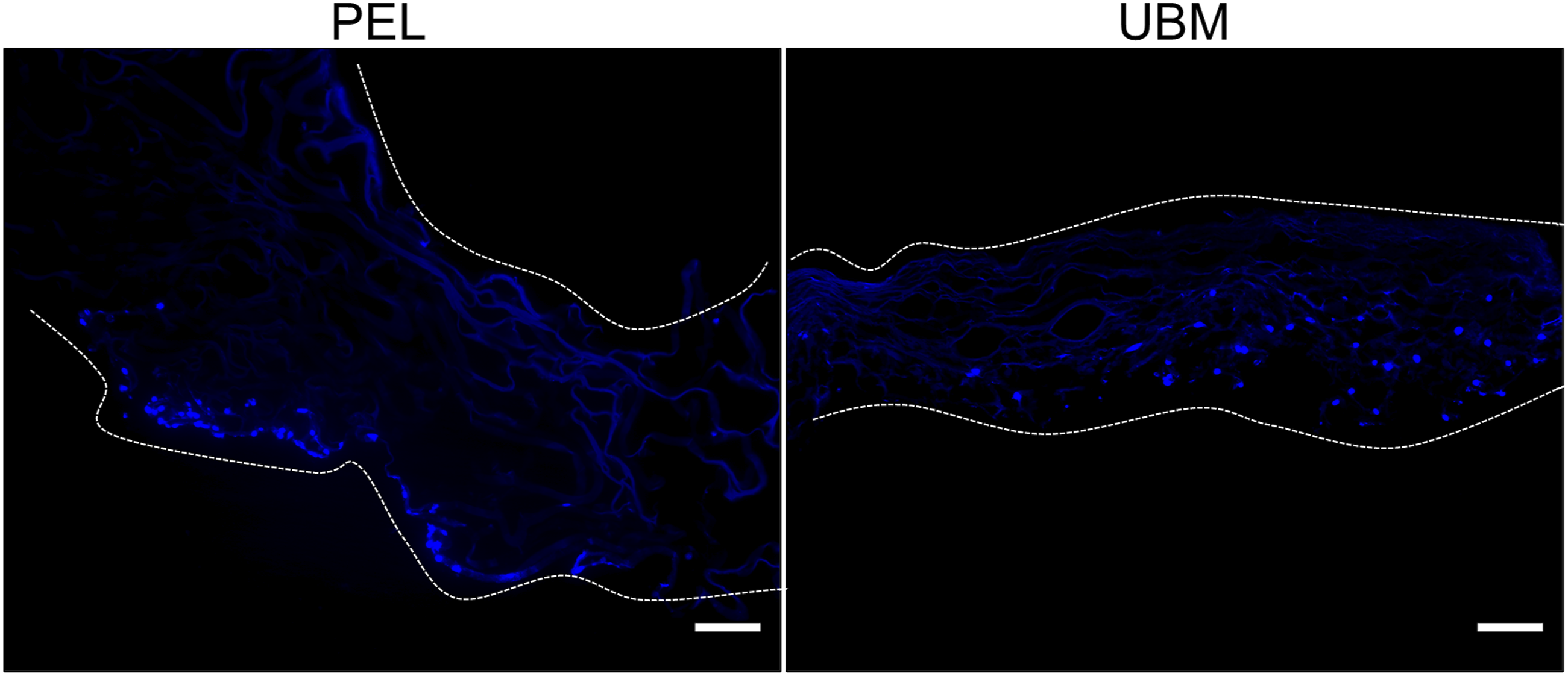
The adhesion of L929 fibroblasts on the lamina-propria side of UBM as compared with PEL was investigated by SEM. The SEM images show that the lamina propria side of UBM scaffolds was composed by inter-connected fibers, and the PEL scaffolds displayed a three-dimensional pore structure. A large number of fibroblasts adhered to the surface of UBM when cultured with complete medium, but fewer cells were seen on the surface of PEL (Figure 1(a)). When the medium was used alone, there were almost no cells adhering to the surface of PEL, while some cells were still visible on the surface of UBM (Figure 1(b)). No cells were observed on the surface of unseeded PEL and UBM controls (Supplemental Figure S1). We grew fibroblasts on the surface of both materials to observe the cells growing in from the surface to the center of the scaffold after 3 days. The cells were stained dark blue by DAPI. Figure 2(a) shows that the cells remained on the surface of the PEL, while the cells on the UBM have mostly grown into the center of the scaffold. Representative images of cells grown on the surface of PEL and on the lamina propria of UBM after adherence for 6 h in DMEM medium containing 10% FBS (a) and in DMEM medium alone (b) are shown. The figure shows the low magnification and high magnification images taken by scanning electron microscopy. Representative images of cells on the PEL surface as well as on the UBM lamina propria after growing in 3D are shown. Cells were stained dark blue by DAPI dye. (scale bar is 100 μm).
