
Abstract
← Objective
← Methods
← Results
← Conclusion
High molecular weight hyaluronan (HA) is a nonsulfated glycosaminoglycan polymer that is a ubiquitously distributed component in extracellular matrix (1). Recent studies have shown that peritoneal mesothelial cells (MC) synthesize large amounts of HA and that the glycosaminoglycan may contribute to the remodeling and regeneration of the peritoneal membrane (2,3).
Hyaluronan is a major component in pericellular coats and, despite its simple structure, it is involved in several cellular biological functions (4). Concerning peritoneal dialysis (PD), it has been demonstrated that HA modifies the inflammatory response and peritoneal permeability during peritonitis (5), preserves peritoneal transport characteristics (6), and reduces peritoneal fluid absorption (7). According to these advantages and the idea that HA might be washed out of the peritoneal membrane by the PD process (6), the addition of exogenous HA to dialysis solutions has been suggested in order to stabilize the long-term function of the peritoneum as a dialysis membrane (8).
In the context of tissue repair and wound healing, HA has been reported to probably also have procoagulant and antifibrinolytic properties (9,10). As we have shown in our previous work, MC have an important role in maintaining an adequately functioning fibrinolytic system in the peritoneal cavity by secreting and degrading the fibrinolytic enzyme tissue-type plasminogen activator (t-PA), as well as its specific plasminogen activator inhibitor type 1 (PAI-1) (11). This plasma independent fibrinolytic system is crucial in the dissolution of fibrinous exudates caused by a chronic inflammatory state of the peritoneum during PD (12). Depending on the mesothelial fibrinolytic capacity, fibrin deposits are lysed or become organized with proliferation of fibroblasts and may promote the formation of peritoneal lesions (13).
Whether HA directly affects the synthesis of fibrinolytic system components in MC is unknown. In the present study, we investigated the impact of HA on the synthesis of t-PA and PAI-1.
Materials and Methods
Materials
Medium M199 and newborn calf serum were obtained from Gibco BRL (Eggenstein, Germany); tissue-culture plates were from Costar (Cambridge, Massachusetts, USA). Human serum was prepared from freshly collected blood of healthy donors and stored at –20°C. Fibronectin from human serum and trypsin were purchased from Boehringer (Mannheim, Germany). Collagenase type II was from Worthington (Freehold, New York, USA).
Monoclonal antibodies against cytokeratins 8 and 18 and against vimentin were a gift from Dr. G. van Muijen (University of Nijmegen, The Netherlands). Hybond-N filters, Hyperfilm MP, and [α-32P] dCTP (3 μCi/μmol) were obtained from Amersham (Buckinghamshire, UK).
Hyaluronan (average molecular weight 2.55 ∞ 106 Daltons) was purchased from Genzyme Pharmaceuticals (Cambridge, Massachusetts, USA).
Cell Culture Experiments
Human peritoneal MC were isolated from the omental tissue of consenting patients undergoing elective surgery, as described previously (14). Cells were grown in fibronectin-coated dishes in M199 medium supplemented with 25 mmol/L HEPES (pH 7.3), 2 mmol/L glutamine, 10% (v/v) human serum, 10% (v/v) newborn calf serum (heat inactivated), penicillin (100 IU/mL), and streptomycin (100 μg/mL) at 37°C under 5% CO2/95% air atmosphere. The medium was replaced every 2 to 3 days. Subcultures were obtained by trypsin/EDTA treatment at a split ratio of 1:3. The cells were purely MC, as assessed by their uniform cobblestone appearance at confluence, by the absence of von Willebrand factor, and by their uniform positive staining for cytokeratins 8 and 18 and for vimentin. In addition, incubation of cells with doses of the tested compounds did not have any significant effect on cell viability (as tested by vital cell staining with acridine orange and ethidium bromide, where living cells show green and dead cells show red fluorescence) when analyzed by fluorescence microscopy (cell viability exceeded 95%). For the experiments, confluent cultures were used at the second or third passage, and cells were always refed 24 hours before the experiment with M199 supplemented with 2% human serum. Conditioned media were obtained by incubating cells in 2-cm2 dishes at 37°C with 0.5 mL serum-free M199 containing the appropriate concentration of the test compound or stock solvent. Conditioned media were centrifuged 5 minutes to remove cells and cellular debris, and samples were frozen at –20°C until use.
