
Abstract
Background:
The alpha-1 antitrypsin (AAT) protein has an important role in the anti-inflammatory and apoptotic response. AAT inhibits not only serine proteases but also cysteine and aspartic proteases. Apoptosis results from the sequential activation of cysteine proteases of the caspase family. This study aimed to evaluate the effect of AAT on formaldehyde-induced apoptosis of human peritoneal mesothelial cells (HPMCs).
Methods:
HPMCs were cultured and treated with formaldehyde (250 µM) to induce apoptosis. In the AAT group, the cultured HPMCs were pretreated with AAT (2 mg/mL) for 1 h before formaldehyde treatment. We used 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assays to determine cell viability, and flow cytometry and terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assays to detect apoptosis. The MTT assays were used to find optimal concentrations of formaldehyde and AAT. We measured caspase-3 activity and used Western blotting to estimate Bcl-2 and Bad expression.
Results:
Flow cytometry and TUNEL assays revealed that formaldehyde exposure significantly increased apoptosis compared with the control treatment, but pretreatment with AAT significantly inhibited this effect. Compared with the control, caspase-3 activity was significantly increased and the ratio of Bcl-2 to Bad expression significantly decreased following treatment with formaldehyde. However, caspase-3 activity was significantly lower and the Bcl-2 to Bad expression ratio higher in the AAT group than in the formaldehyde-only group.
Conclusion:
AAT inhibits formaldehyde-induced apoptosis of HPMCs via a caspase-mediated pathway. These data support a potential use for AAT as a therapeutic agent for the inhibition of peritoneal cell apoptosis during peritoneal dialysis.
Keywords
Introduction
Alpha-1 antitrypsin (AAT) is known as an acute-phase protein and an abundant serine protease inhibitor, such as neutrophil elastase, cathepsin G, and proteinase 3, in the circulation. 1 In recent studies, it has been also known that the ability of AAT to inhibit not only a broader range of serine proteases but also other classes of proteases, such as cysteine and aspartic proteases. 2,3 Therefore, AAT can effectively regulate inflammatory response than inhibits only specific proteases.
Apoptosis, the process of programmed cell death, is a vital component of various processes including inflammation. 4,5 The sequential activation of cysteine proteases of the caspase family causes apoptosis. 6 Among caspases, especially, casepase-3 plays a crucial role in apoptosis, because it is frequently activated and catalyzes the specific cleavage of various key cellular proteins. 7 In the previous study, AAT directly binds to caspase-3, thus it can inhibit caspase-3-mediated apoptosis. 8
Peritoneal dialysis (PD) is an essential renal replacement therapy for patients with chronic kidney disease, and its efficacy depends on the functional and structural integrity of the peritoneum. 9 However, chronic exposure of PD fluids that contain high glucose concentrations and glucose degradation products (GDPs) results in apoptosis of human peritoneal mesothelial cells (HPMCs) and reduce cell proliferation. 10 It has been known that there are many kinds of GDPs in PD fluids, such as formaldehyde, methylglyoxal, and 5-hydroxymethylfuraldehyde, 11,12 and we conducted preliminary experiments using these GDPs to make the most appropriate apoptosis model (data not shown). In the preliminary study, formaldehyde resulted in lower cell viability than other GDPs, and previous studies have confirmed that formaldehyde induces apoptosis. 13,14 Therefore, we studied the formaldehyde-induced apoptosis model. The anti-inflammatory effects of AAT have been identified in a mouse peritoneum model, 15 but the anti-apoptotic property of AAT, the protective effect of this protein on the peritoneal membrane is not yet known. The objective of this study was to investigate the effect of AAT on formaldehyde-induced apoptosis of HPMCs.
Methods
Primary culture of HPMCs
Human omental tissue was obtained from patients undergoing abdominal surgery, who provided their informed consent. HPMCs were isolated by enzymatic disaggregation with 0.25% trypsin-EDTA (Life Technologies, Carlsbad, California) for 30 min at 37°C water bath with continuous rotation. After incubation, digested omental tissue were discarded and the suspension was centrifuged at 1500 r/min for 3 min. The cell pellet was washed twice in phosphate-buffered saline (PBS), cultured in the Medium 199 supplemented with 20% fetal bovine serum (FBS), 100 Unit/mL penicillin, and 100 µg/mL streptomycin at 37°C, 5% CO2 conditions. To confirm the HPMC population, we usually tested the HPMC population via immunofluorescence staining of cytokeratin and a-SMA. 16 HPMCs between the second and fourth passages were used in the experiments. The study protocol was reviewed and was approved by the Institutional Review Board of the Kyungpook National University Hospital (KNU-2016-02-034).