UBM soluble components enhance adhesion and migration of fibroblasts in vitro
The cell adhesion assay (Figure 3(a)) showed that after 2.5 h, more fibroblasts cultured with UBM soluble components adhered to the bottom of the well plate. The results of the quantitative statistics of the relative number of adherent cells (Figure 3(c)) supported this, and the difference was statistically significant (p < 0.001). We used transwell assay to assess the effect of soluble components on fibroblast motility. Representative images of fibroblast migration at 12 and 24 h are shown in Figure 3(b). It can be seen that UBM is more effective in promoting fibroblast migration. The results of the quantitative statistics of the relative number of migrating fibroblasts (Figure 3(d)) were consistent with the previous results, and the differences were statistically significant (p < 0.001). UBM promotes cell migration nearly twice as much as PEL. Representative images of PEL and UBM soluble component cultured cells after 2.5h of adhesion are shown in Figure a (scale bar 100 μm). Representative images of soluble component chemotactic cells at 12 and 24 h low and high magnification are shown in Figure b (low magnification scale 100 μm, high magnification scale 25 μm). Cells were stained purple by crystalline violet. Quantitative statistical plots of fibroblast adhesion and migrating cell counts (c, and d). The data points represent mean ± SD (n = 6, **: p < 0.01, ***: p < 0.001).
UBM promotes ingrowth of endogenous cells
