
Abstract
Objective
Fatigue is among the most common complaints in community-dwelling older adults, yet its etiology is poorly understood. Based on models implicating frontostriatal pathways in fatigue pathogenesis, we hypothesized that smaller basal ganglia volume would be associated with higher levels of subjective fatigue and reduced set-shifting in middle-aged and older adults without dementia or other neurologic conditions.
Methods
Forty-eight non-demented middle-aged and older adults (Mage = 68.1, SD = 9.4; MMMSE = 27.3, SD = 1.9) completed the Fatigue Symptom Inventory, set-shifting measures, and structural MRI as part of a clinical evaluation for subjective cognitive complaints. Associations were examined cross-sectionally.
Results
Linear regression analyses showed that smaller normalized basal ganglia volumes were associated with more severe fatigue (β = −.29, P = .041) and poorer Trail Making Test B-A (TMT B-A) performance (β = .30, P = .033) controlling for depression, sleep quality, vascular risk factors, and global cognitive status. Putamen emerged as a key structure linked with both fatigue (r = −.43, P = .003) and TMT B-A (β = .35, P = .021). The link between total basal ganglia volume and reduced TMT B-A was particularly strong in clinically fatigued patients.
Conclusion
This study is among the first to show that reduced basal ganglia volume is an important neurostructural correlate of subjective fatigue in physically able middle-aged and older adults without neurological conditions. Findings suggest that fatigue and rapid set-shifting deficits may share common neural underpinnings involving the basal ganglia, and provide a framework for studying the neuropathogenesis and treatment of subjective fatigue.
Introduction
Fatigue is among the most widely reported complaints among community-dwelling adults in the U.S. aged 50 years and older, with an estimated prevalence of 31.2%. 1 Among non-disabled older adults, self-reported fatigue is associated with increased risk for mood disorders, 2 decreased quality of life, 3 functional impairment, 4 future onset of disability, 5 and mortality. 6 Additionally, fatigue is the strongest predictor of increased healthcare costs for aging adults compared to other Frailty Syndrome symptoms. 7
There is also mounting literature linking heightened fatigue ratings to reduced neurocognitive functioning in older adults, even in the absence of neurological conditions and independent of depression and sleep quality.8,9 Specifically, fatigue and cognitive decline have been shown to progress in tandem over 5 years in a large national sample, 8 and a recent community-based study found that fatigue was inversely associated with a frontal-subcortical pattern of neuropsychological deficits including executive functioning, attention, and psychomotor speed. 9 At present, the nature of this association remains poorly understood.
While fatigue in older adults is often discussed in the context of depression, insomnia, and medical illnesses, 10 there is increasing evidence that fatigue may be etiologically distinct and conceptualized separately from these conditions.11,12 In fact, an early population-based study showed that nearly 16% of fatigue cases could not be ascribed to a diagnosed medical or psychological condition, 13 highlighting the need to better understand etiological drivers of fatigue in older adults.
There is growing recognition that the basal ganglia, comprising interconnected subcortical nuclei including the striatum (caudate and putamen) and globus pallidus (internal and external segments), are involved in non-motor functions. 14 Chaudhuri and Behan’s 15 leading neural model of fatigue posits a central role of the basal ganglia in fatigue etiology in which basal ganglia degeneration (or dysfunction) is thought to disrupt striato-cortical and striato-thalamo-cortical dopaminergic pathways involved in initiating and sustaining self-motivation, internal cueing for goal-directed tasks, and regulating energy expenditure and perception. 15 Consistent with this model, neuroimaging studies have routinely linked diminished basal ganglia structure and function with higher levels of fatigue in clinical populations including multiple sclerosis, Parkinson’s disease, traumatic brain injury, and chronic fatigue syndrome.15-17
By contrast, a recent comprehensive review 18 identified only 4 studies investigating structural neural correlates of fatigue in older adults without neurological conditions, of which only 2 utilized validated self-report measures of fatigue.19,20 In a control group of healthy older adults, the first study 19 did not find a difference in basal ganglia volumes between high- and low-fatigue participants. Conversely, in a study of sedentary septuagenarians and octogenarians with poor physical functioning, the other 20 found that reduced gray-matter volumes of basal ganglia and limbic structures were associated with higher levels of fatigability. Additional studies are needed to elucidate neurostructural correlates of fatigue in older adults without frank neurologic conditions, 18 and to examine whether the basal ganglia model of fatigue 15 may extend to this population.
Striatofrontal dysfunction is a key pathogenic feature of the basal ganglia model of fatigue thought to accompany basal ganglia degeneration. 15 Set-shifting, the ability to fluidly alternate between cognitive sets, is a core executive function dependent on striatal modulation of prefrontal systems that can provide a behavioral index of striatofrontal function. 21 Accordingly, measures of set-shifting have been shown to be sensitive to striatal degeneration 22 and hypoactivation. 23 It is therefore reasonable to propose that clinically fatigued individuals may exhibit an association between basal ganglia atrophy and deficits in set-shifting, though this is yet to be examined.
