
Abstract
Keywords
Introduction
Enterococcus faecalis is frequently detected in failed root canal treatments and persistent apical periodontitis, with prevalence ranging from 24% to 77% in such infections. 1 This organism represents a significant challenge in endodontics due to its remarkable ability to survive in harsh root canal environments. 2 The resistance of E. faecalis to eradication is primarily attributed to its ability to form highly organized biofilm structures within the root canal system. 3 Biofilm-embedded E. faecalis exhibits tolerance to conventional disinfectants exceeding 100-fold, resulting from the physical barrier of the extracellular polysaccharide matrix and coordinated defense mechanisms mediated by quorum sensing (QS) systems. 5
Current clinical root canal disinfection protocols rely primarily on sodium hypochlorite irrigation combined with calcium hydroxide medication. 6 However, these approaches have notable limitations. Sodium hypochlorite causes tissue irritation and exhibits limited penetration into deep dentinal tubules. 7 Additionally, E. faecalis can tolerate the alkaline environment created by calcium hydroxide through upregulation of stress response genes and increased cell surface hydrophobicity. 8 These survival mechanisms allow E. faecalis to persist even after conventional treatment protocols. Therefore, novel and effective adjunctive antimicrobial strategies are urgently needed.
Carbon monoxide (CO), as an endogenous gaseous signaling molecule, has recently demonstrated unique advantages in antibacterial applications. 9 CO exerts anti-inflammatory effects through the mitogen-activated protein kinase pathway 10 and possesses cytoprotective properties in various organ injury models. 11 Carbon monoxide-releasing molecules (CORMs), serving as controlled CO delivery vehicles, exert antibacterial effects by disrupting bacterial respiratory chains and inducing oxidative stress. 12 CORMs target bacterial respiration and global transcriptional regulators, causing extensive modifications in gene expression. 13 CORM-3 ([Ru(CO)3Cl(glycinate)]) is a water-soluble ruthenium carbonyl compound that slowly releases CO under physiological conditions with favorable biocompatibility. Previous studies have confirmed the significant bactericidal activity of CORM-3 against Pseudomonas aeruginosa both in vitro and in animal models.14,15 CORM-2 has been shown to reduce the viability of multidrug-resistant uropathogenic Escherichia coli within biofilms and following host-cell colonization. 16 However, research on the effects of CORMs against E. faecalis remains limited. This gap is significant because E. faecalis differs fundamentally from these Gram-negative organisms. As a Gram-positive persister with a thick peptidoglycan cell wall, intrinsic tolerance to multiple disinfectants, and an fsr quorum sensing system that governs biofilm-specific virulence, its susceptibility to CO cannot be extrapolated from earlier work on P. aeruginosa or E. coli. 5
Based on this background, the present study aimed to systematically evaluate the dual effects of CORM-3 on E. faecalis biofilms, colonization inhibition and mature biofilm eradication—while exploring its molecular mechanisms. We comprehensively assessed its biosafety through in vitro and in vivo experiments to provide experimental evidence for the clinical application of CORMs in endodontic treatment.
Materials and methods
CORM-3 solution preparation and CO release kinetics
CORM-3 was dissolved in PBS to prepare a 10 mM stock solution, freshly prepared before use. Inactivated control (iCORM-3) was obtained by incubating CORM-3 solution at 37°C for 24 h to completely exhaust CO release. 14
CO release kinetics were measured using the deoxymyoglobin assay. 15 Deoxymyoglobin (deoxy-Mb, final concentration 60 μM) was incubated with CORM-3 (final concentration 40 μM) in PBS buffer (pH 7.4) at 37°C. Absorbance changes at 540 nm were continuously monitored by UV spectrophotometry. CO release was calculated using the molar extinction coefficient, and time-CO release curves were plotted to determine the half-release time (t1/2).
Experimental groups
The following experimental groups were established: (1) Control group: PBS treatment; (2) iCORM-3 group: inactivated CORM-3 treatment (400 μM) as vehicle control; (3) CORM-3 groups: low-dose (200 μM) and high-dose (400 μM) for dose-response evaluation. All experiments were independently repeated at least three times.
Mature biofilm eradication assay
E. faecalis ATCC 29212 suspension (1 × 107 CFU/mL) was inoculated into 96-well plates and cultured anaerobically at 37°C for 72 h to establish mature biofilm models. 4 After removing the culture medium, wells were gently washed three times with PBS. Fresh BHI medium containing various concentrations of CORM-3 (0, 200, 400 μM) or iCORM-3 (400 μM) was added, followed by continued incubation for 24 h.
Crystal violet staining quantification
Following treatment, culture medium was discarded and wells were washed three times with PBS. Biofilms were stained with 0.1% crystal violet for 15 min, washed with distilled water, and the dye was dissolved in 95% ethanol. Absorbance was measured at 550 nm (OD550). Eradication rate (%) = (1−OD550 of treatment group/OD550 of control group) × 100%.
XTT metabolic activity assay
After treatment as described above, XTT/menadione working solution (0.2 mg/mL XTT + 25 μM menadione) was added. Plates were incubated at 37°C in the dark for 3 h, and absorbance was measured at 490 nm. Metabolic inhibition rate (%) = (1−OD490 of treatment group/OD490 of control group) × 100%.
Biofilm colonization inhibition assay and SEM observation
Sterile round coverslips (14 mm diameter) were placed in 24-well plates. E. faecalis suspension (1 × 107 CFU/mL) was co-cultured with various concentrations of CORM-3 (0, 200, 400 μM) under anaerobic conditions at 37°C for 24 h to assess the inhibitory effect of CORM-3 on initial bacterial colonization and biofilm formation.
For SEM sample preparation, coverslips were washed with PBS after culture, fixed overnight with 2.5% glutaraldehyde at 4°C, dehydrated through a graded ethanol series, substituted with tert-butanol, and freeze-dried. After gold sputter-coating, samples were observed using scanning electron microscopy (Hitachi SU8010) at 5 kV accelerating voltage. Images were acquired at 2000× and 10,000× magnifications.