The H&E staining results (Figure 4) showed that at 1 week after implantation, there were few cells in the PEL and a large number of cells in the UBM. At 2 weeks after implantation, the cells migrating into the PEL were concentrated in the non-central area of the scaffold, while more cells were dispersed in the UBM. Accumulation of neutrophils was not observed in the interface between subcutaneous tissue and UBM or PEL (Figure 4), indicating no acute inflammation happened. No fibrous capsules appeared in the tissue/scaffold interface until 2 weeks after implantation (Figure 4). In vivo experiments again demonstrated that UBM promoted cell adhesion and migration to the center of the scaffold as compared with PEL. The H&E results of PEL and UBM embedding materials at 1 and 2 weeks postoperatively are shown in Figure a–d are low magnification images (Scale bar is 100 μm), and e–h are the corresponding high magnification images in their red boxes (Scale bar is 25 μm).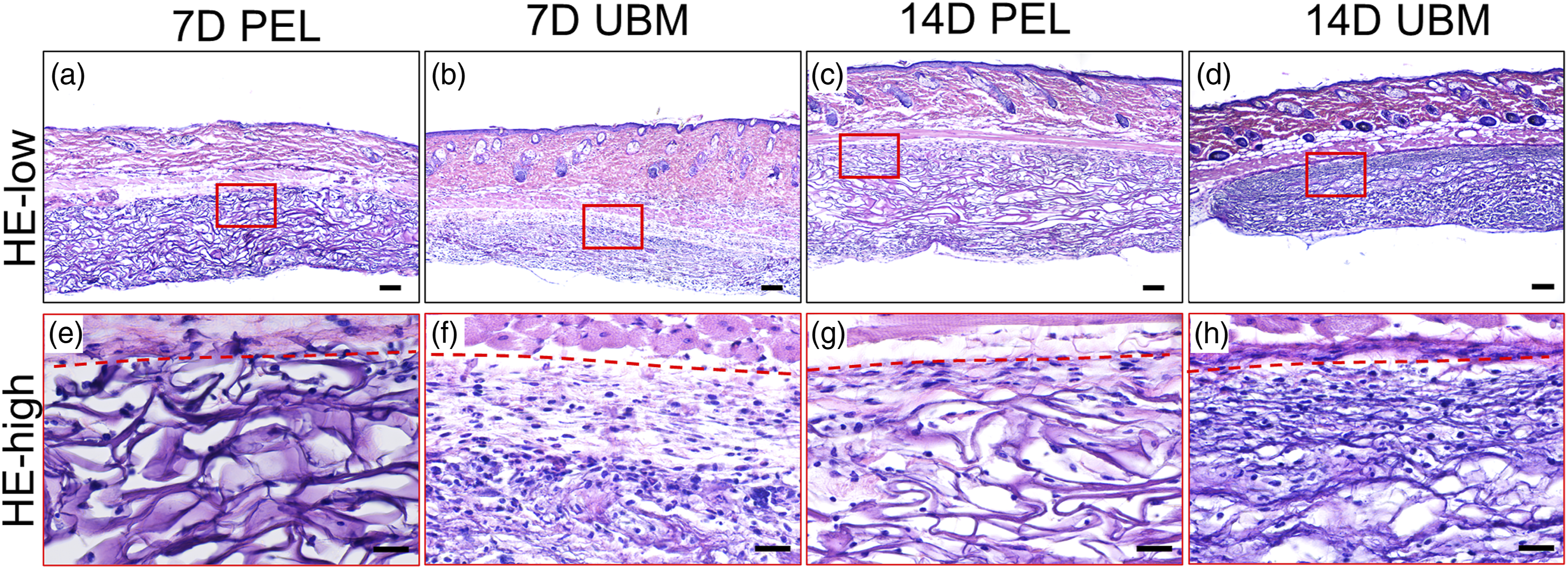
We performed anti-ER-TR7 staining to verify the cell type, and the results (Figure 5(a)) showed that most of the cells stained positive, meaning that most of the cells dispersed in the UBM were indeed fibroblasts. The results of anti-α SMA staining (Figure 5(a)) showed that at day 7 after implantation, most of the fibroblasts in the UBM were activated as myofibroblasts, while the number of activated cells in the center of the PEL scaffold was lower as compared with the UBM scaffold (Figure 5(b)). The number of α-SMA+ cells in UBM scaffold