Assays
Assay of t-PA antigen was performed using the ELISA Coaliza t-PA from Chromogenix (Mölndal, Sweden). In this assay, free t-PA and t-PA/PAI-1 complexes are detected with equal efficiency. PAI-1 antigen was measured by ELISA (Coaliza PAI-1), also obtained from Chromogenix.
RNA Isolation and Northern Blotting
Total RNA was extracted from cells as described by Chomczynski and Sacchi (15). RNA samples were dissolved in H20. The RNA concentration in each sample was determined spectrophotometrically. Equal amounts of RNA from different dishes were analyzed for their t-PA, PAI-1, low-density lipoprotein receptor-related protein (LRP), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA content by Northern blot hybridization. With Northern blotting, RNA samples were subjected to gel electrophoresis in formaldehyde-agarose gels. After electrophoresis, RNA was transferred to Hybond-N, according to the manufacturer's instructions. Hybridization was usually performed at 63°C with 1 ng/mL of probe labeled by random prime labeling to approximately 2 ∞ 108 cpm/μg DNA. The membranes were subsequently exposed to Amersham Hyperfilm-MP with an intensifying screen at –80°C.
cDNA Probes
The following cDNA fragments were used as probes in the hybridization experiments: a 1.9-kb Bgl II fragment of human t-PA cDNA (16), a 2.5-kb EcoRI fragment of human PAI-1 cDNA of the 3.1-kb transcript (17), a 6-kb XhoI/FcoRI fragment of the human LRP cDNA (18), and a 1.2-kb PstI fragment of a rat GAPDH cDNA, which is commonly used as an internal standard probe (provided by Dr. R. Offringa of the University of Leiden, The Netherlands).
Statistical Analysis
Data are given as mean ± SEM. Statistical analysis was performed using the Wilcoxon matched-pairs signed rank test for nonparametric data; a p value < 0.05 was considered indicative of statistically significant differences.
Results
Confluent MC were incubated with various concentrations of high molecular weight HA (10 – 100 mg/dL) for 48 hours. During this period, the number and viability of MC did not change compared to controls. As shown in Figure 1, the addition of HA resulted in a concentration-dependent decrease in t-PA antigen levels in the cell supernatant after 48 hours, which was statistically significant at a concentration of 50 mg/dL (p < 0.008). In the presence of 50 mg/dL HA, t-PA antigen release was diminished to 44% of the control. Hyaluronan equally decreased t-PA synthesis in a time-dependent manner; 48 hours’ incubation was required to observe a significant drop (p < 0.008) (Figure 1).
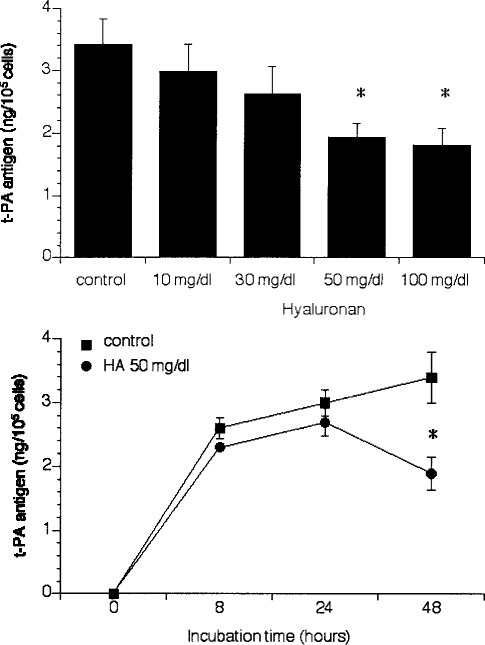
Concentration- and time-dependent effects of hyaluronan (HA) on tissue-type plasminogen activator (t-PA) production in mesothelial cells. Mesothelial cells were incubated for 48 hours with different concentrations of HA (10 – 100 mg/dL) and the conditioned media were analyzed for t-PA antigen (A). Mesothelial cells were incubated for various time periods up to 48 hours in the presence (closed circles) or absence (closed squares) of 50 mg/dL HA (B). Results are shown as mean ± SEM of six independent experiments done in duplicate.