In this cross-sectional study, we first examined structural basal ganglia correlates of fatigue in a sample of community-dwelling middle-aged and older adults without major psychiatric, cognitive, neurological, or medical conditions commonly tied with fatigue. Drawing on the basal ganglia model of fatigue, 15 we hypothesized that smaller total basal ganglia volumes would be associated with higher levels of fatigue while controlling for psychosocial factors commonly conflated with fatigue (i.e., older age, depressive symptoms, and poor sleep quality) and global cognitive status. We also explored individual basal ganglia sub-structure volumes, as it is not known whether specific basal nuclei differentially relate to fatigue pathogenesis. Second, we tested the hypothesis that smaller total basal ganglia volumes would be associated with poorer cognitive set-shifting, particularly among subjects reporting clinically elevated symptoms of fatigue.
Methods
Participants
Participants were selected from the McKnight Brain Institute Registry at the University of Miami Miller School of Medicine in Miami, Florida. Details about the registry have been published elsewhere. 9 Briefly, the registry consisted primarily of patients aged 50 years and older evaluated in the Department of Neurology for subjective memory/cognitive complaints (e.g., forgetting recent conversations, word-finding difficulties, and losing one’s train of thought) and referred for neuropsychological evaluation by their neurologist. Patients were considered for the study if they completed a neuropsychological evaluation involving a global cognitive screen and set-shifting measures, a self-report measure of fatigue, and structural magnetic resonance imaging (MRI) with T1-weighted magnetization-prepared rapid gradient echo (MPRAGE) or spoiled gradient recalled echo (SPGR) assessment with maximum voxel size of 1 mm × 1 mm × 1 mm within 1 year of the neuropsychological evaluation. There were 257 patients in the registry who completed cognitive and fatigue measures. Of these, 59 had a recent MPRAGE or SPGR MRI of acceptable quality within the allotted time frame.
Exclusion Criteria.

Procedures
As part of a clinical evaluation, participants completed neuropsychological measures and self-report questionnaires assessing fatigue, mood, and sleep-related problems. All measures were administered under the supervision of a licensed neuropsychologist. Participants received a clinical neurologic evaluation by a neurologist and completed brain MRI. All participants were unpaid volunteers who provide written informed consent for enrollment in the registry. Exclusionary conditions were identified by review of electronic medical records by trained employees or doctoral-level students.
Study procedures were approved by the University of Miami Central Institutional Review Board. Participants were assessed between March of 2016 and March of 2019.
Measures
Subjective fatigue. The Fatigue Symptom Inventory (FSI) 24 is a validated self-report instrument with acceptable psychometric properties 25 assessing subjective fatigue symptoms over the past week. The FSI-severity scale composite used in this study is derived by calculating the average across 3 items assessing one’s subjective fatigue severity (1) on average in the past week; (2) on the day they felt most fatigued in the past week; and (3) on the day they felt least fatigued in the past week, rated on 11-point Likert-type scales (0 = not at all fatigued, 10 = as fatigued as I could be). 24 A FSI-severity scale cutoff score of ≥ 3 is recommended for identifying clinically elevated fatigue. 25
Depression symptoms and sleep quality. Depressive symptoms were assessed with the Beck Depression Inventory-II. 26 Total scores were classified as minimal (0-13), mild (14-19), moderate (20-28), or severe (29-63) depression. Sleep quality was assessed with the Pittsburgh Sleep Quality Index (PSQI) 27 final composite score. These measures are described in greater detail elsewhere. 9
Dementia screening. The Mini-Mental State Examination (MMSE) 28 is a brief screening tool for assessing global cognitive impairment. Participants with scores < 24/30 were excluded in order to minimize the likelihood of including patients with frank dementia based on clinical recommendations. 29
Cognitive set-shifting. The Trail Making Test (TMT) and Wisconsin Card Sorting Test (WCST) are well validated and commonly used measures for assessing set-shifting abilities and frontal-lobe dysfunction that are described in detail elsewhere. 30 Both tests were administered in-person in accordance with manual testing procedures. The TMT is a timed paperpencil task in requiring the examinee to connect circles on a page in numerical sequence (part A) before alternating between letter-number sets in ascending order (part B). The TMT B-A difference score was used to assess rapid set-shifting ability; subtracting part A completion time from part B is thought to remove visuomotor processing speed and working memory demands associated with part A, yielding a more refined index of executive set-shifting ability. 31 The WCST requires the examinee to manually match cards according to specific, changing rules which he or she must learn, reason through, and adapt to based on limited feedback from the examiner. The WCST perseverative errors score was used to assess untimed set-shifting ability. For both measures, raw scores were converted to normative T-scores (M = 50, SD = 10) based on published age- and education-adjusted norms with higher scores indicating superior performance.
Neuroimaging data processing. MRI data acquisition was conducted for most cases using Siemens scanners (Skyra 3-T n = 22, Trio 3-T n = 18, Avanto 1.5-T n = 4, and Symphony 1.5-T n = 3) equipped with a standard head coil. One image was acquired using a GE Signa 1.5-T. Anatomical scans of the entire brain were collected using T1-weighted MPRAGE or SPGR sequences; MRI acquisition parameters are provided in Supplementary Table 1. Structural images were processed using FreeSurfer Image analysis suite, which is documented and freely available for download online (http://surfer.nmr.mgh.harvard.edu). 32 Image processing involves automated motion correction of volumetric weighted images, computerized removal of non–brain tissue using a hybrid watershed/surface deformation procedure, intensity normalization, tessellation of the gray matter/white matter boundary, automated topology correction, and surface deformation following intensity gradients to optimally place the gray/white and gray/cerebrospinal fluid borders at the location where the greatest shift in intensity defines the transition to the other tissue class. Visible parcellation errors were corrected using control points. Labels for grey-matter volumetric segmentation were assigned to each voxel via a probabilistic atlas and Bayesian classification rule (84 gray matter volumetric segmentations). Volumes of interest (i.e., total basal ganglia volume, caudate, putamen, and globus pallidus) were selected a priori. In order to reduce the number of comparisons, we examined summed bilateral volumes. All volumes of interest are presented as ratios to total intracranial volume (ICV) to control for head size.
