
Abstract
Objective
The continuous contact of glucose-containing peritoneal dialysis (PD) fluids with the peritoneum results in the intraperitoneal formation of early and advanced glycation end-products. This nonenzymatic glycation of proteins may cause morphological and functional alterations to the peritoneum, which may contribute to patient dropout from PD therapy. Because fibrinolytic system components have been demonstrated to play an important role in the balance of intraperitoneal generation and degradation of fibrin, we studied the effect of early and advanced glycated human serum albumin, methylglyoxal, and 3-deoxyglucosone on the synthesis of tissue-type plasminogen activator (tPA), as well as its specific inhibitor (PAI-1), in human peritoneal mesothelial cells (HPMC).
Methods
Antigen concentrations in the supernatants of cultured HPMC were measured by ELISA. Northern blot analysis was conducted for mRNA expression. Electrophoretic mobility shift assays were applied to demonstrate the involvement of the transcription factors nuclear factor kappa B (NF-κB) and activator protein-1 (AP-1) in signal transduction.
Results
Incubation of HPMC with early glycated albumin (GHSA) resulted in a time- and concentration-dependent increase in PAI-1 mRNA expression and antigen secretion. In contrast, no changes in PAI-1 synthesis occurred after stimulation with either the 1,2-dicarbonyl compounds methylglyoxal and 3-deoxyglucosone, or with late advanced glycation end-products. tPA synthesis was not affected by any of the tested components. Furthermore, HPMC exposed to GHSA induced NF-κB and AP-1 DNA binding activity, suggesting that GHSA-induced over-expression of PAI-1 is transcriptionally regulated by both transcription factors.
Conclusions
We conclude that Amadori modified glycated albumin upregulates PAI-1 synthesis in HPMC, possibly mediated through the activation of the transcription factors NF-κB and AP-1. The present data support the clinical relevance of the formation of glycated proteins and their involvement in pathological processes in PD patients. Thus, glycated albumin may contribute to an imbalance between intraperitoneal formation and degradation of fibrin that causes peritoneal structural alterations, with subsequent membrane failure.
Keywords
Previous studies have focused predominantly on intraperitoneal formation of AGEs and their pathophysiological impact on peritoneal structure and function (5,6). In contrast, little is known about the role of Amadori adducts of glycated albumin, the most abundant protein in peritoneal dialysate.
As we have shown in our previous work, human peritoneal mesothelial cells (HPMC) play an important role in maintaining an adequately functioning fibrinolytic system in the peritoneal cavity by secreting and degrading the fibrinolytic enzyme tissue-type plasminogen activator (tPA), as well as its specific plasminogen activator inhibitor-1 (PAI-1) (7,8). This plasma-independent fibrinolytic system is crucial in the dissolution of fibrinous exudates caused by a chronic inflammatory state of the peritoneum during PD (9,10). Depending on the mesothelial fibrinolytic capacity, fibrin deposits are lysed or become organized with a proliferation of fibroblasts, and may promote the formation of peritoneal lesions (11,12).
The goal of the present experiments was to evaluate whether early glycated albumin or advanced glycated albumin modulates the synthesis of fibrinolytic system components in HPMC.
In addition to our previous findings, where we showed that high concentrations of d-glucose increase mesothelial tPA production, we report here that exposure of HPMC to glycated human serum albumin (GHSA), but not to advanced glycated albumin, stimulates the expression of PAI-1 in HPMC. By investigating possible participating signal transduction pathways, we identified the involvement of the sequence-specific DNA binding of the transcription factors nuclear factor kappa B (NF-kB) and activator protein-1 (AP-1).