Formaldehyde treatment
Cultured HPMCs were seeded at 80% confluence in Medium 199 containing 20% FBS and incubated with 1% FBS-containing Medium 199 for 24 h to synchronize cell growth. HPMCs were then pretreated with or without 2 mg/mL AAT (Aralast®; Baxter Healthcare, Vienna, Austria) for 1 h and subsequently treated with or without 250 µM formaldehyde (Sigma, St. Louis, Missouri, USA) for 72 h to investigate the apoptotic cells using the microscope, flow cytometric analysis, terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay, and caspase-3 activity.
Cell viability assay
To investigate cell viability in the presence of AAT or formaldehyde, HPMCs were seeded in 96-well plates and treated with various concentrations of AAT (1–10 mg/mL) or formaldehyde (30–500 µM) for 24, 48, or 72 h. The treated cells were then incubated with 500 µg/mL 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Amresco, Solon, Ohio, USA) 17 for 4 h. The converted dye was solubilized with 200 µL dimethyl sulfoxide (Amresco), and absorbance at 570 nm was measured using a microplate reader (Model 550; Bio-Rad, Hercules, California, USA). The results are expressed as percentages of the control value.
Assessment of cell apoptosis
Flow cytometric analysis of apoptosis was achieved using a Fluorescein Isothiocyanate Annexin V Apoptosis Detection Kit (BD Biosciences, Franklin Lakes, New Jersey, USA). After treatment, the cells were trypsinized and gently centrifuged. The cell pellet was resuspended in 100 µL 1× annexin binding buffer and incubated with 5 µL annexin V and 5 µL propidium iodide (PI) for 15 min in the dark. After adding 200 µL PBS, the cells were analyzed in a FACScan flow cytometer (BD Biosciences). The ratio of annexin V-positive and/or annexin V/PI double-positive cells to total cells was used to calculate the percentage of apoptotic cells.
Apoptosis was also investigated with the TUNEL assay using an In situ Cell Death Detection Fluorescein Kit (Roche, Mannheim, Germany). Briefly, the treated cells were fixed in 4% paraformaldehyde for 1 h at room temperature and subsequently permeabilized in 0.1% Triton X-100/0.1% sodium citrate for 2 min at 4°C. After being washed with PBS, the cells were incubated for 1 h at 37°C with 50 µL TUNEL reagent mixture and counterstained for 1 min with 4′,6-diamidino-2-phenylindole (DAPI; Sigma) to visualize nuclei. Finally, the cells were mounted in ProLong Gold antifade reagent (Invitrogen, Eugene, Oregon, USA) and observed under a confocal microscope (Carl Zeiss, Göttingen, Germany). The number of TUNEL-positive cells was counted in three to five randomly selected slides per experiment. The ratio of TUNEL-positive to DAPI-positive cells was used to calculate the percentage of apoptotic cells.
Measurement of caspase-3 activity
After 72 h of treatment, caspase-3 activity was measured using a colorimetric caspase-3 assay kit (Sigma) according to the manufacturer’s protocol. Treated HPMCs were lysed in lysis buffer for 15 min on ice. The cell lysate was reacted with the fluorometric caspase-3 substrate N-acetyl-Asp-Glu-Val-Asp p-nitroanilide in assay buffer. To test for nonspecific hydrolysis of substrate, a control reaction mixture was exposed to the caspase-3 inhibitor acetyl-Asp-Glu-Val-Asp-CHO in assay buffer. Both mixtures were incubated for 90 min at 37°C before measuring absorbance at 405 nm using a microplate reader (Bio-Rad Model 550).