RT-qPCR analysis of biofilm-related gene expression
Biofilm bacteria from control and CORM-3-treated groups (400 μM, 24 h) were collected. Total RNA was extracted using TRIzol method, and concentration and purity were assessed by NanoDrop (A260/A280 ratio 1.8-2.0). After reverse transcription to cDNA, real-time quantitative PCR was performed. Target genes included: esp (enterococcal surface protein, mediating initial adhesion), 17 gelE (gelatinase, involved in tissue invasion), 18 fsrB (core component of fsr quorum sensing system), 5 and cylL (cytolysin precursor). 19 23S rRNA served as the internal reference gene, and relative expression was calculated using the 2−ΔΔCt method.
ROS generation and membrane integrity assays
Intracellular reactive oxygen species (ROS) generation and membrane integrity were evaluated in mature E. faecalis biofilms treated for 24 h with PBS (control), iCORM-3 (400 μM), or CORM-3 (200 or 400 μM). For ROS detection, treated bacteria were collected, washed twice with PBS, and incubated with 10 μM 2',7'-dichlorodihydrofluorescein diacetate (DCFH-DA) at 37°C for 30 min in the dark. After washing to remove the unloaded probe, fluorescence intensity was measured using a microplate reader at excitation and emission wavelengths of 488 nm and 525 nm, respectively, and expressed as relative fluorescence units (RFU). To assess membrane damage, culture supernatants were collected after treatment and centrifuged at 12,000 × g for 10 min to remove residual cells. The concentration of DNA released into the supernatant was determined from the absorbance at 260 nm, and the amount of leaked protein was quantified using a bicinchoninic acid (BCA) protein assay kit with bovine serum albumin as the standard. All assays were performed with three independent biological replicates.
Cytocompatibility assessment
HOK and HGE cells were seeded into 96-well plates at 5 × 103 cells/well. After cell attachment, medium was replaced with culture medium containing various concentrations of CORM-3 (0, 100, 200, 400, 800 μM). Cell viability was assessed using CCK-8 assay at 24 h and 48 h. Ten microliters of CCK-8 solution was added to each well, and after 2 h incubation at 37°C, absorbance was measured at 450 nm. Cells treated with 0.1% Triton X-100 served as positive control (complete killing), and untreated cells served as negative control. Relative viability (%) = (OD of treatment group−OD of positive control)/(OD of negative control−OD of positive control) × 100%. According to ISO 10993-5 standard, cell viability >70% indicates no significant cytotoxicity.
Animal experiments
SPF-grade male SD rats (180–220 g, 6–8 weeks old) were randomly assigned to two groups (n = 6/group): control group (intraperitoneal injection of equal volume saline) and CORM-3 group (intraperitoneal injection of CORM-3, 8 mg/kg/day, equivalent to approximately 400 μM plasma concentration). Administration was performed once daily for seven consecutive days. Animal experiments were approved by the institutional ethics committee.
Histopathological examination
After blood collection, rats were euthanized. Heart, liver, spleen, lung, and kidney were immediately harvested and fixed in 4% paraformaldehyde. Tissues were paraffin-embedded, sectioned at 4 μm, and stained with hematoxylin-eosin (H&E). Morphological changes were observed under light microscopy.
Complete blood count analysis
Twenty-four hours after the final administration, blood was collected from the abdominal aorta under isoflurane anesthesia and anticoagulated with EDTA-K2. An automated hematology analyzer was used to measure white blood cell count (WBC), red blood cell count (RBC), hemoglobin (HGB), and platelet count (PLT).
Statistical analysis
All experiments were independently repeated at least three times. Data are presented as mean ± standard deviation (x̄ ± s). Statistical analysis was performed using GraphPad Prism 9.0. Comparisons between two groups were analyzed by independent samples t-test, and comparisons among multiple groups were analyzed by one-way ANOVA. p < 0.05 was considered statistically significant.
Results
CO release characteristics of CORM-3
The chemical structure of CORM-3 is shown in Figure 1(A). Its molecular formula is [Ru(CO)3Cl(glycinate)], a water-soluble ruthenium carbonyl compound. The ruthenium atom (Ru) coordinates with three carbonyl groups (CO), which gradually dissociate under physiological conditions to release CO.
12
The activation mechanism of CORM-3 primarily depends on hydrolysis. In aqueous environments, the chloride and glycinate ligands are progressively displaced, destabilizing the Ru-CO bonds and triggering CO release. CO release kinetics were measured using the deoxymyoglobin assay (Figure 1(B)). In PBS buffer at 37°C and pH 7.4, CORM-3 (40 μM) incubated with deoxymyoglobin (60 μM) showed continuous CO release and binding to form carboxymyoglobin (MbCO). CO release exhibited rapid initial kinetics followed by gradual deceleration. Release reached 12.8 ± 1.6 μM at 1 min, 30.4 ± 2.5 μM at 5 min, and plateaued at 32.8 ± 2.7 μM by 10 min, with a half-release time (t1/2) of approximately 1.5 min. This rapid onset with sustained release pattern is advantageous for rapidly establishing effective CO concentrations at the treatment site.