In contrast to t-PA antigen, the synthesis of PAI-1 in MC was distinctly elevated 48 hours after the addition of HA, but the increase in PAI-1 expression was not significant (Figure 2).
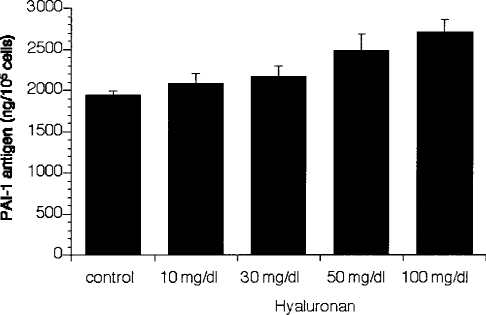
Concentration-dependent effect of hyaluronan (HA) on plasminogen activator inhibitor type 1 (PAI-1) production in mesothelial cells. Mesothelial cells were incubated for 48 hours with different concentrations of HA (10 – 100 mg/dL) and the conditioned media were analyzed for PAI-1 antigen. Results are shown as mean ± SEM of six independent experiments done in duplicate.
As shown in Figure 3, the decrease in t-PA secretion in the presence of HA (50 mg/dL) was preceded by comparable changes in mRNA expression after 8 and 12 hours, and this effect was also concentration dependent. These results indicate that the effect of HA manifests at the level of gene transcription or mRNA stability. Although the accumulation of PAI-1 antigen in conditioned media tended to increase, the mRNA levels of PAI-1 remained unchanged in the presence of HA (Figure 3). In addition, we investigated the mRNA expression of LRP, a multi-ligand receptor that is responsible for rapid degradation of t-PA in MC by receptor-mediated endocytosis (19). The mRNA expression of this receptor was not affected by HA. Therefore, it is reasonable to assume that increased t-PA degradation by receptor induction is not responsible for decreased t-PA levels after the addition of HA (Figure 3).
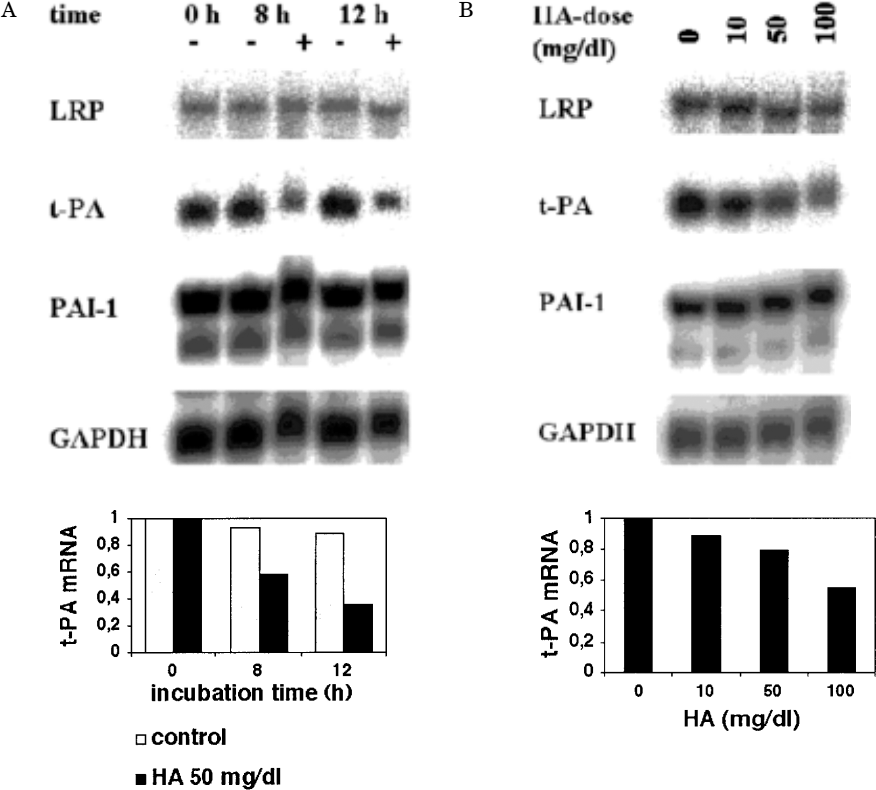
Low-density lipoprotein receptor-related protein (LRP), tissue-type plasminogen activator (t-PA), and plasminogen activator inhibitor type 1 (PAI-1) mRNA expression in mesothelial cells incubated with or without hyaluronan (HA). Mesothelial cells were incubated for 8 and 12 hours in the absence (–) or presence (+) of 50 mg/dL HA (A). Mesothelial cells were incubated for 12 hours with different concentrations of HA (10 – 100 mg/dL) (B). Total RNA was isolated and analyzed by Northern blot hybridization for t-PA, PAI-1, LRP, and GAPDH mRNA. The experiment shown is representative of three experiments. The signals for t-PA mRNA were quantified by densitometry and adjusted for corresponding GAPDH mRNA signals. The amount of t-PA is expressed relative to that found at time = 0 hours (A) or to that found at control condition (B).