Materials and Methods
Materials
Medium M199 and newborn calf serum were obtained from Gibco BRL (Eggenstein, Germany), tissue culture plates were from Costar (Cambridge, Massachusetts, U.S.A.). Human serum was prepared from freshly collected blood of healthy donors and stored at –20°C. Fibronectin from human serum and trypsin were purchased from Boehringer (Mannheim, Germany). Collagenase type II was from Worthington Biochemical (Freehold, New York, U.S.A.). Monoclonal antibodies against cytokeratins 8 and 18 and against vimentin were a gift from Dr. G. van Muijen (University of Nijmegen, The Netherlands). Hybond-N filters, Hyperfilm-MP, and [a-32P]dCTP (3 mCi/mmol) were obtained from Amersham (Buckinghamshire, United Kingdom). Human albumin and GHSA were obtained from Sigma (Deisenhofen, Germany). The GHSA contained 1 – 5 moles fructosamine per mole albumin and was produced as described by Baynes et al. (13). The GHSA preparation did not contain measurable AGEs as demonstrated by fluorescence assays (360 – 600 nm). Contamination of the glycated human albumin preparation with endotoxin was excluded using a Limulus amebocyte lysate assay (LAL endotoxin kit; Endosafe Inc., Charleston, South Carolina, U.S.A.).
Advanced glycation end-product–human serum albumin [AGE-HSA (glu)] and AGE-HSA (G-6-P) were prepared by incubation of HSA (50 mg/mL) with glucose (0.5 mol/L) or glucose-6-phosphate in 0.1 mol/L phosphate buffer (pH 7.4) at 37°C in the dark. Aliquots were removed after 9 weeks, dialyzed against phosphate-buffered saline (PBS), and stored frozen at –20°C until analyzed.
Methylglyoxal was freshly prepared by the acid hydrolysis of 1,1-dimethoxyacetone (Sigma) and purified by distillation as described (14). 3-Deoxyglucosone was prepared as described (15) and purified with a Kieselgel 60 column (Merck, Darmstadt, Germany) with pentane/ethyl acetate/methanol (2:1:1) as the solvent system. Decolorization was carried out with activated carbon. The preparation was dried over P2O5. Thin layer chromatography (silica gel 60) and 1H-NMR (nuclear magnetic resonance) analysis indicated a pure substance. More details concerning analysis of 1,2-dicarbonyl compounds are provided by Schalkwijk et al. (4).
In the experiments, 15 mmol/L HSA, 15 mmol/L AGE-HSA (glu), 15 mmol/L AGE-HSA (G-6-P), 330 mmol/L methylglyoxal, and 1 mmol/L 3-deoxyglucosone were used, amounts that were derived from data found in peritoneal dialysates from patients on continuous ambulatory PD (4).
Tissue Culture
Human peritoneal mesothelial cells were isolated, as described previously (16), from the omental tissue of consenting patients undergoing elective surgery, and the experimental protocol was approved by the local Ethics Committee.
Cells were grown in fibronectin-coated dishes in M199 medium supplemented with 25 mmol/L HEPES (pH 7.3), 2 mmol/L glutamine, 10% (v/v) human serum (HS), 10% (v/v) newborn calf serum (heat-inactivated), penicillin (100 U/mL), and streptomycin (100 m g/mL) at 37°C under 5% CO2/95% air atmosphere. The medium was replaced every 2 – 3 days. Subcultures were obtained by trypsin/EDTA treatment at a split ratio of 1:3. Cells from omental tissue were pure mesothelial cells as assessed by their uniform cobblestone appearance at confluence, by the absence of von Willebrand factor, and by their uniform positive staining for cytokeratins 8 and 18 and vimentin (16). For the experiments, confluent cultures were used at the second or third passage, and cells were always refed 48 hours before the experiment with M199 supplemented with 0.5% HS and antibiotics. Incubation of cells with doses of tested compounds for up to 24 hours did not have any significant effect on cell viability as tested by vital cell staining with acridine orange and ethidium bromide and by LDH release (cell viability exceeded 95%). Conditioned media were obtained by incubating cells in 2-cm2 dishes at 37°C with 0.5 mL serum-free medium containing the appropriate concentration of the test compound. Conditioned media were centrifuged 5 minutes (2000g) to remove cells and cellular debris, and samples were frozen at –20°C until use. All experiments were done with cells from 5 to 8 individual donors and were measured in quadruplicate.
Assays
Assay of tPA antigen was performed by Coaliza tPA from Chromogenix (Mölndal, Sweden). In this assay, free tPA and tPA/PAI-1 complexes are detected with equal efficiency. PAI-1 antigen was measured by ELISA (Coaliza PAI-1), also obtained from Chromogenix. Diluted aliquots of the cell supernatants were assayed without prior purification.