14
In contrast, iCORM-3 released only 1.36 ± 0.16 μM under identical conditions at 10 min, confirming complete loss of CO-releasing activity and validating its use as a negative control. Structure and CO release characteristics of CORM-3. (A) Schematic diagram of CO generation from CORM-3 decomposition. (B) CO release kinetics curve at 37°C and pH 7.4.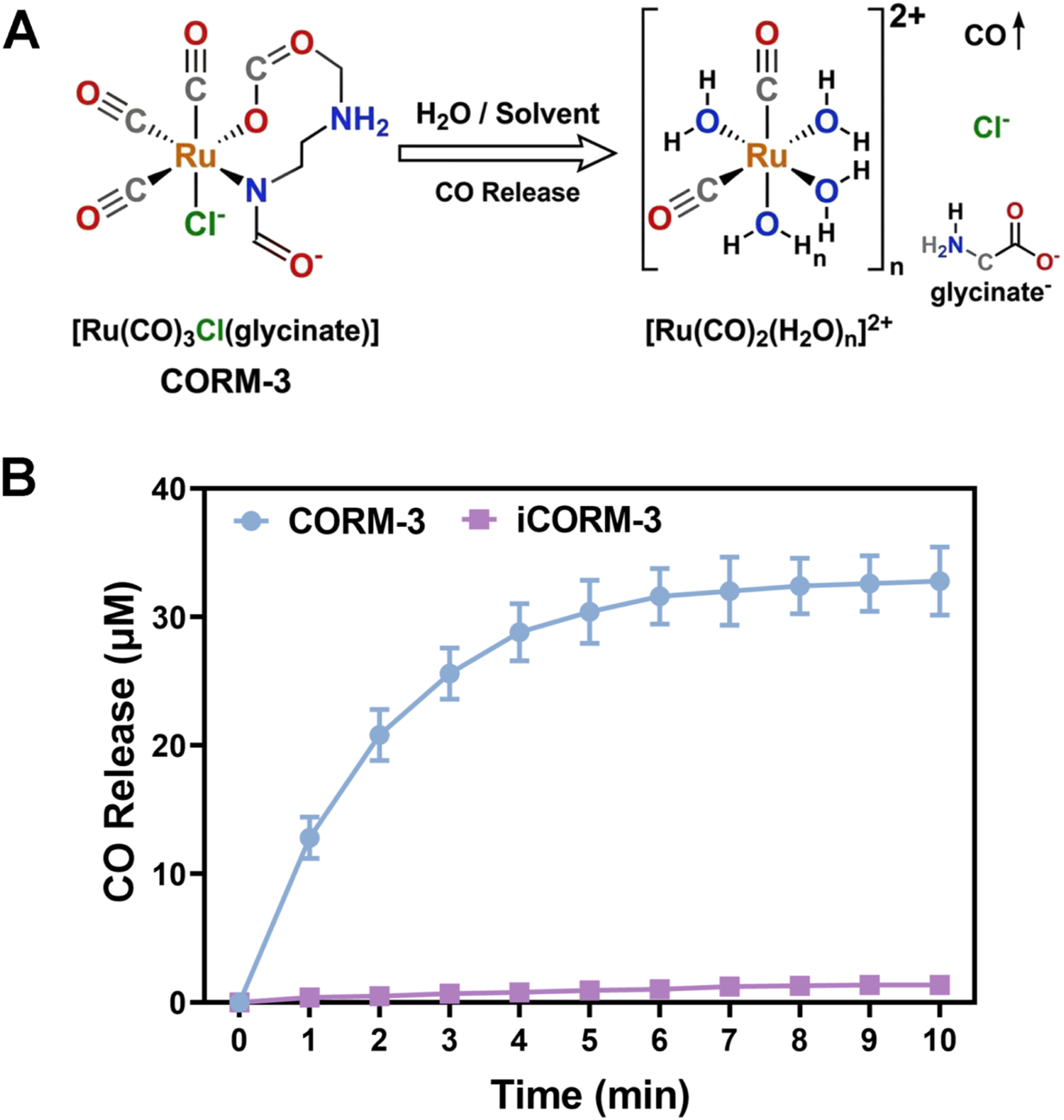
CORM-3 eradicates mature E. faecalis biofilms
To address the more clinically challenging mature biofilms, we constructed 72-h E. faecalis biofilm models and evaluated CORM-3 eradication efficacy.
4
Crystal violet staining visually demonstrated dense dark-purple staining in control and iCORM-3 groups, while CORM-3-treated groups showed markedly reduced staining in a dose-dependent manner (Figure 2(A)). Quantitative analysis revealed no significant difference between iCORM-3 (400 μM) and control groups, confirming that the antibacterial activity of CORM-3 originates from released CO rather than the ruthenium carrier (Figure 2(B)). After 24 h treatment with 200 μM CORM-3, mature biofilm eradication rate reached 42.8 ± 5.8%, increasing significantly to 65.3 ± 4.7% at 400 μM. XTT metabolic activity results were consistent with crystal violet quantification (Figure 2(C)). The iCORM-3 group showed no significant difference in metabolic activity compared to control. Treatment with 200 μM CORM-3 reduced residual biofilm metabolic activity by 38.5 ± 6.2%, while 400 μM treatment achieved 61.7 ± 5.3% metabolic inhibition. These results indicate that CORM-3 not only physically disrupts biofilm structure but also effectively kills or inhibits the metabolic activity of surviving bacteria within the biofilm. CORM-3 eradicates mature E. faecalis biofilms. (A) Crystal violet staining showing eradication effects on 72-h mature biofilms in different groups. (B) Quantitative analysis of residual biofilm mass by crystal violet assay. (C) XTT assay measuring metabolic activity of residual biofilms. *p < 0.05, **p < 0.01; ns indicates no significant difference.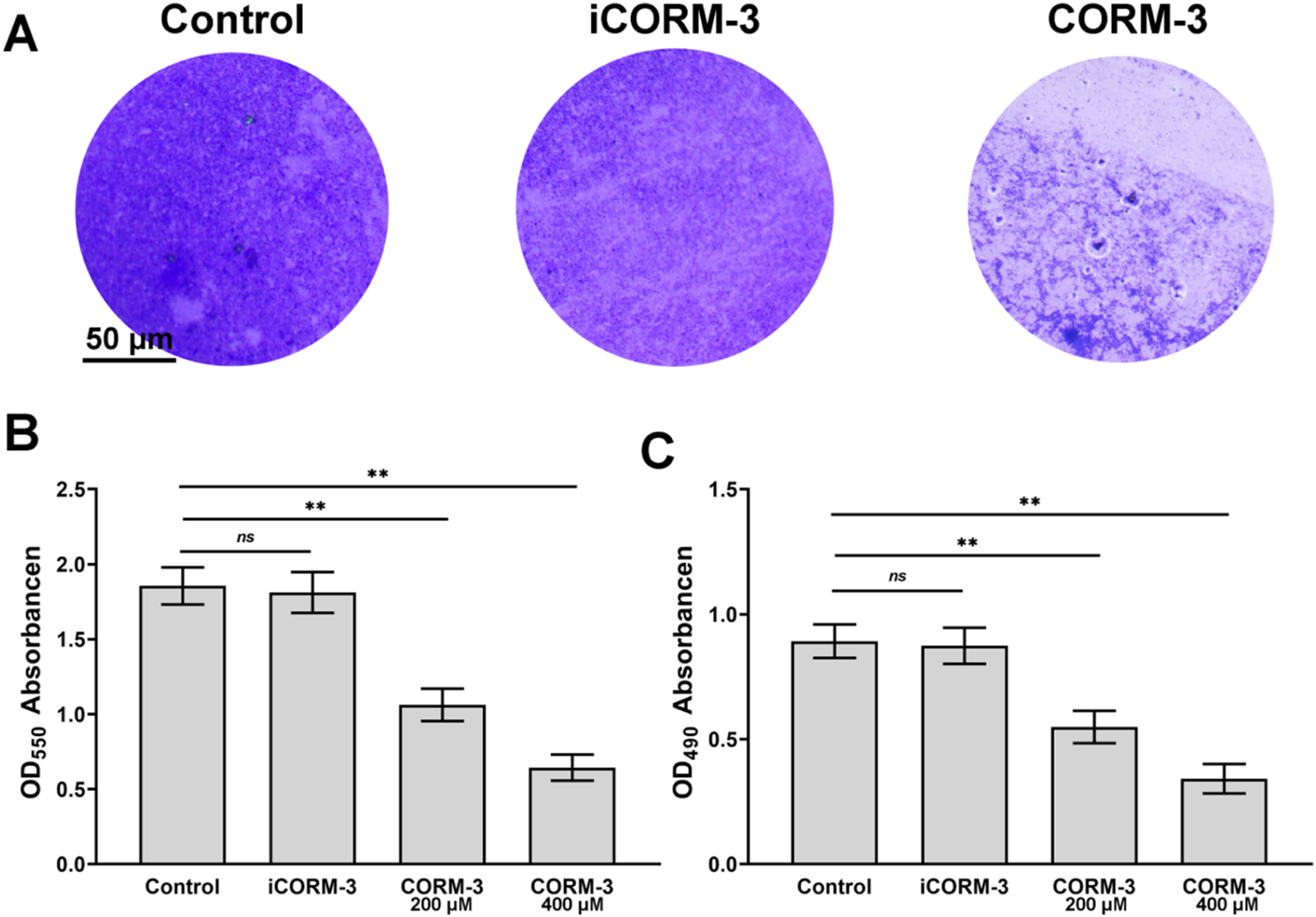
CORM-3 inhibits E. faecalis biofilm colonization