appeared to reduce at day 14 compared with day 7 (Figure 5(c)). The results of counting the number of positive cells in the scaffold center area were similar and statistically significant (Figure 5(b) and (c)). The anti-ER-TR7 and α-SMA results of PEL and UBM at 1 and 2 weeks postoperatively are shown in Figure a (scale bar is 25 μm). The cells were stained brownish-yellow. The differences in the quantitative statistics of the number of anti-ER-TR7 (b) and α-SMA (c) staining positive cells were statistically significant. The data points represent mean ± SD (n = 6, ***: p < 0.001).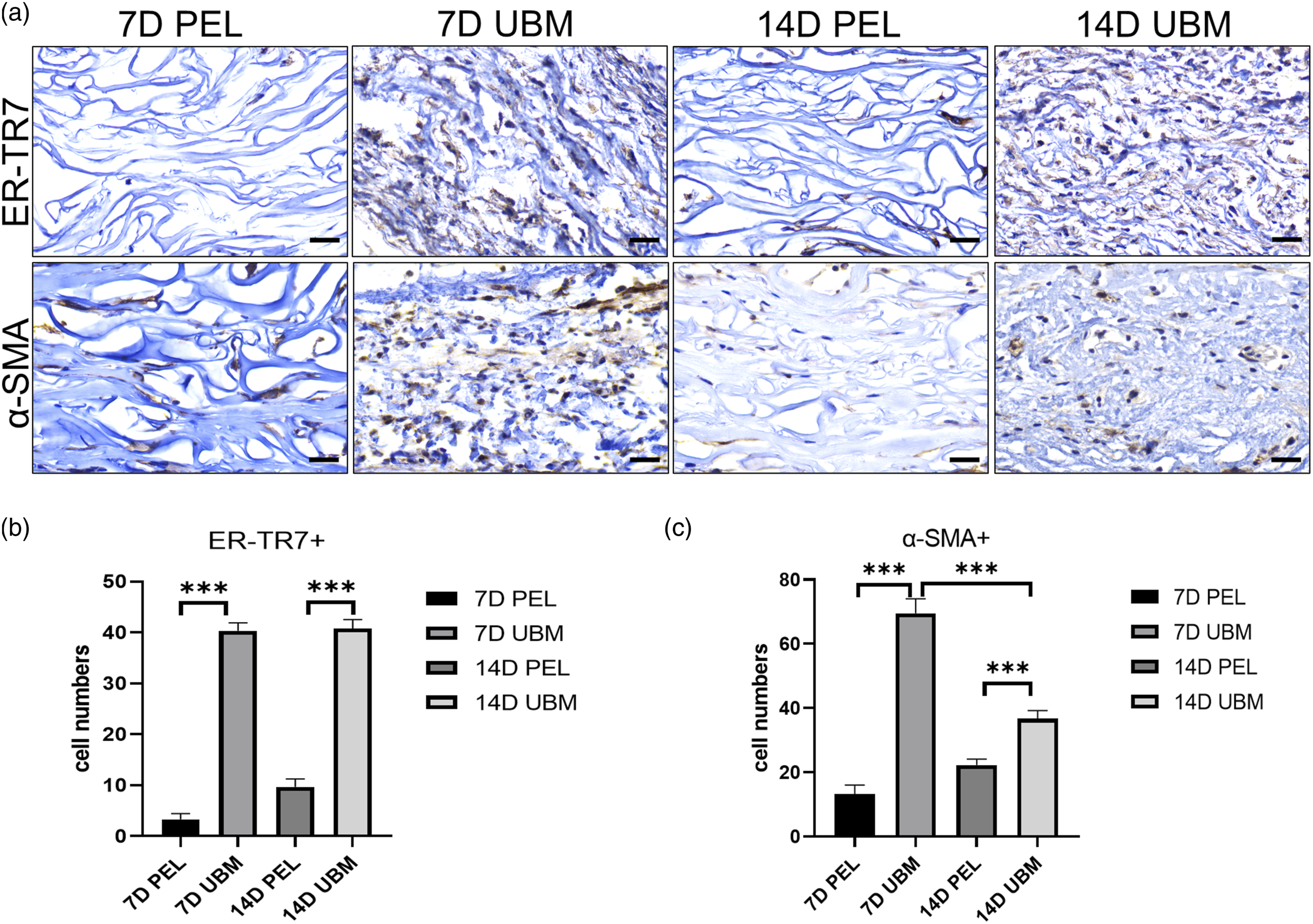
UBM produces pro-regenerative and pro-remodeling microenvironment
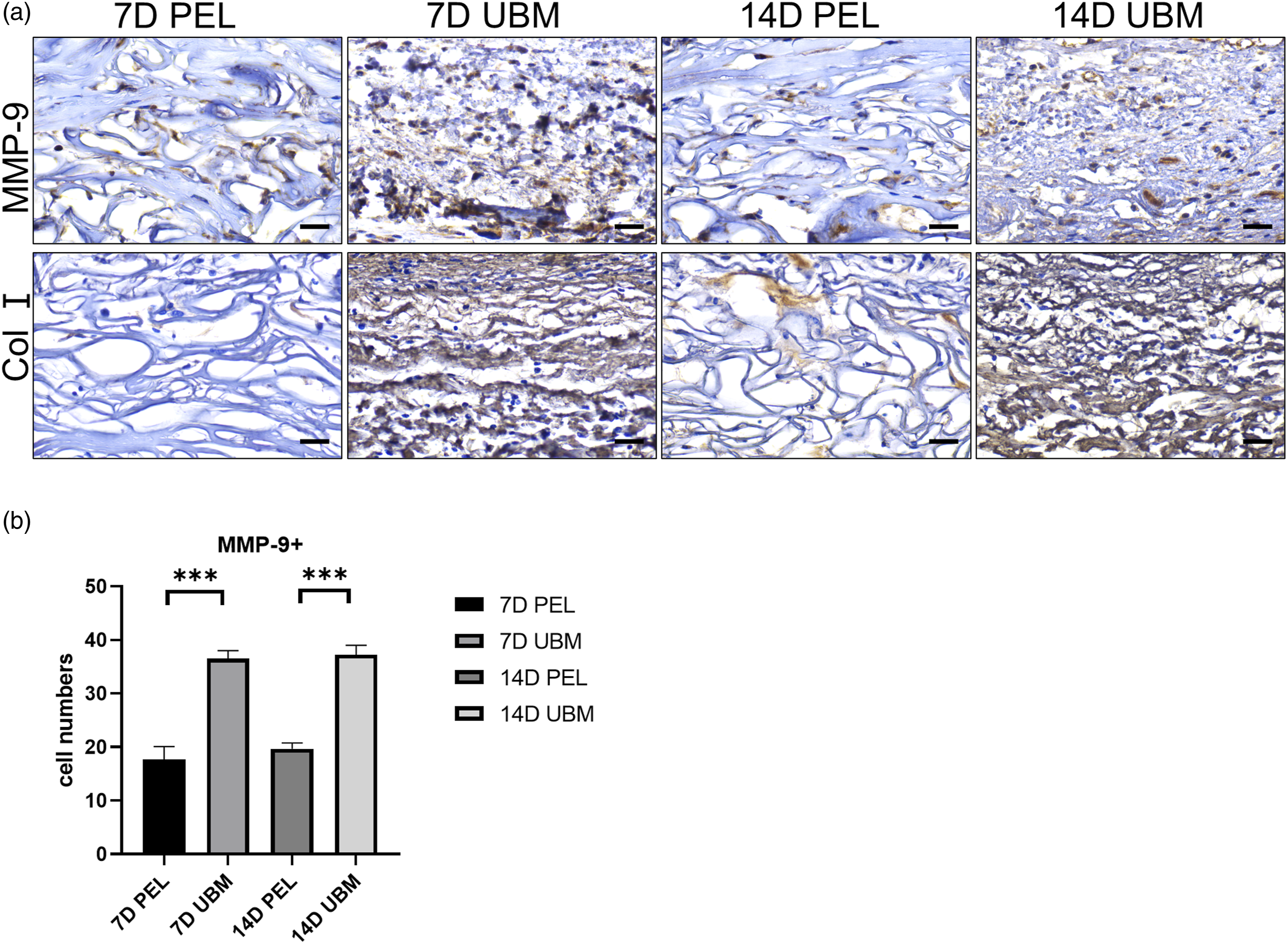
The results of anti-TGF β1 and anti-VEGF staining confirmed that the distribution of cells was in a similar spatiotemporal pattern of myofibroblast distribution (Figure 6(a)). The number of TGF-β1+ (Figure 6(b)) and VEGF+ (Figure 6(c)) positive cells in the central region of the scaffold was counted as seen at 1 and 2 weeks after implantation, and the number of positive cells in the UBM scaffold was significantly higher than that in the PEL, and the results were statistically significant. It is evident that UBM can create a regenerative microenvironment. The distribution of MMP-9+ cells (Figure 7(a)) showed that the number of positive cells in the UBM was significantly higher than that in the PEL at 1 and 2 weeks after surgery, and the results were statistically significant when counting the number of