Rna Isolation and Northern Blotting
Total RNA was extracted from cells, as described by Chomczynski and Sacchi (17). RNA samples were dissolved in H2O. The RNA concentration in each sample was determined spectrophotometrically. Equal amounts of RNA from different dishes were analyzed by Northern blot hybridization for their PAI-1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA content. With Northern blotting, RNA samples were subjected to gel electrophoresis in formaldehyde–agarose gels. After electrophoresis, RNA was transferred to Hybond-N according to the manufacturer's instructions. Hybridization was usually performed at 63°C with 1 ng/mL probe, labeled by random prime labeling with [a-32P]dCTP (Mega-prime; Amersham Pharmacia Biotech, Freiburg, Germany) to approximately 2 x 108 cpm/mg DNA. The membranes were subsequently exposed to Hyperfilm-MP with an intensifying screen at -80°C.
cDNA PROBES
The following cDNA fragments were used as probes in the hybridization experiments: a 2.5-kb EcoRI fragment of human PAI-1 cDNA of the 3.1-kb transcript (18), and a 1.2-kb PstI fragment of a rat GAPDH cDNA provided by Dr. R. Offringa (University of Leiden, The Netherlands), which is commonly used as an internal standard probe (19).
Electrophoretic Mobility Shift Assays (EMSA) for the Transcription Factors Nf-Kb and Ap-1
HPMC were grown to confluence on 60-cm2 plates and were then stimulated with GHSA, tumor necrosis factor alpha (TNFa), HSA, or control medium for 1 hour. After that, cells (2 – 3 x 106) were washed with PBS medium and scrubbed off the plates with a cell scraper. Nuclear extracts were prepared according to the method of Ziegler–Heitbrock et al. (20). Briefly, cells were centrifuged and resuspended in 150 mL buffer A (10 mmol/L HEPES, pH 7.9; 1.5 mmol/L MgCl2; 10 mmol/L KCl; and 0.5% aprotinin) and disrupted by ultrasound. After centrifugation, the nuclear pellets were resuspended in 40 mL ice-cold buffer B (20 mmol/L HEPES, pH 7.9; 25% glycerol; 1.5 mmol/L MgCl2; 0.42 mol/L NaCl; 0.2 mmol/L EDTA; and 2 mmol/L DTT) and incubated for 1 hour. The extracts were then centrifuged for 5 minutes at 20 000 rpm and the supernatants were mixed with 40 mL buffer C (20 mmol/L HEPES, pH 7.9; 20% glycerol; 100 mmol/L KCl; 0.05 mmol/L EDTA; 0.5 mmol/L PMSF; and 1% NP-40) and frozen at -70°C until used. The protein contents of the extracts were determined by a DC protein assay kit from BioRad (Munich, Germany), and nuclear protein (10 mg) was mixed with double-stranded oligonucleotide containing the specific DNA binding domain for NF-kB (cs 5’-GGG GAC TTT CCC-3’) or AP-1 (cs 5’-CGC TTG ATG AGT CAG CCG GAA-3’). Oligonucleotides were end labeled with [g-32P]ATP using T4 polynucleotide kinase. Extracts were then incubated for 15 minutes at room temperature with labeled NF-kB or AP-1 oligonucleotides. DNA–protein complexes were separated on a nondenaturing 4% acrylamide gel. The gel was then dried and exposed to Kodak XAR-5 film.
Statistical Analysis
Data are given as mean ± SEM. Statistical analysis was performed using the Wilcoxon matched-pairs signed rank test for nonparametric data. A p value of less than 0.05 was considered indicative of statistically significant differences.