SEM observation directly revealed the inhibitory effect of CORM-3 on E. faecalis initial colonization and biofilm formation (Figure 3(A)). In the control group, abundant E. faecalis adhered to coverslip surfaces. Bacteria were tightly connected by extensive extracellular matrix, forming dense three-dimensional biofilm network structures. Following CORM-3 treatment, bacterial adhesion was substantially reduced. Only scattered individual bacteria or small clusters were observed, with virtually no organized biofilm structure formation. Additionally, some bacteria exhibited morphological alterations including surface wrinkling and irregular depressions, suggesting compromised cell membrane integrity. These results demonstrate that CORM-3 effectively inhibits initial E. faecalis adhesion to substrate surfaces and subsequent biofilm colonization. CORM-3 inhibits E. faecalis biofilm colonization and downregulates related gene expression. (A) SEM observation of E. faecalis colonization on coverslip surfaces after 24-h treatment with various CORM-3 concentrations. (B) RT-qPCR analysis of relative expression changes of esp, gelE, fsrB, and cylL genes after 24-h treatment with 400 μM CORM-3. *p < 0.05, **p < 0.01.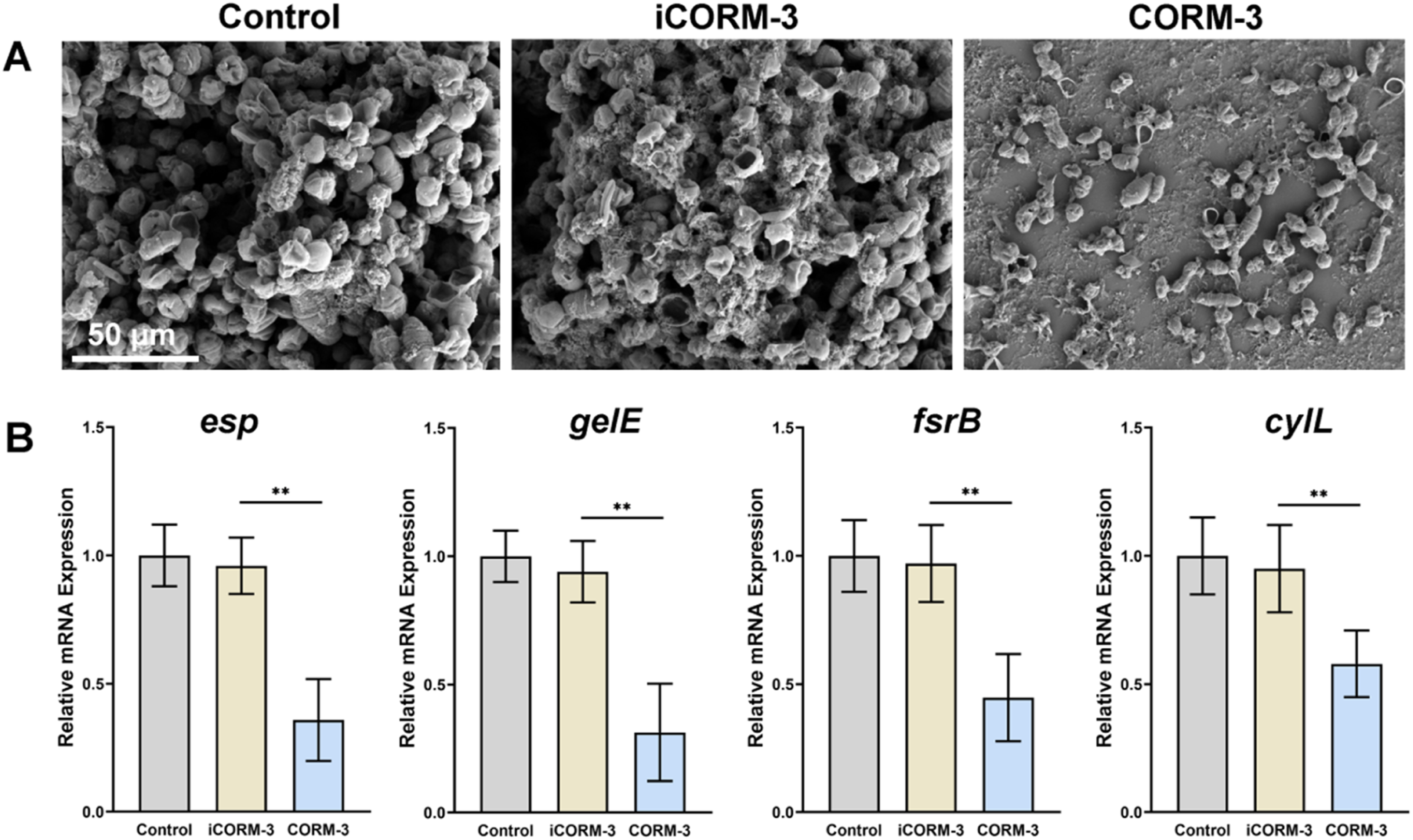
CORM-3 downregulates biofilm-related gene expression
To elucidate the molecular mechanisms underlying CORM-3 anti-biofilm activity, we analyzed biofilm-related gene expression changes by RT-qPCR (Figure 3(B)). Results showed significant downregulation of multiple key virulence genes in E. faecalis after 24 h CORM-3 treatment. Expression of esp was downregulated by 64.2 ± 7.5%, gelE by 68.7 ± 5.8%, fsrB by 55.3 ± 8.1%, and cylL by 42.1 ± 9.3%. These findings indicate that CORM-3 simultaneously inhibits E. faecalis adhesion capacity (esp), extracellular matrix metabolism (gelE), and the quorum sensing system (fsrB) at the transcriptional level, thereby blocking biofilm colonization and maturation through multiple pathways.5,17,20
CORM-3 induces ROS generation and compromises membrane integrity
To further elucidate the antibacterial mechanism of CORM-3, we examined intracellular ROS accumulation and bacterial membrane integrity in mature biofilms following treatment (Figure 4). Intracellular ROS levels detected by the DCFH-DA fluorescence assay increased in a dose-dependent manner after CORM-3 treatment (Figure 4(A)). Relative to the control group (1398 ± 172 RFU), ROS fluorescence rose to 3085 ± 298 RFU at 200 μM and 4592 ± 412 RFU at 400 μM, corresponding to approximately 2.2- and 3.3-fold increases, respectively, whereas