Statistical Analyses
Descriptive statistics and data screening. Descriptive statistics and analyses were performed using SPSS software version 24.0. Data were screened for normality and violation of regression assumptions statistically and visually by examining metrics of skewness and kurtosis, histograms, residual plots, and plots of standardized predicted values. An outlier (> 3.0 SD) was identified on the PSQI and subsequently Winsorized. No additional transformations were required. Missing data were deleted listwise (< 7% across variables of interest).
Covariates. In regression analyses, age, depression symptoms, sleep quality, hypertension, and diabetes were selected a priori as covariates because of known effects on fatigue and cognition. Total MMSE scores were included to account for differences global cognitive status. We also included covariates for scanner field strength (0 = 3-T; 1 = 1.5-T) and model (Skyra = 1, Other = 0). Prior to hypothesis testing, neither Hispanic/Latinx ethnicity nor sex was found to be significantly associated with fatigue severity, TMT B-A, WCST, or bilateral basal ganglia volume (P-value range: .30-.95) and therefore, were not included as covariates in the regression models. Education level was not included as a covariate given our use of education-corrected norms for TMT B-A and WCST and because it was not correlated with fatigue severity (r = .06, P = .692).
Basal ganglia volumes and fatigue. Ordinary least squares multiple linear regression analysis was performed to examine the association of total basal ganglia volume (normalized for ICV) with fatigue severity. Total basal ganglia volume, age, scanner field strength, and scanner model were entered in the first block (base model); depressive symptoms, sleep quality, hypertension, diabetes, and MMSE score were added simultaneously in the second block (full covariate model). To quantify the magnitude of a significant direct association, we calculated Cohen’s local effect size f2 values 33 within the full covariate models. Values of .02, .15, and .35 represent small, medium, and large effects, respectively. We also examined squared-semipartial correlations. The P-value was set at .05. We conducted post-hoc bivariate partial correlations, adjusted for age and scanner field strength, scanner model, and MMSE for individual basal ganglia structure volumes with fatigue.