MMP-9+ positive cells in the central area of the scaffold (Figure 7(b)). The results of type I collagen deposition showed that a large amount of type I collagen was present in the UBM at 1 week. It is evident that UBM can promote scaffold remodeling. The anti-TGF-β1 and VEGF of PEL and UBM at 1 and 2 weeks postoperatively are shown in Figure a (scale bar is 25 μm) The cells were stained brownish-yellow. Statistical differences in the number of positive cells stained for anti-TGF-β1 (b) and VEGF (c) were statistically significant by quantitative analysis. The data points represent mean ± SD (n = 6, ***: p < 0.001). PEL and UBM anti-MMP-9 and anti-Col I at 1 and 2 weeks postoperatively are shown in Figure a (scale bar is 25 μm). The cells were stained brownish-yellow.The differences in the quantitative analysis of the number of positive anti-MMP-9 (b) staining cells were statistically significant. The data points represent mean ± SD (n = 6, ***: p < 0.001).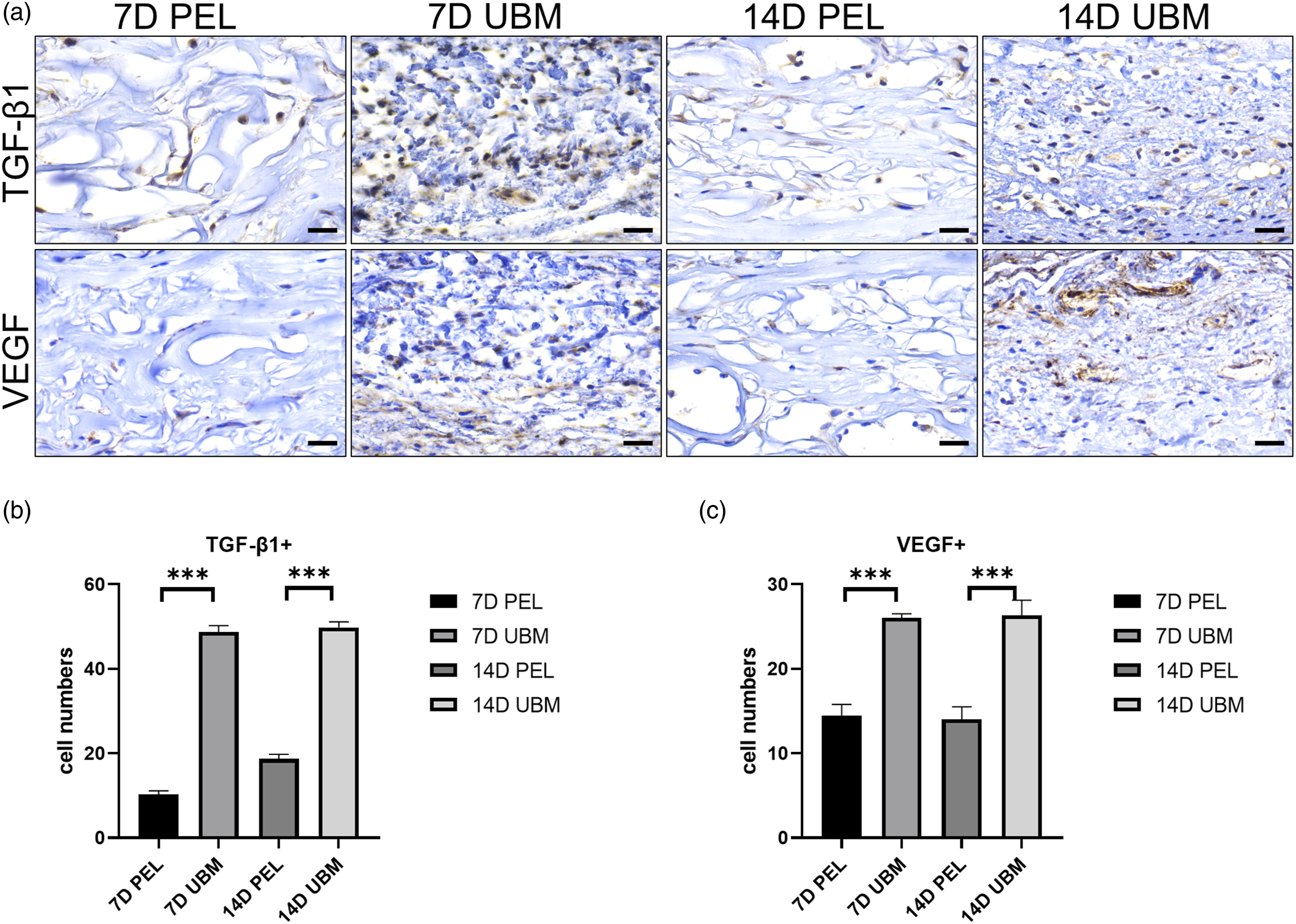
Discussion
UBM has been shown in various scenarios of tissue repair to display prominently enhanced activity for ISTR as compared with type I collagen-based synthetic scaffolds. 20 Whether the promotion of ISTR by UBM is dependent on its inherent activity for adhesion, migration, and activation of endogenous cells remains to be confirmed. For this reason, we evaluated the activity of UBM for ISTR in vitro and in vivo. It was verified that the fibrous meshwork structure and intrinsic activity of UBM aid in promoting fibroblast adhesion and migration, creating a remodeling microenvironment, and inducing cell activation, thus promoting scaffold remodeling and tissue regeneration.