Immunoblotting of Bcl-2 and Bad
After treatment with AAT and formaldehyde, HPMCs were lysed in ice-cold radioimmunoprecipitation assay buffer (50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, and 1% NP-40) containing Protease Inhibitor Cocktail Set III (Calbiochem, Darmstadt, Germany). After centrifugation at 12,000 × g for 15 min at 4°C, the concentration of cell lysates was determined by Bradford assay. Twenty micrograms of cell lysate were separated on a 12% sodium dodecyl sulfate-polyacrylamide gel and transferred to a nitrocellulose membrane. The membrane was blocked for 1 h and incubated overnight at 4°C with primary antibodies against Bcl-2 (1:1000; Cell Signaling Technology, Danvers, Massachusetts, USA), Bad (1:1000; Cell Signaling Technology), and β-actin (1:5000, Sigma). After washing, the membrane was incubated with horseradish peroxidase-conjugated secondary antibodies (1:10,000; Dako, Glostrup, Denmark) for 1 h and antibody binding was visualized using enhanced chemiluminescence advanced detection reagents (GE Healthcare, Little Chalfont, UK). The intensity of the bands was quantified using Scion Image software (Scion, Frederick, Maryland, USA).
Statistical analysis
Data are presented as means ± standard errors of the means and derive from at least three independent experiments. Statistical analysis, comprising one-way analysis of variance and Tukey’s post hoc test, was performed using SPSS version 20.0 (SPSS, Chicago, Illinois, USA). p Values less than 0.05 were considered statistically significant.
Results
Effect of AAT on HPMC viability
The effect of AAT on HPMC viability was evaluated using the MTT assay. HPMCs were treated with different concentrations of AAT (1, 2, 5, or 10 mg/mL) for 24, 48, or 72 h. Their viability was significantly decreased by exposure to 5 and 10 mg/mL AAT for 72 h (Figure 1(a)). Therefore, subsequent experiments were performed with 2 mg/mL AAT, a concentration at which viability was not affected.
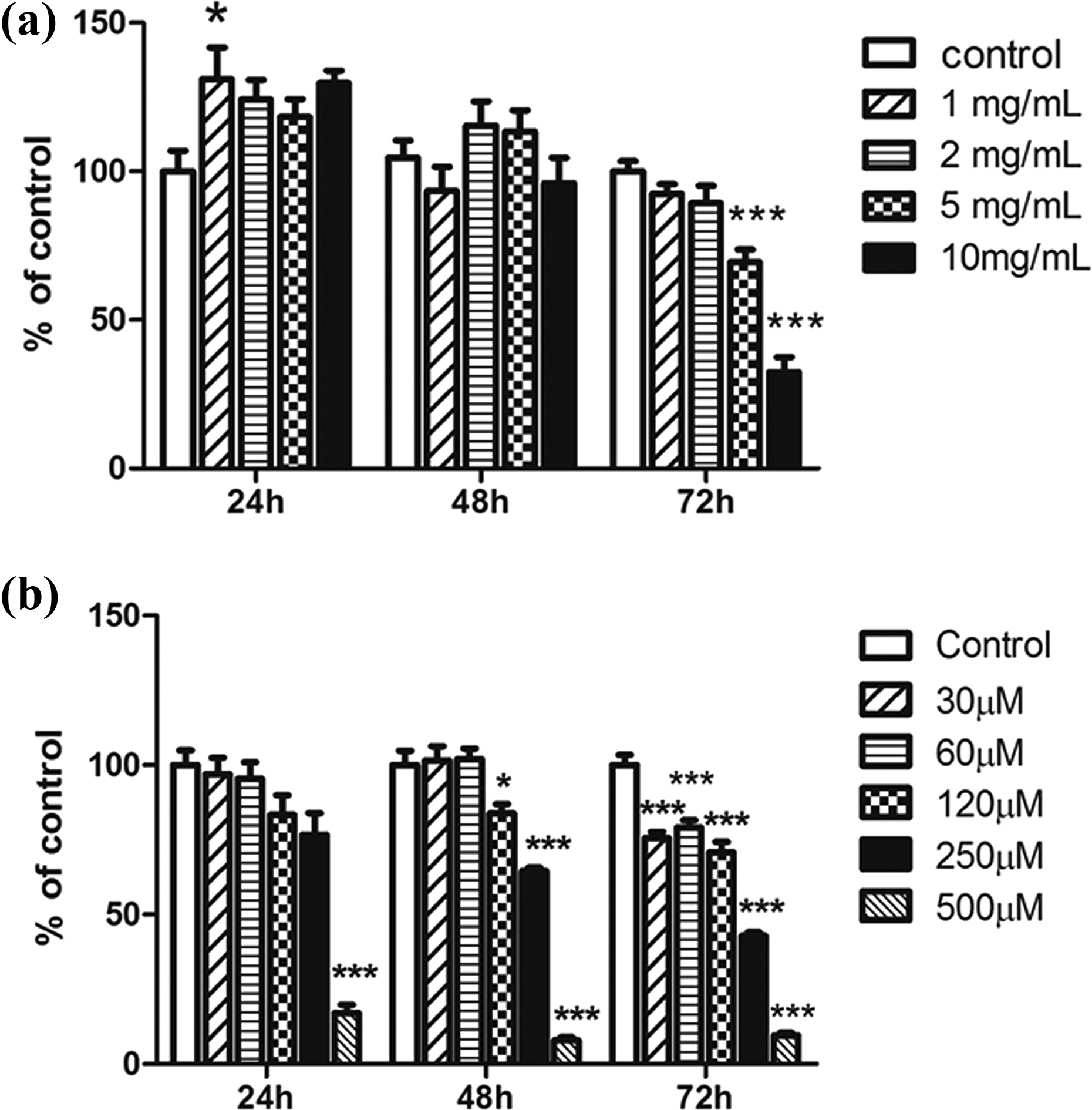
Cell viability determined using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. (a) HPMCs were treated with AAT (1, 2, 5, or 10 mg/mL) for 24, 48, or 72 h. (b) HPMCs were treated with formaldehyde (30–500 µM) for 24, 48, or 72 h. Data are presented as means ± standard errors of the means of three or more independent experiments; *p < 0.05 and ***p < 0.001 compared to the control.