Basal ganglia volume and set-shifting. First, we conducted separate multiple linear regression analyses for total basal ganglia volume with TMT B-A and WCST scores following the procedure described above. Post-hoc bivariate partial correlations were performed for individual basal structure volumes with the 2 set-shifting measures for descriptive purposes. Second, subjects were categorized as clinically elevated fatigue or minimal fatigue based on the FSI-severity cutoff score (≥ 3), 25 and partial bivariate correlations were examined for total basal ganglia volume with set-shifting scores.
Sensitivity analysis. In addition to controlling for field strength and scanner model, we conducted a series of post-hoc analyses to confirm whether findings were robust to differences in field strength and scanner models. These are described in the Supplementary Material.
Results
Sample Characteristics and Descriptive Statistics.
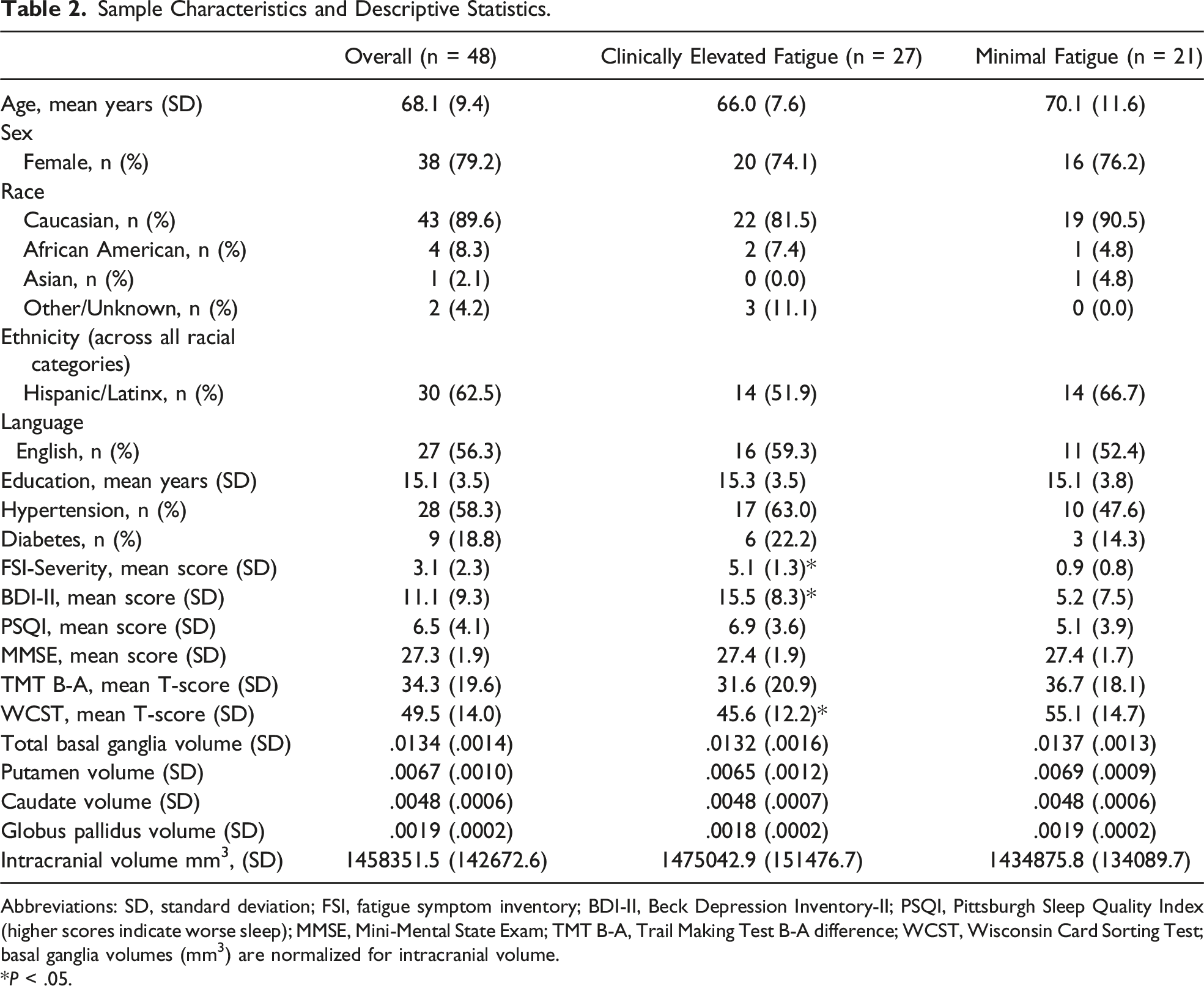
Abbreviations: SD, standard deviation; FSI, fatigue symptom inventory; BDI-II, Beck Depression Inventory-II; PSQI, Pittsburgh Sleep Quality Index (higher scores indicate worse sleep); MMSE, Mini-Mental State Exam; TMT B-A, Trail Making Test B-A difference; WCST, Wisconsin Card Sorting Test; basal ganglia volumes (mm3) are normalized for intracranial volume.
*P < .05.
Descriptive statistics for key study variables are also shown in Table 2. The mean fatigue severity score fell just at the cutoff for clinically elevated fatigue. The mean norm-adjusted T-scores for TMT B-A and WCST fell in the borderline impaired range and average range, respectively; they were not correlated (r = .03, P > .05). Fatigue was not significantly correlated with TMT B-A (r = −.15, P > .05). There were small-to-medium sized correlations among bilateral basal ganglia structures (putamen with caudate: r = .35, P = .014; putamen with pallidum: r = .60, P < .001) with the exception of caudate and pallidum (r = −.23, P = .106).
Total Basal Ganglia Volume and Fatigue
Standardized Regression Coefficients for Total Basal Ganglia Volume with Fatigue and Set-Shifting.
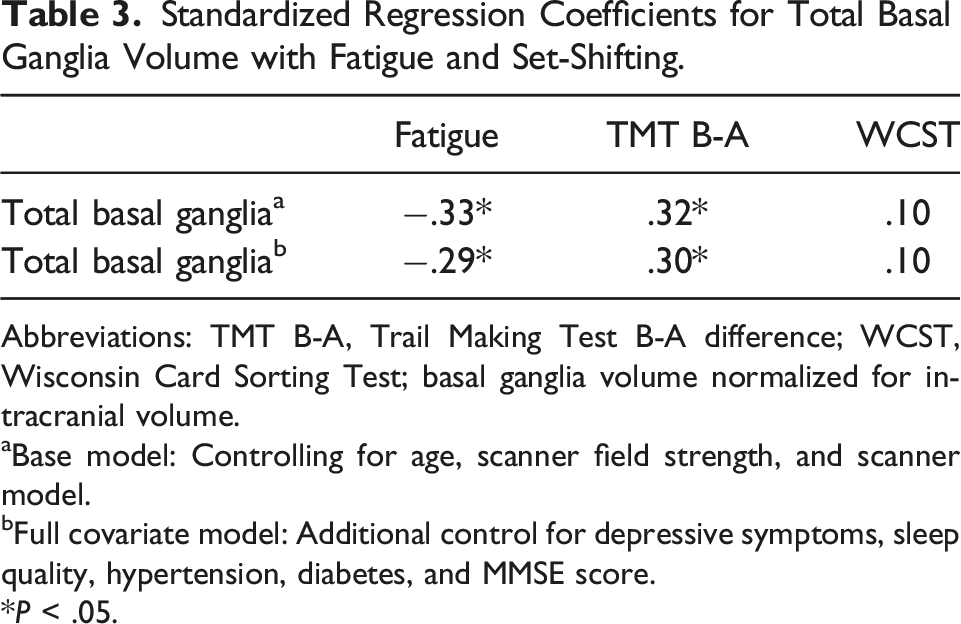
Abbreviations: TMT B-A, Trail Making Test B-A difference; WCST, Wisconsin Card Sorting Test; basal ganglia volume normalized for intracranial volume.
aBase model: Controlling for age, scanner field strength, and scanner model.
bFull covariate model: Additional control for depressive symptoms, sleep quality, hypertension, diabetes, and MMSE score.
*P < .05.