the iCORM-3 group (1487 ± 195 RFU) showed no significant difference from the control. Membrane damage was assessed by quantifying the leakage of intracellular contents into the culture supernatant. Extracellular DNA leakage increased from 7.1 ± 1.0 μg/mL in the control group to 23.8 ± 3.1 μg/mL at 200 μM and 37.2 ± 4.2 μg/mL at 400 μM CORM-3 (Figure 4(B)), and protein leakage rose correspondingly from 19.2 ± 2.5 μg/mL to 64.8 ± 6.7 μg/mL and 101.5 ± 8.9 μg/mL (Figure 4(C)). For both indicators, the iCORM-3 group remained comparable to the control, confirming that these effects originate from released CO rather than the ruthenium carrier. Collectively, these findings demonstrate that CORM-3 induces intracellular oxidative stress and compromises membrane integrity, resulting in leakage of cellular contents and bacterial death. Together with the transcriptional downregulation of biofilm-related genes, this mechanism accounts for the antibacterial and anti-biofilm activity of CORM-3 against E. faecalis (Figure 4(D)). CORM-3 induces ROS generation and compromises bacterial membrane integrity in E. faecalis. Mature E. faecalis biofilms were treated for 24 h with PBS (Control), iCORM-3 (400 μM), or CORM-3 (200 or 400 μM). (A) Intracellular ROS levels detected by DCFH-DA fluorescence assay. (B) Extracellular DNA leakage into the culture supernatant. (C) Protein leakage into the supernatant quantified by BCA assay. (D) Antibacterial mechanism of CORM-3 against E. faecalis. *p < 0.05, **p < 0.01, ***p < 0.001; ns indicates no significant difference.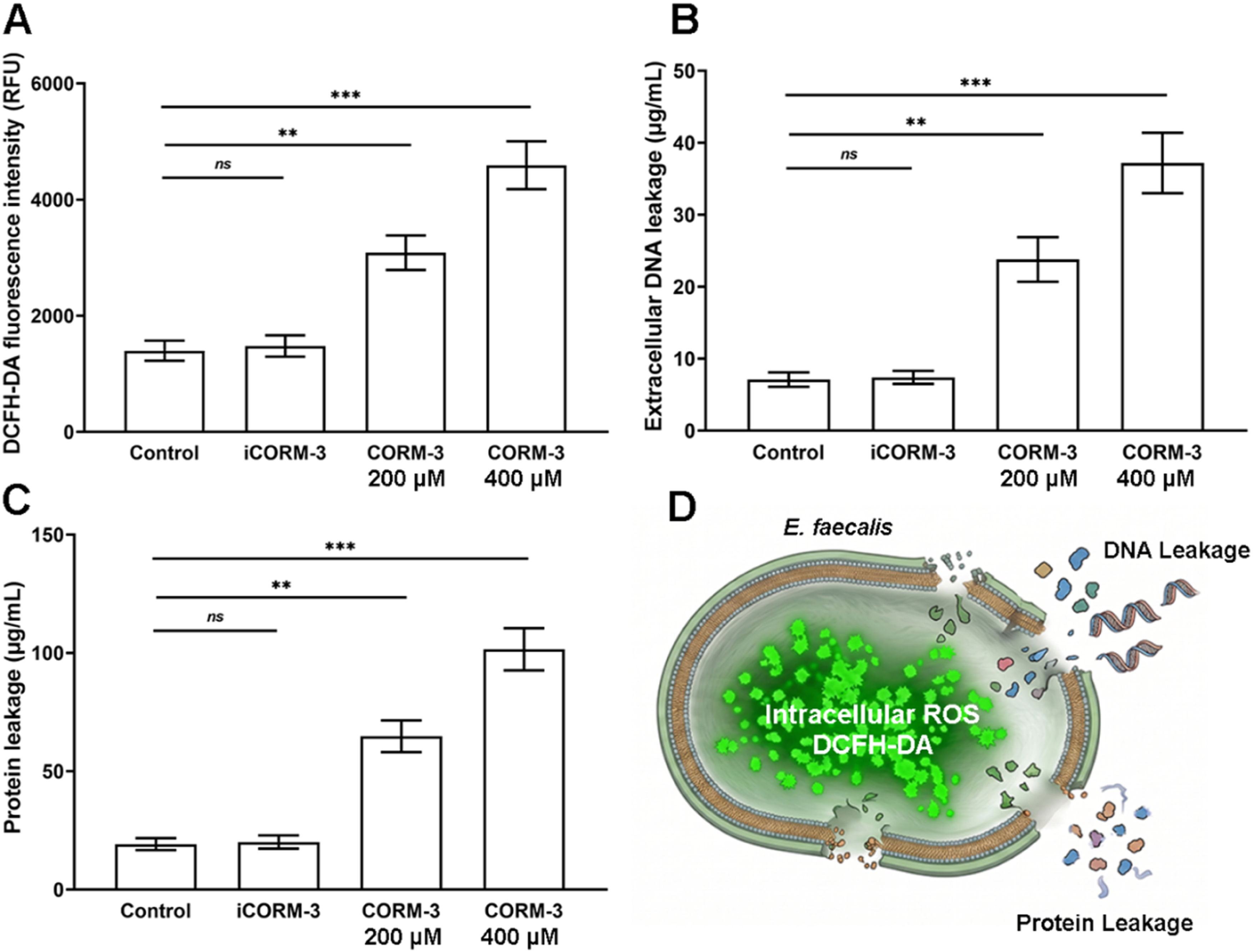
Cytocompatibility of CORM-3
To evaluate CORM-3 safety for oral tissue cells, we assessed its cytotoxicity to HOK and HGE cells using CCK-8 assay (Figure 5). For HOK cells (Figure 5(A) and (B)), cell viability after 24 h treatment with 100, 200, 400, and 800 μM CORM-3 was 96.8 ± 2.6%, 93.4 ± 3.4%, 87.3 ± 3.6%, and 68.5 ± 5.2%, respectively. At 48 h, viability was 94.5 ± 3.0%, 90.7 ± 3.5%, 83.6 ± 4.1%, and 61.2 ± 6.4%, respectively. For HGE cells (Figure 5(C) and (D)), cell viability after 24 h treatment at corresponding concentrations was 97.2 ± 2.1%, 94.6 ± 2.9%, 89.1 ± 4.3%, and 70.3 ± 6.7%, respectively. At 48 h, viability was 95.3 ± 2.6%, 91.8 ± 3.3%, 85.2 ± 4.7%, and 63.4 ± 6.9%, respectively. According to ISO 10993-5 standard, CORM-3 demonstrated favorable