To elucidate whether the cell surface antigen CD44, a specific receptor for HA, is involved in the signal transduction pathway, we pre-incubated cultured MC for 60 minutes with the monoclonal antibody BU75, which specifically blocks CD44. After pre-incubation, cells were treated either with medium as control or with HA 100 mg/dL for 48 hours. As shown in Figure 4, blocking of CD44 with BU75 could not inhibit the HA-induced decrease in t-PA production.
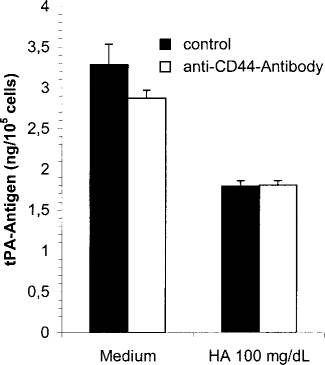
Effect of CD44 blocking of tissue-type plasminogen activator (t-PA) production in mesothelial cells incubated with hyaluronan (HA). Mesothelial cells were pre-incubated for 60 minutes with blocking anti-CD44 antibody Mab BU75 (white bars) or with medium (black bars). Then the cells were incubated with Mab BU75 (10 μg/mL) (white bars) containing medium or HA 100 mg/dL or with medium (black bars) in the presence or absence of HA 100 mg/dL. Incubation time was 48 hours. The conditioned media were analyzed for t-PA antigen. Results are shown as mean ± SEM of six independent experiments done in duplicate.
A recent publication (20) showed that HA increases the fibrinolytic response of MC exposed to tumor necrosis factor alpha (TNFα). We co-incubated MC with various concentrations of TNFα (10 – 1000 U/mL) and HA 100 mg/dL for 48 hours. As shown in Figure 5, HA was not able to block the TNFα-induced decrease in t-PA synthesis.
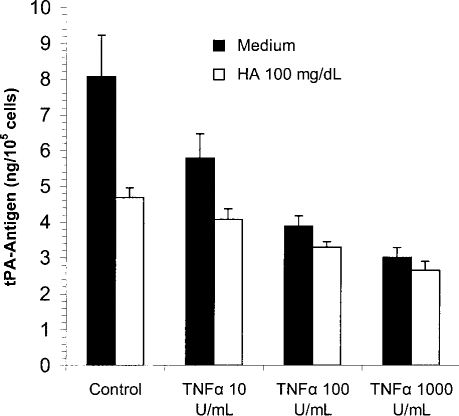
Effect of co-incubation with tumor necrosis factor alpha (TNFα) and hyaluronan (HA) on tissue-type plasminogen activator (t-PA) production in mesothelial cells. Mesothelial cells were incubated with TNFα at various concentrations, up to 1000 U/mL, in the presence (white bars) or absence (black bars) of HA 100 mg/dL. Incubation time was 48 hours. The conditioned media were analyzed for t-PA antigen. Results are shown as mean ± SEM of six independent experiments done in duplicate.