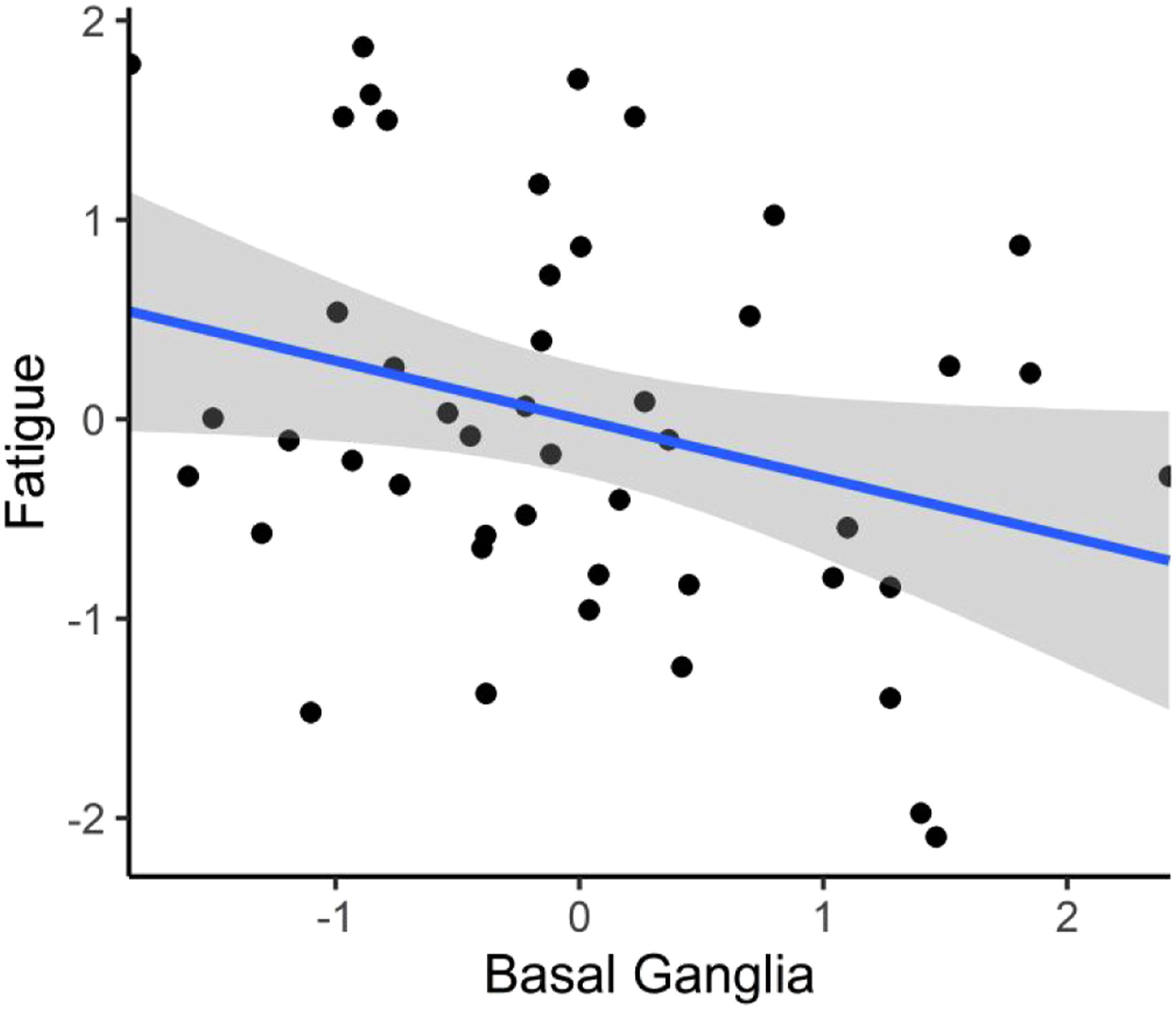
The standardized association between total basal ganglia volume and fatigue in the full covariate model.