Results
Confluent HPMC were incubated in medium M199 supplemented with various concentrations of glycated albumin (1.5 – 30 m mol/L) over 48 hours. As shown in Figure 1(a), the addition of GHSA resulted in a dose-dependent increase in PAI-1 antigen levels in the cell supernatant, with statistical significance at a concentration of 7.5 m mol/L (p < 0.05). PAI-1 antigen was enhanced almost twofold by 30 mmol/L GHSA after a 48-hour incubation period compared to nonstimulated conditions (362.35 ± 17.96 ng/105 cells vs 160.99 ± 2.5 ng/105 cells, n = 7). PAI-1 synthesis was not affected by nonglycated HSA (152.75 ± 2.99 ng/105 cells). In contrast to PAI-1 antigen, the production of tPA was not affected by any of the concentrations of glycated albumin used (data not shown). In contrast to GHSA, neither the intermediates in the formation of AGEs, methylglyoxal and 3-deoxyglucosone, nor advanced glycated albumin [AGE-HSA (glu), AGE-HSA (G-6-P)] affected PAI-1 production [Figure 1(b)].
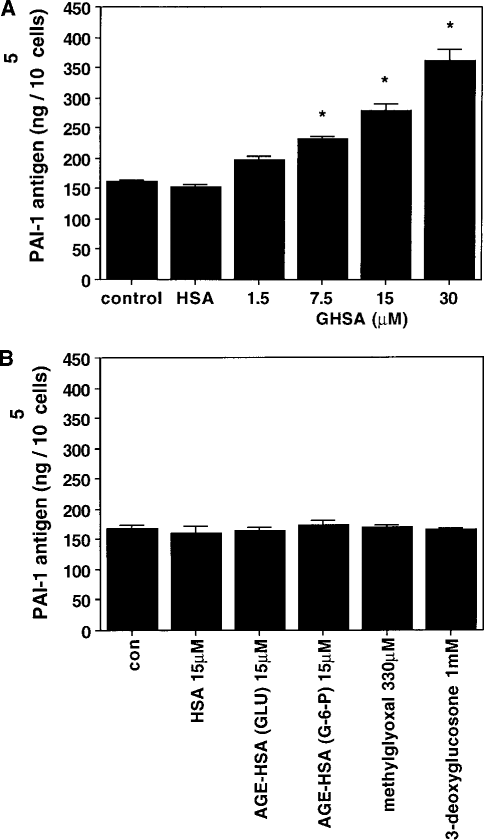
Effect of increasing concentrations of glycated human serum albumin (GHSA) on plasminogen activator inhibitor (PAI-1) synthesis in human peritoneal mesothelial cells (HPMC) (A). PAI-1 antigen levels were assayed in conditioned medium obtained after a 48-hour incubation of HPMC in M199 supplemented with increasing amounts of GHSA (1.5 – 30 mmol/L) and 15 mmol/L human serum albumin (HSA). Values are mean ± SEM of 7 independent experiments.
In order to assess the time course of GHSA on PAI-1 synthesis in HPMC, confluent cells were incubated with 30 mmol/L GHSA for various time periods. As shown in Figure 2, PAI-1 protein levels increased continuously, reaching statistically significant differences after 24 hours.
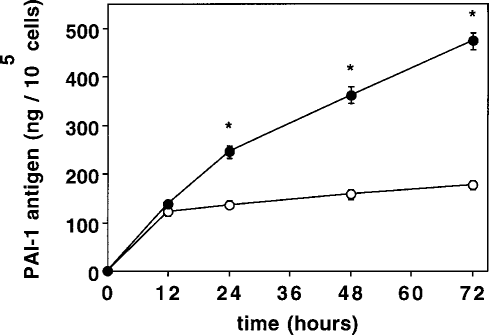
Time course of plasminogen activator inhibitor (PAI-1) synthesis in human peritoneal mesothelial cells exposed to 30 mmol/L glycated human serum albumin (GHSA) (closed circles), or control medium without GHSA (open circles). PAI-1 antigen levels were assayed in conditioned medium obtained after different incubation times (0 – 72 hours). Values are mean ± SEM of 5 independent experiments.
Figure 3 shows that the mRNA expression of PAI-1 was significantly upregulated by 1.5 – 30 mmol/L human glycated albumin. The increase in PAI-1 antigen levels [Figure 1(a)] was very well reflected in PAI-1 mRNA expression (Figure 3). No effect has been observed with nonglycated HSA on PAI-1 mRNA expression. In contrast to PAI-1, tPA antigen levels and mRNA expression were not affected by GHSA (data not shown).