Formaldehyde-induced HPMC death
Cell viability was determined at different formaldehyde concentrations and exposure times by the MTT assay. HPMCs were treated with formaldehyde (30–500 µM) for 24, 48, or 72 h. Although significant decreases in cell viability were noted at concentrations of 120 µM and above after 48 and 72 h, a 50% reduction in viability was observed following exposure to 250 µM formaldehyde for 72 h (Figure 1(b)). This concentration and exposure time were therefore used in the experiments that followed.
Morphological changes in HPMCs
Figure 2 shows the morphological changes of HPMCs before and after treating formaldehyde alone or with AAT. After formaldehyde treatment, HPMC viability was decreased and cells displayed apoptotic changes. However, pretreatment with AAT in HPMCs improved cell viability and decreased apoptotic changes.

Morphological changes of HPMCs. HPMCs were cultured and treated with formaldehyde (250 µM) for 72 h to induce apoptosis. In the AAT group, HPMCs were pretreated with AAT (2 mg/mL) for 1 h before formaldehyde treatment (original magnification ×100).
AAT treatment inhibited HPMC apoptosis
To determine the effect of AAT on formaldehyde-induced apoptosis, we performed an annexin V/PI assay using flow cytometry. The percentage of apoptotic HPMCs was calculated as the ratio of annexin V-positive and/or annexin V/PI double-positive cells to total cells (Figure 3). In the nontreated control groups, 2.8% of cells were annexin V+/PI−, whereas 11.7% were annexin V+/PI+. After induction of apoptosis with formaldehyde, a significant increase in the numbers of early and late apoptotic cells was identified, with 23.6% being annexin V+/PI− and 16.7% annexin V+/PI+. However, HPMC apoptosis was less frequent in the AAT pretreatment group than the formaldehyde-only group, with 14.9% of cells in the former being annexin V+/PI− and 13.5% annexin V+/PI+. Quantitative analysis revealed that the percentage of early, late, and total apoptotic cells was higher in the formaldehyde-treated group than the control group. However, the percentage of apoptotic cells was significantly lower in the AAT pretreatment group than the formaldehyde-treated group.