Total Basal Ganglia Volume and Set-Shifting
Table 3 also shows the standardized regression coefficients for set-shifting measures regressed on total basal ganglia volume. Smaller total basal ganglia volume was significantly associated with poorer TMT B-A performance both in the base model (β = .32, t (43) = 2.16, P = .036, R2 = .11, local f2 = .12) and full covariate model (β = .30, t (38) = 2.21, P = .033, R2 = .38, local f2 = .11). Figure 2 depicts the standardized association between smaller total basal ganglia volume and poorer TMT B-A controlling for covariates. In the full covariate model, the local f2 value fell in the small-to-medium effect size range, and the semipartial correlation indicated that bilateral basal ganglia volume accounted for 7.6% of unique variance in TMT B-A performance. Total basal ganglia volume was not associated with WCST scores in the base model (β = .10, t (43) = .66, P = .515, R2 = .15) or full covariate model (β = .10, t (38) = .76, P = .455, R2 = .45). The standardized association between total basal ganglia volume and TMT B-A in the full covariate model. Shaded regions represent the 95% confidence intervals.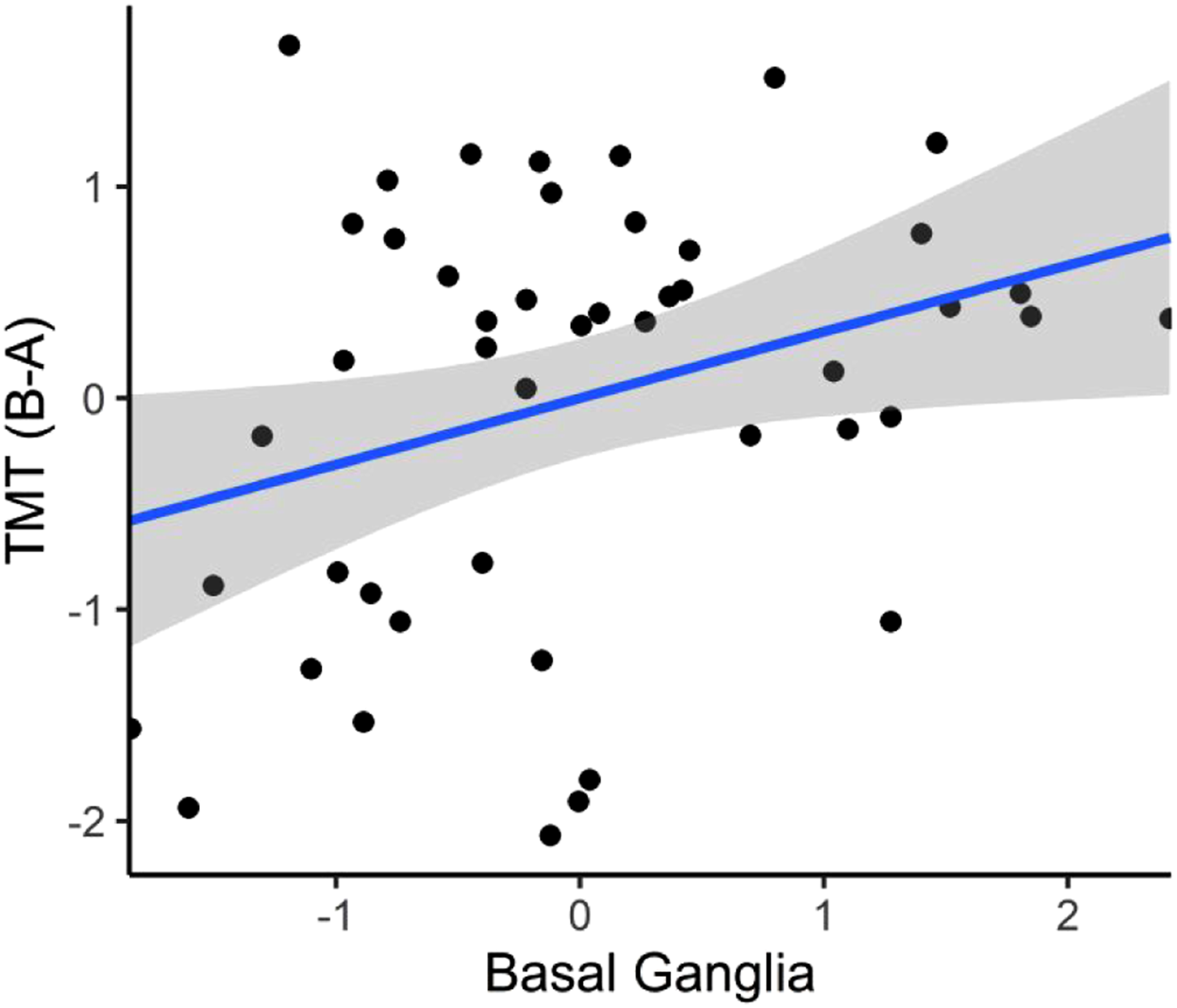
Sensitivity analysis. Post-hoc analyses confirmed that these findings were robust to differences in field strength and model among scanners. Results are provided in the Supplementary Material.
Individual Basal Ganglia Structures, Fatigue, and Set-Shifting
Partial Correlations for Basal Ganglia Structures with Fatigue and Set-Shifting.
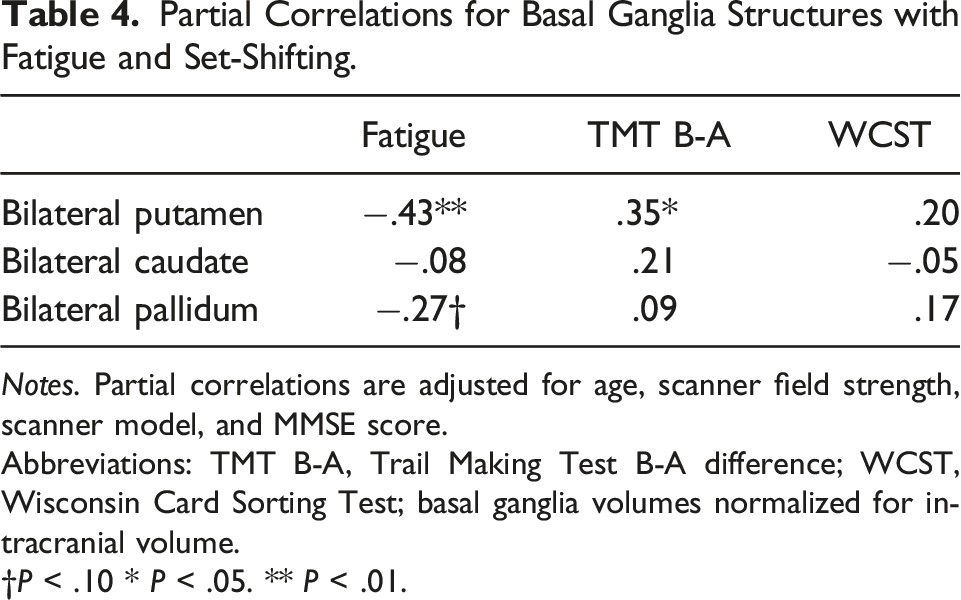
Notes. Partial correlations are adjusted for age, scanner field strength, scanner model, and MMSE score.