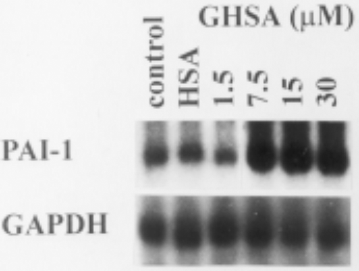
Effect of increasing concentrations of glycated human serum albumin (GHSA) on plasminogen activator inhibitor (PAI-1) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA expression in human peritoneal mesothelial cells (HPMC). HPMC were incubated for 12 hours in M199 supplemented with GHSA (1.5 – 30 mmol/L) and 15 mmol/L human serum albumin (HSA). Total RNA was extracted from cells and analyzed by Northern blot hybridization.
Activation of Nf-Kb and Ap-1 by Ghsa in HPMC
It has been determined that the expression of PAI-1 is initiated by the transcription factors NF-kB and AP-1. Therefore, we investigated whether these transcriptional regulators might also be involved in GHSA-dependent gene expression. As demonstrated by EMSA, GHSA induced both NF-kB and AP-1 DNA binding activity (Figure 4). These results indicate that both transcription factors may be important for the upregulation of PAI-1 expression in HPMC.
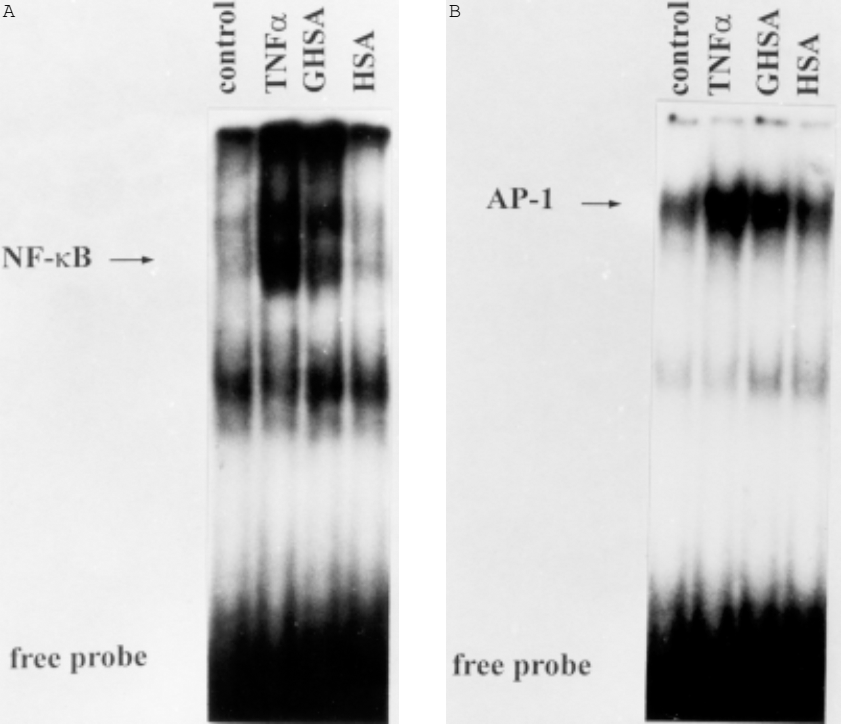
Gel shift assay for nuclear factor kappa B (NF-kB) (A) and activator protein-1 (AP-1) (B) in nuclear extracts from human peritoneal mesothelial cells (HPMC). HPMC were incubated for 1 hour in medium M199 alone or in medium supplemented with tumor necrosis factor alpha (TNFa) (500 U/mL), glycated human serum albumin (GHSA) (30 mmol/L), or human serum albumin (HSA) (30 mmol/L). Nuclear extracts were prepared and analyzed for activation of NF-kB and AP-1 by electrophoretic mobility shift assays. One of three representative experiments is shown.
Discussion
In patients treated by PD, continuous exposure of the peritoneal cavity to dialysis solutions containing high glucose (75 – 216 mmol/L) results in glycation of intraperitoneal proteins as well as accumulation of AGEs (2). Early and advanced glycation end-products may cause morphological and functional alterations of the peritoneum (6,21), which, in sequence, may lead to patient dropout from PD therapy.