Facilitated cell attachment to the scaffolds is a prerequisite for bioactive materials facilitated ISTR. 21 Our results suggest that the UBM itself promotes fibroblast adhesion and migration (Figures 1 and 2). In particular, the cells could still adhere to the UBM surface under FBS-free conditions, indicating that the inherent structure and activity of UBM render it the bioactivity of promoting cell adhesion (Figure 1(b)). Cell adhesion, cellular migration, and aggregation are all closely related to the ECM. 22 One study 23 showed that a three-dimensional porous scaffold prepared by decellularization from cartilage extracellular matrix displayed the activity for significant adhesion to bone marrow mesenchymal stem cells. Chitosan microspheres with an ECM-mimicking nanofibrous structure have been prepared and shown to have an excellent cell adhesion capacity. 24 It was also shown that amniotic membrane (AM) significantly induced the accumulation of Paxillin in HaCaT cells, and thus, it promoted their migration ability. 25 UBM, as a natural ECM material, has a similar fibrous meshwork structure that promotes cell adhesion and motility.
The components extracted from UBM also have a positive effect on cell adhesion and migration (Figure 3). UBM contains PDGF-BB 26 that is a powerful stimulator of the migration of fibroblasts.27,28 It has been shown that fibroblasts exhibit more pronounced chemotactic behavior towards scaffolds containing PDGF-BB. 29 We speculate that PDGF-BB contained in UBM soluble components is the main reason for promoting the adhesion and migration of fibroblasts.
After obtaining the above in vitro experimental results, we similarly demonstrated in an in vivo mouse implantation model that UBM can promote fibroblast infiltration (Figures 4 and 5), necessary for tissue repair. Prominent different patterns of recellularization between UBM and PEL were observed (Figure 4). Regarding the recellularization of UBM and PEL, similar findings were reported in other animal models such as a porcine model of hypospadias repair 30 and a murine model of cutaneous wound healing. 31 The lack of cell infiltration into PEL corresponds with a major clinical limitation of PEL, its slow integration into the tissue defects, especially bone- or tendon-exposed wounds. Consequently, VSD treatment is combined to aid PEL integration, whereas UBM displays significantly fast and sufficient integration in the scenarios of tendon-exposed wounds and infected wounds without the aid of VSD.32,33 Cross-linking is demonstrated to be a determining factor that hinder the integration of biomaterials by inducing moderate to strong foreign body reaction (FBR). Though, for PEL no significant FBR were found in our and others’ studies, 34 several studies verified the impact of collagen scaffold cross-linking agents and procedures 35 on the micro-environment which cells reside in and react to certain stimuli.