Abbreviations: TMT B-A, Trail Making Test B-A difference; WCST, Wisconsin Card Sorting Test; basal ganglia volumes normalized for intracranial volume.
†P < .10 * P < .05. ** P < .01.
Clinically Elevated and Minimal Fatigue Group Comparisons
Group comparisons are shown in Table 2. Approximately 56% of the sample reported scores above the cutoff for clinically elevated fatigue severity. Compared to the minimal fatigue group, subjects in the clinically elevated fatigue group reported significantly higher levels of depressive symptoms, t (46) = −4.44, P < .001, and performed worse on the WCST, t (46) = 2.41, P = .020). They did not significantly differ in age, gender, ethnicity, language of testing, education, hypertension, diabetes, sleep quality, or normalized basal ganglia volumes (all P > .05).
Partial Correlations for Total Basal Ganglia Volume with Set-Shifting in the Clinically Elevated and Minimal Fatigue Subgroups.

Notes. Partial correlations are adjusted for age, scanner field strength, scanner model, and MMSE score.
Abbreviations: TMT B-A, Trail Making Test B-A difference; WCST, Wisconsin Card Sorting Test; basal ganglia volumes normalized for intracranial volume.
*P < .05.
Discussion
In the present study, we found that smaller bilateral basal ganglia volumes were associated with higher levels of subjective fatigue controlling for age, depressive symptoms, poor sleep quality, hypertension, diabetes, and global cognitive status in a sample of middle-aged and older adults without dementia clinically evaluated for cognitive complaints. Among the individual basal ganglia structures examined, subjective fatigue was most strongly associated with reduced putamen volumes. Moreover, we found a similar link between smaller total basal ganglia volume and weaker timed set-shifting ability, an effect that was pronounced in subjects with clinically elevated levels of fatigue compared to their less fatigued counterparts.
This study is among the first to identify reduced basal ganglia volume as a neurostructural correlate of fatigue in physically able middle-aged and older adults without diagnosed neurological conditions or frank dementia. Consistent with the basal ganglia model of fatigue, 15 this finding suggests that degeneration within the basal nuclei, and putamen in particular, may contribute to fatigue by disrupting dopamine-dependent striato-cortical neural systems involved in energy monitoring and regulation through modulation of effort perception and motivation.11,15,16,34,35
Our findings add to a growing body of structural neuroimaging findings linking reduced basal ganglia grey-matter volumes with higher levels of fatigue in clinical populations15-17 including Parkinson’s disease 19 and multiple sclerosis. 36 We advance this literature by demonstrating this link in middle-aged and older adults free of known neurological illnesses, suggesting that neuropathogenic mechanisms outlined in the basal ganglia model of fatigue 15 are not disease specific and may overlap across diverse populations.
Compared to the few published studies examining neurostructural correlates of fatigue measured by validated self-report in adults without diagnosed neurological conditions, 18 our findings agree with a recent study 20 showing that reduced gray-matter volumes in the putamen, hippocampus, and thalamus were associated with higher levels of physical fatigability among adults with poor physical functioning in their seventh and eighth decades of life. Similarly, a study in healthy young adults 37 found that higher levels of self-reported fatigue were associated with lower neural density in the right putamen, pallidum, and caudate, suggesting a basal ganglia-fatigue link may span across the aging continuum. In contrast to our findings, a study involving 41 healthy older adult controls 19 with higher mean MMSE scores, lower BDI-II scores, and minimal fatigue symptoms compared with our sample did not find an association between basal ganglia volumes and self-reported fatigue. This discrepancy may be attributed to differences in our sample characteristics including our use of a clinic sample with cognitive complaints that was not deliberately selected to be in optimal health.
In our data, reduced total basal ganglia volume was also associated with poorer rapid set-shifting measured by the TMT B-A, complementing previous reports linking attenuated basal ganglia grey-matter volumes with speeded executive functioning deficits.38,39 Given that rapid set-shifting is dependent on striatofrontal integrity, 21 it is tenable to propose that this association reflects striatofrontal dysfunction secondary to basal ganglia degeneration. The fact that this finding was pronounced in subjects with clinically elevated fatigue but absent in their minimally fatigued counterparts suggests that striatofrontal dysfunction may be an important feature of clinical fatigue among middle-aged and older adults.
Unlike the TMT B-A, we did not find an association between basal ganglia volume and performance on the WCST in either group. It is possible that the TMT B-A score provided a more targeted behavioral index of striatal dysfunction. This timed measure has previously been linked with striatal atrophy and hypoperfusion in older adults,23,38 and the B-A correction is thought to provide a more refined representation of rapid set-shifting by adjusting for working memory and information processing demands. 31 In contrast, the WCST is an untimed measure involving a wider range of executive functions weighted heavily on reasoning, attentional control, and working memory, 30 and elicits predominant frontal activations. 40 Alternatively, as the TMT is a largely motor-based task, it is plausible that poorer scores in part reflect dysfunction of striatal motor pathways. However, our use of the TMT B-A correction likely reduced potential confounding effects of motor impediments.