biocompatibility toward both oral cell types at concentrations ≤400 μM. Notably, 400 μM represents the optimal anti-biofilm concentration in this study, indicating acceptable cellular safety at effective antibacterial concentrations. Cytocompatibility of CORM-3. CCK-8 assay measuring cell viability after treatment with various CORM-3 concentrations. (A) HOK cells at 24 h. (B) HOK cells at 48 h. (C) HGE cells at 24 h. (D) HGE cells at 48 h. Dashed line indicates the 70% viability threshold.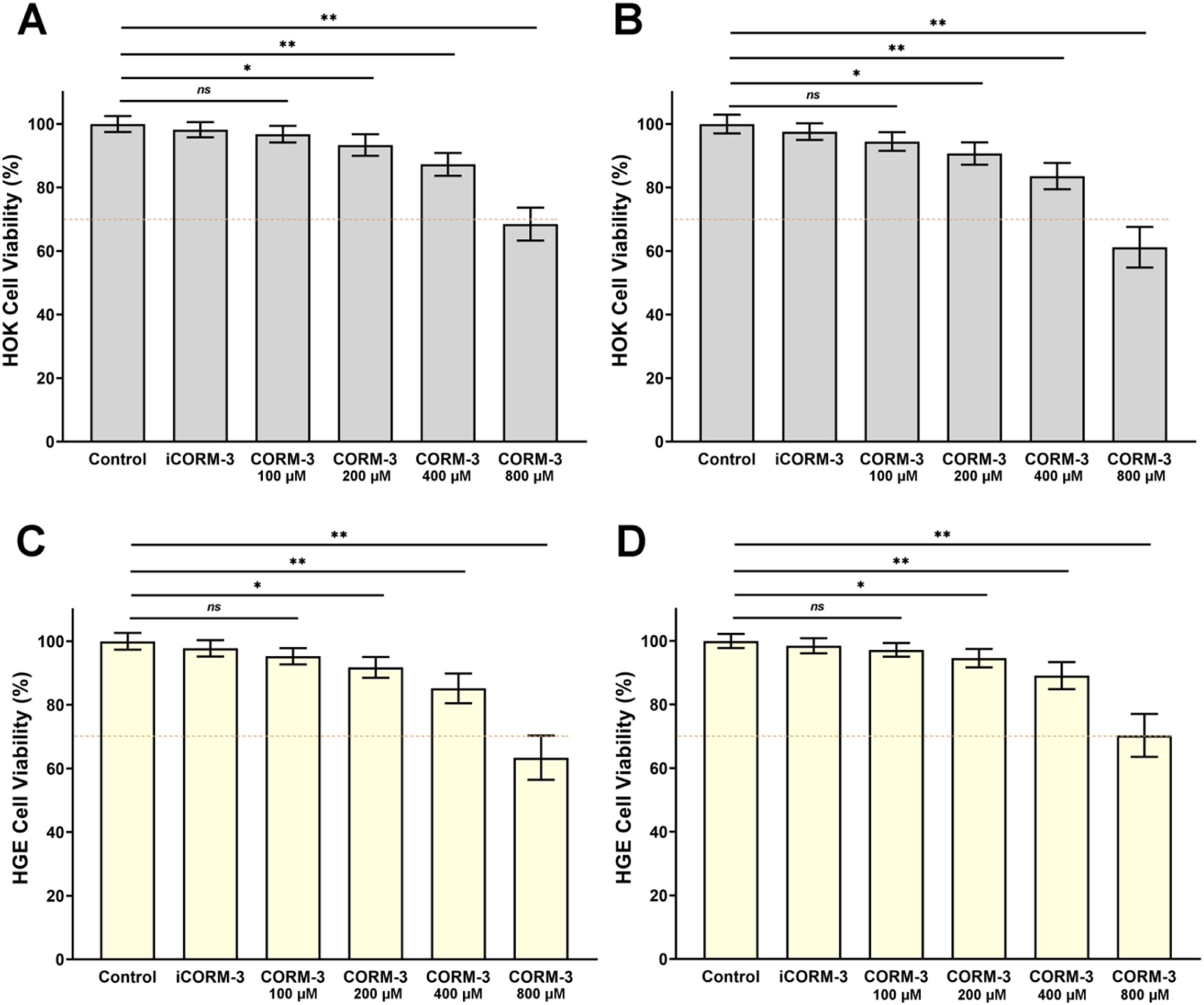
In vivo safety evaluation of CORM-3
To further evaluate CORM-3 systemic toxicity, acute toxicity experiments were performed in SD rats (Figure 6). Histopathological examination revealed no obvious pathological changes in H&E-stained sections of heart, liver, spleen, lung, and kidney from both control and CORM-3 groups (Figure 6(A)). Specifically, cardiac muscle fibers were regularly arranged, hepatocyte cords showed normal organization, splenic red and white pulp structures were preserved, alveolar structures remained intact, and glomerular morphology appeared normal. Complete blood count analysis showed no significant differences in WBC, RBC, HGB, and PLT between CORM-3-treated rats (8 mg/kg/day for 7 days) and control group (Figure 6(B)), indicating that CORM-3 at this dose does not cause significant hematological toxicity. These results demonstrate that CORM-3 exhibits no apparent toxic damage to major organs in rats at the experimental dose and possesses favorable in vivo safety. In vivo safety evaluation of CORM-3. (A) H&E-stained sections of major organs from SD rats after 7 days of intraperitoneal CORM-3 administration. (B) Complete blood count results including white blood cell count (WBC), red blood cell count (RBC), hemoglobin (HGB), and platelet count (PLT). ns indicates no significant difference.
Discussion
E. faecalis biofilm represents a key factor in root canal treatment failure.1,2 Its high tolerance to conventional disinfectants has driven ongoing exploration of novel antibacterial strategies. 