UBM also creates a microenvironment rich in TGF-β1, VEGF, and MMP-9 (Figures 6 and 7). Fibroblasts are activated and acquire a smooth muscle cell-like phenotype, called myofibroblasts, in response to TGF-β1 stimulation.36,37 Myofibroblasts are the major extracellular matrix (ECM) secreting cells. 38 TGF-β1 promotes their proliferation and migration, as well as the production of ECM components, 39 inducing the replacement of the temporary wound matrix, thus facilitating the normal tissue repair process.
Vascularization is the key to successful tissue engineering repair of damaged tissues, and VEGF is the main inducer of angiogenesis. One study designed a composite scaffold loaded with VEGF and demonstrated that the release of VEGF better promoted the growth of blood vessels on the scaffold. 40 As the polymer scaffold degrades, new tissue begins to form through an early stage of inflammation-mediated vascular remodeling.41–43 Matrix metalloproteinase-9, MMP-9, is thought to potentially promote ECM remodeling during neointima formation.44,45 MMP-9 plays an important role in the balance between the production and removal of structurally significant ECM constituents, which together with graft degradation will determine the evolution of vascular neointima. It has been shown that 6 months after graft implantation the scaffold degrades and is subsequently replaced by collagen-dominated neointima. 46 In this study, it is suggested that at 2 weeks after UBM implantation, dense type I collagen filled the middle of the scaffold and MMP-9 expression was significantly higher (Figure 7), indicating that the ECM is continuously forming. The microenvironment rich in TGF-β1, VEGF, and MMP-9 formed after UBM implantation in vivo can induce fibroblast activation and promote neo-vascularization, while continuously remodeling the ECM until graft degradation and promote in situ tissue regeneration.
Staged treatment, lengthy hospitalization, and the risks of infection are key drawbacks of collagen or other polymers-based synthetic grafts. 47 Some naturally-derived ECM grafts such as UBM show intriguingly prominent efficacy in the repair and restoration of complicated soft tissue defects including tendon-exposed, 12 lower-extremity ulcerous, and even other infected chronic wounds without assistance of negative-pressure wound therapy. 20 Though in-depth randomized controlled trials are required to strongly support the distinct potential of UBM in the induction of the repair of soft tissue defects, animal and laboratory studies consistently provide evidence to support the inherent activity of UBM for cell chemotaxis and activation, induction of pro-regenerative microenvironment, and even inhibition of microbes. 48 It is evident that UBM has an important role in clinical wound management. In addition, our study provided clues for the design and screening of more potential and customized bioactive materials that further optimize and accelerate the treatment of soft tissue defects under the ISTR strategy. However, it is acknowledged that this study is limited by short length of follow-up of the implanted scaffolds and relatively weak relevance of the animal model to clinical scenarios such as burn wounds, diabetic foot ulcers, and abdominal wall hernias, etc.
Conclusion
Based on our findings, we conclude that the soluble components of UBM and its own fibrous mesh structure are more conducive to fibroblast adhesion and migration to the center of scaffolds. UBM also provides a microenvironment containing TGF-β1, VEGF and MMP-9 to induce fibroblasts to differentiate into myofibroblasts and continuously reshape ECM.
Supplemental Material
Supplemental Material - Characterizing the inherent activity of urinary bladder matrix for adhesion, migration, and activation of fibroblasts as compared with collagen-based synthetic scaffold
Supplemental Material for Characterizing the inherent activity of urinary bladder matrix for adhesion, migration, and activation of fibroblasts as compared with collagen-based synthetic scaffold by Xiaoyu Tang, Fengbo Yang, Guoping Chu, Xiaoxiao Li, Qiuyan Fu, Mingli Zou, Peng Zhao, and Guozhong Lu in Journal of Biomaterials Applications
Footnotes
Acknowledgements
The authors thank the Social Development Program of Jiangsu Province (BE2018626) and the Special Program of Translational Medicine of Health Committee of Wuxi City (ZM005) for support of this work.
Author contributions
XYT and PZ wrote the main manuscript text, FBY, GPC and XXL, QYF participated in the experiments and analyzed the data, GZL and PZ controlled the direction of the entire research.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.