Hyperglycemia is the driving force for increased albumin glycation, and glucose concentrations correlate with the glycation rate of proteins in PD fluids (22). A major target for glycation is serum albumin, the most abundant protein in the peritoneal cavity (23). Thus, for our experiments we used the early glycation products of albumin in Amadori configuration, which have been detected in dialysates of PD patients (1) and are known to be involved in microvascular complications (24). The AGEs of albumin have been reported to affect important cellular functions in different cell types, including the synthesis of extracellular matrix proteins, inflammatory mediators, and nitric oxide (25-27).
HPMC are the major source of fibrinolytic components in the peritoneal cavity (16). They produce tPA as well as its specific inhibitor, PAI-1. An imbalance between the mesothelial production of tPA and PAI-1 may account for the peritoneal fibrotic thickening that has been found after long-term PD (11,28). In the present study, exposure of HPMC to GHSA resulted in a concentration- and time-dependent increase in PAI-1 mRNA expression and antigen secretion without affecting tPA synthesis. A similar effect of glycated low density lipoprotein has been shown on PAI-1 synthesis in vascular endothelial cells (29).
Our findings may contribute to a plasma-independent hypercoagulatory state with permanent intraperitoneal fibrin formation, which has been described in PD patients (12,30). In contrast, the 1,2-dicarbonyl compounds, methylglyoxal and 3-deoxyglucosone, which are present in PD fluids and have been identified as intermediates in the formation of AGEs (4), were not able to modulate PAI-1 synthesis. Also, AGE-modified albumin, which was found to stimulate PAI-1 expression in human microvascular endothelial cells (31), did not affect PAI-1 synthesis in our experiments. This difference may be explained by cell-type specific regulation of tPA and PAI-1 synthesis in endothelial and mesothelial cells, as shown in our previous work (7).
The effect of GHSA is independent of hyperglycemia, which per se stimulates tPA synthesis in HPMC without increasing PAI-1 expression (32). Although the mechanism by which GHSA stimulates PAI-1 synthesis is still speculative, it has been suggested that a cell-associated ligand-receptor system for the Amadori modified protein is involved in mediating the effects of glycated albumin on cell function (33-35). This possibility is supported by the finding of specific cell surface receptors for early glycosylated products of albumin in rat peritoneal cells (36).
It is known that transcription factors such as NF-kB and AP-1 are involved in the regulation of PAI-1 gene expression (37,38), and that glycated albumin activates NF-kB in vascular smooth muscle cells (39). Thus, we investigated whether the transcriptional regulators NF-kB and AP-1 might also play a role in GHSA-dependent PAI-1 gene expression. We demonstrated that HPMC exposed to GHSA induce both NF-kB and AP-1 DNA binding activity. These results suggest that GHSA-induced overexpression of PAI-1 could possibly be transcriptionally regulated by both transcription factors.
In summary, the findings reported herein are the first to demonstrate that the early glycated protein albumin in Amadori configuration upregulates the major physiological regulatory protein of the fibrinolytic system PAI-1 in HPMC. This stimulatory effect is possibly mediated through binding to a still undefined receptor, followed by a signal transduction pathway that involves the transcription factors NF-kB and AP-1. The present data further support the clinical relevance of the formation of glycated proteins and their involvement in pathological processes in PD patients. It is reasonable to speculate that glycated albumin may contribute to an imbalance between the intraperitoneal formation and degradation of fibrin that causes peritoneal structural alterations with subsequent membrane failure (11).
If glycated albumin is proved to be involved in pathological processes in PD patients, in particular an imbalance between intraperitoneal formation and degradation of fibrin, then glucose-polymer–based PD fluids may be an alternative to glucose-based PD fluids, since it has recently been demonstrated that the formation of glycated albumin is significantly lower with the glucose polymer icodextrin than with glucose (40). Alternatively, amino acid- and glycerol-based PD fluids might be considered.
Footnotes
Acknowledgment
This work was supported by a grant from Else Kröner-Fresenius-Stiftung to Thomas Sitter.