3 This study is the first to systematically evaluate the dual antibacterial activities of CORM-3 against E. faecalis biofilms, colonization inhibition and mature biofilm eradication, while comprehensively assessing its biosafety through in vitro and in vivo experiments. The conceptual advance here lies not in re-demonstrating that CO is antibacterial, which is already established, but in showing that a single agent can both prevent initial colonization and eradicate established biofilms of a Gram-positive endodontic persister, a dual capability not previously documented for CORMs, and in linking this action to the coordinated transcriptional suppression of adhesion, matrix, and quorum-sensing determinants. Earlier CORM studies in Gram-negative pathogens established bactericidal activity together with broad disruption of respiration and global transcriptional regulation,13,16 but did not resolve the biofilm-specific regulatory determinants targeted here.
First, as a controlled CO release carrier, the antibacterial action of CORM-3 depends on its CO release kinetics. Our study demonstrated a half-release time of approximately 1.5 min under physiological conditions, with CO release reaching 32.8 μM and plateauing within 10 min. This rapid-onset release pattern offers unique advantages for root canal treatment. The root canal system is a relatively closed anatomical space where conventional liquid disinfectants have difficulty penetrating deep dentinal tubules, isthmuses, and lateral canals. 7 As a small molecule gas, CO exhibits excellent tissue penetration. It can freely diffuse through biofilm matrix and dentinal tubules to reach areas inaccessible to liquid disinfectants. Furthermore, the enclosed nature of the root canal system facilitates local CO accumulation, prolonging contact time with biofilms and thereby enhancing antibacterial efficacy. 14
Second, our results demonstrate that CORM-3 exerts dual effects on E. faecalis biofilms, effectively inhibiting bacterial colonization and eradicating mature biofilms. In root canal retreatment cases, the core clinical challenge is eliminating established persistent biofilms in the root canal system. Preventing iatrogenic bacterial recolonization during initial or retreatment procedures is equally important. The antibacterial mechanism of CO differs from traditional antibiotics. CO primarily binds to terminal cytochrome oxidases in the bacterial respiratory chain, blocking electron transport and oxidative phosphorylation while inducing intracellular ROS accumulation and oxidative stress damage.12,13 This multi-target mechanism may explain why CORM-3 effectively kills biofilm-embedded bacteria.