The present study identified the putamen as a key basal ganglia structure in relation to both fatigue and rapid set-shifting. These parallel associations are consistent with the putamen’s roles in modulating striato-cortical circuits underpinning motivational, affective, and cognitive functions. 35 Interestingly, the putamen has been implicated in fatigue pathogenesis for decades including early findings of putaminal hypometabolism in Parkinson’s disease 41 and more recent models positing the putamen as locus of neural networks governing fatigue in older adults. 42 Additionally, prior neuroimaging studies in older adults have linked putaminal atrophy with poorer executive functioning 43 and associated reduced resting state putaminal-cortical functional connectivity with age-related declines in executive control. 44 It is somewhat surprising we did not find caudate volume to be associated with fatigue or either set-shifting measure. In the current literature, the caudate is understood to play a role in learning, memory, initiation, and maintenance of adaptive goal-directed behavior, whereas putamen is commonly implicated in less complex cognitive processes including habit learning, stimulus response associations, and sensorimotor coordination. 45 As much of this work comes from animal studies and disease models of basal ganglia dysfunction, more work is needed to clarify the differential non-motor functions of the basal nuclei in older adults without neurological disease.
Altogether, our findings suggest that basal ganglia atrophy, particularly involving the putamen, may be a common neural substrate underlying co-occurring experiences of elevated fatigue and reduced rapid set-shifting in middle-aged and older adults. This may help shed light on the documented relationship between elevated fatigue and cognitive deficits in older adults8,9 as well as recent findings linking decreased striato-cortical functional connectivity with heightened subjective fatigue ratings during demanding executive function tasks. 42 There are numerous factors that may explain diminished basal ganglia and putamen volumes among older adults including developmental neuroanatomical differences, subcortical grey-matter atrophy associated with “normal” aging, 46 vulnerability to neuroinflammation, 47 cerebral small-vessel disease and silent lacunar infarcts, 48 and Alzheimer’s disease pathology. 49 Further studies are needed to elucidate disease mechanisms of basal ganglia degeneration and their potential downstream consequences for vitality and cognition.
Limitations of this study include the generalizability of our findings to older adults without subjective cognitive complaints and in some cases, mild cognitive deficits. It is noteworthy, however, that our sample comprised non-demented community-dwelling individuals and that the findings were maintained controlling for global cognitive status. Another limitation is that we cannot attribute our findings to aging specifically, as we controlled for age in the analyses and did not include a younger comparison group. While we identified these findings in an older cohort, the study design does not suggest they are unique to this age group. Due to limited sample size and data availability, we did not statistically control for medical conditions beyond hypertension and diabetes or medication use. However, we attempted to stringently exclude confounding neurological and medical conditions by chart review. In addition, the present study was cross-sectional and thereby precludes drawing definitive conclusions regarding directionality or causality. For instance, we acknowledge that fatigue may precede basal ganglia atrophy through reduced engagement in salutary behaviors (e.g., exercise) associated with healthy brain aging. Furthermore, fatigue was assessed by self-report, and therefore the findings may not generalize to more objective measures of fatigue or fatigability. Finally, MRI data came from clinical scans across multiple scanners rather than a standardized protocol; however, volumetric data were derived almost exclusively from T1-weighted MPRAGE sequences, scanner strength and model were statistically controlled in all analyses, and sensitivity analyses suggested that using multiple scanners did not bias the results.
Clinical Implications and Conclusions
We believe the present study offers an important first step toward understanding neuropathological mechanisms of subjective fatigue and co-occurring cognitive deficits in middle-aged and older adults without dementia or neurologic conditions. Our findings suggest that the basal ganglia model of fatigue 15 may extend to this population, thus providing a framework and potential neural targets for studying the treatment of idiopathic fatigue. While treatments for fatigue in the older adult population are scarce, interventions targeting the basal ganglia may hold clinical promise. For example, a 12-month coordination training intervention was associated with augmented basal nuclei volumes in older adults. 39 Additionally, some psychostimulant medications have been shown to ameliorate fatigue in clinical populations ostensibly by correcting dopamine imbalances within striato-frontal circuits. 34 Whether similar interventions may benefit fatigued older adults without neurological illnesses is an intriguing question for future research. Finally, our findings suggest that high levels of subjective fatigue may warrant enhanced clinical attention (e.g., cognitive and neuroimaging assessments), and raise the question of whether idiopathic fatigue, particularly in the context of cognitive complaints, may be an early preclinical marker of pathological brain aging. An important next step will be to prospectively determine whether heightened fatigue in middle age predicts development of dementia in later life, and whether neurodegeneration of the basal ganglia may contribute to this link.
Supplemental Material
sj-pdf-1-jgp-10.1177_08919887211070264 – Supplemental Material for Structural Basal Ganglia Correlates of Subjective Fatigue in Middle-Aged and Older Adults
Supplemental Material, sj-pdf-1-jgp-10.1177_08919887211070264 for Structural Basal Ganglia Correlates of Subjective Fatigue in Middle-Aged and Older Adults by Nikhil Banerjee, Sonya Kaur, Anita Saporta, Sang H. Lee, Noam Alperin and Bonnie E. Levin in The American Surgeon
Footnotes
Acknowledgments
We thank the McKnight Brain Institute and all of the study participants who generously gave their time in support of this research.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was completed at the University of Miami Miller School of Medicine in Miami, FL.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.