Discussion
Hyaluronan is a prominent component of extracellular matrix that has been shown to modify the inflammatory response and peritoneal permeability during peritonitis (5). Additionally, this glycosaminoglycan of high molecular weight preserves peritoneal transport characteristics and reduces peritoneal fluid absorption (6,7). Because HA is possibly washed out of the peritoneal membrane by the PD process (6), there have been suggestions to employ these properties in PD by adding HA, in concentrations up to 10 mg/dL, to the dialysis solution in order to preserve the peritoneum as a dialysis membrane.
Mesothelial cells that line the peritoneum as a superficial monolayer are not only a major source of intraperitoneally produced HA (21), but also of fibrinolytic system components in the peritoneal cavity. Depending on this mesothelial fibrinolytic capacity, fibrin deposits are lysed or become organized with proliferation of fibroblasts, and may promote the formation of peritoneal lesions (13). A culture model of these cells being the first contact layer with PD fluids and being crucial in the intraperitoneal fibrinolytic system seemed suitable to us in order to examine the effects of high HA concentrations — much higher than can be produced by the cells themselves. We focused on production of the fibrinolytic system component t-PA and its specific inhibitor PAI-1, taking into account that HA might also have procoagulant and antifibrinolytic properties (9,10). We found that concentrations of HA up to 30 mg/dL do not change t-PA and PAI-1 production significantly. A significant depression of t-PA levels (in protein as well as in mRNA), however, resulted — after incubation with HA concentrations of 50 mg/dL and higher — in a time-dependent order, whereas PAI-1 levels were slightly but not significantly elevated. According to these results, we cannot assume a generally depressive effect on gene transcription, or a generally reduced mRNA stability for HA in this fibrinolytic system. Moreover, non-elevated LRP mRNA levels in the presence of HA suggest that the decreased t-PA level is not a consequence of receptor-mediated endocytosis.
We presume that the effects of HA on gene transcription of fibrinolytic system components are the results of a nonspecific stimulatory effect and may be due partly to the viscosity of the HA solution, because the HA effect on t-PA and PAI-1 production as described above was not altered by inhibition of CD44 cell-surface antigen (with monoclonal antibody BU75), which is well known as a HA receptor and has a high density on MC (22). Accordingly, we were not able to inhibit this effect by treatment with PD98509, a specific inhibitor of the extracellular regulated kinase pathway (23) (data not shown). As we have shown previously, this pathway is involved in other HA effects (24).
There are studies reporting HA effects similar to our observations: Horton et al. noticed a PAI-1 enhancement and a urokinase-type plasminogen activator reduction under HA fragment treatment in mouse macrophages (10). Another study found a rise in PAI-1 due to increased HA levels in vascular smooth muscle cells of patients with type II diabetes (9). Similar to a recent publication (20), we co-incubated MC with TNFα and HA. According to the results of this group, we did not find major alterations of the TNFα-induced t-PA reduction due to HA in the culture supernatants. Against the background of our finding that HA, per se, reduces t-PA antigen in MC, we cannot confirm their thesis that HA increases the fibrinolytic response of MC exposed to TNF.
Hyaluronan also plays an enormous role in the noncellular fibrinogen clotting process: it binds specifically to fibrinogen (25), and this binding decreases the clotting time of fibrinogen solutions by enhancing the rate of fibrin polymerizations (26). Previous studies have demonstrated that, after the onset of tissue injury, a meshwork of fibrin rich in HA accumulates in the extracellular space (1). This provisional matrix becomes organized by invading cells (9).
In conclusion, we could demonstrate that high concentrations of HA change the fibrinolytic properties of mesothelial cells. At concentrations up to 30 mg/dL, we did not find these alterations in t-PA and PAI-1 expression. Consequently, the supplementation of HA in these low concentrations to PD solutions may not have a negative impact on the mesothelial fibrinolytic capacity. Nevertheless, it should never be forgotten that HA plays a key role in wound healing and fibrotic processes.
Footnotes
Acknowledgment
This work constitutes part of the doctoral thesis of Matthias Sauter, University of Munich.