When benchmarked against established endodontic irrigants, the eradication achieved here (65.3% at 400 μM) is lower than that typically reported for sodium hypochlorite at clinical concentrations (2.5% to 5.25%), which remains the most effective antibiofilm irrigant. The efficacy of sodium hypochlorite, however, is markedly attenuated by organic matter such as dentin and pulpal remnants, its penetration into dentinal tubules is limited, 7 and it offers no sustained activity after irrigation while remaining cytotoxic to periapical tissues. Chlorhexidine, although better tolerated, lacks tissue-dissolving capacity and shows reduced efficacy against mature biofilms. CORM-3 is therefore positioned not as a replacement for these agents but as a biocompatible adjunct whose gaseous CO can diffuse into regions that liquid irrigants penetrate poorly, complementing rather than substituting for conventional irrigation.
Additionally, RT-qPCR results revealed the molecular mechanisms underlying CORM-3 anti-biofilm activity. The fsr quorum sensing system is the most important density-dependent regulatory network in E. faecalis.5,18 It coordinates downstream virulence gene expression through secretion and sensing of the autoinducing peptide GBAP (gelatinase biosynthesis-activating pheromone). 20 Our finding that CORM-3 significantly downregulated fsrB expression (55.3%) suggests that CO may inhibit biofilm maturation and synchronized virulence factor expression by interfering with quorum sensing signal transduction. 19 The esp gene encodes enterococcal surface protein (Esp), a key adhesin mediating E. faecalis attachment to host cells and abiotic material surfaces. 17 Its N-terminus contains multiple repeat domains that specifically bind host extracellular matrix proteins. The 64.2% downregulation of esp expression and the significantly reduced bacterial adhesion observed by SEM form a complete evidence chain from gene to phenotype. The gelE gene encodes gelatinase, a metalloproteinase involved not only in extracellular matrix degradation and remodeling but also in biofilm formation regulated by the fsr system.18,21 Its 68.7% downregulation suggests that CORM-3 may simultaneously impair E. faecalis tissue invasiveness, potentially preventing periapical infection spread. Because these analyses are correlative, the observed transcriptional changes may reflect either a direct action of CO on redox-sensitive transcriptional regulators or a secondary consequence of respiratory inhibition and oxidative stress, a distinction that the present data cannot resolve.
Safety is a critical prerequisite for CORM-3 clinical translation. Notably, CO is not purely a toxic molecule in mammalian cells. Appropriate concentrations of exogenous CO have been shown to possess anti-inflammatory, anti-apoptotic, and tissue repair-promoting biological activities.10,11 In this study, at the effective antibacterial concentration (400 μM), CORM-3 maintained cell viability above 87% for both HOK and HGE cells at 24 h and above 83% at 48 h, in all cases well above the 70% safety threshold specified by ISO 10993-5. In vivo experiments further confirmed that hematological parameters and major organ pathology remained normal after administration. This safety profile is particularly important for root canal treatment applications. The apical foramen directly communicates with periapical tissues, and any intracanal medication may affect surrounding tissues. The CO released by CORM-3 may provide protective effects on inflamed periapical tissues while exerting antibacterial action, 10 a hypothesis warranting further investigation.
This study has certain limitations. First, experiments used the reference strain ATCC 29212, whereas clinical isolates often harbor more virulence genes, exhibit stronger biofilm-forming capacity, and show greater phenotypic heterogeneity. 4 Future studies should include multiple clinical isolates to validate the broad-spectrum antibacterial activity of CORM-3. Second, in vitro experimental conditions cannot fully simulate the complex root canal microenvironment, including the three-dimensional anatomy of dentinal tubules, the hypoxic and slightly acidic intracanal environment, and clinically common polymicrobial infection patterns. 3 Finally, this study focused on CORM-3 application alone. The combined effects with existing root canal disinfection protocols and potential synergistic mechanisms warrant further exploration, which may provide new insights for developing more effective root canal disinfection strategies. In addition, although the combination of transcriptomic and phenotypic data provides convergent evidence, protein-level validation of the affected virulence factors was not performed, as validated antibodies for several of these targets remain limited in availability. More fundamentally, the present work is a proof-of-concept study. A clinically usable delivery vehicle for CORM-3 has not yet been defined, and its retention, effective dosing, and CO release kinetics within the anatomically complex root canal, together with its compatibility with endodontic sealers and irrigants, remain to be characterized. These translational questions must be resolved before clinical application can be considered.
Conclusion
This study demonstrates that CORM-3 exerts dual antibacterial activities against E. faecalis biofilms through controlled CO release: inhibiting initial bacterial colonization and biofilm formation, thereby preventing bacterial recolonization, while eradicating established mature biofilms. The molecular mechanism involves downregulation of quorum sensing system (fsrB), adhesion-related gene (esp), and extracellular enzyme gene (gelE) expression. Both in vitro cell experiments and in vivo animal studies confirm that CORM-3 exhibits favorable biosafety at effective antibacterial concentrations. These findings provide proof-of-concept evidence supporting the further development of CORM-3 as an adjunctive antimicrobial agent in endodontic treatment. Clinical translation will require defining a suitable delivery system and evaluating its retention, dosing, and compatibility with existing materials within the root canal environment.
Footnotes
Ethical considerations
This study was approved by the Institutional Animal Care and Use Committee of Sichuan University. All animal procedures strictly followed the Guidelines for the Care and Use of Laboratory Animals.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.