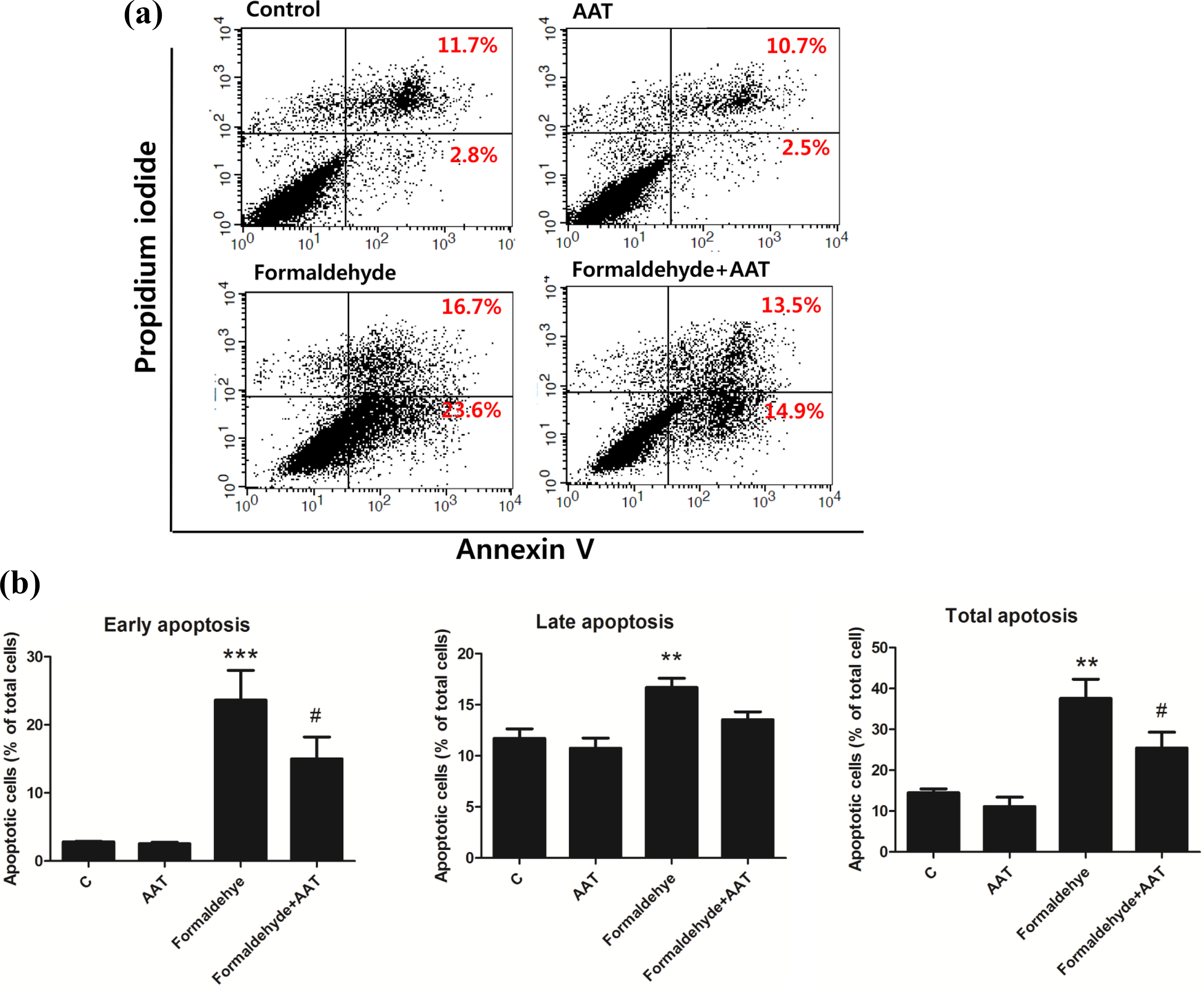
Apoptotic cells were detected using flow cytometric analysis of annexin-V and PI staining. Human peritoneal mesothelial cells were pretreated with AAT (2 mg/mL) for 1 h and subsequently exposed to formaldehyde (250 µM) for 72 h. (a) Dot plot showing the control, 2 mg/mL AAT, 250 µM formaldehyde, and 2 mg/mL AAT + 250 µM formaldehyde groups. (b) Quantitative analysis of early apoptotic (annexin-V+/PI−), late apoptotic (annexin-V+/PI+), and total apoptotic (early and late apoptotic) cells. Data are presented as means ± standard errors of the means of three or more independent experiments; **p < 0.01 and ***p < 0.001 compared to the control; #p < 0.05 compared to the formaldehyde-only group.
We used the TUNEL assay to determine the effect of AAT on formaldehyde-induced apoptosis of HPMCs. The apoptosis rate was calculated as the percentage of TUNEL-positive cells following AAT and/or formaldehyde treatment (Figure 4). The TUNEL assay revealed that the rate of apoptosis was significantly higher among formaldehyde-treated cells than control cells and was lower in the 2 mg/mL AAT pretreatment group than the formaldehyde-only group (p < 0.001).
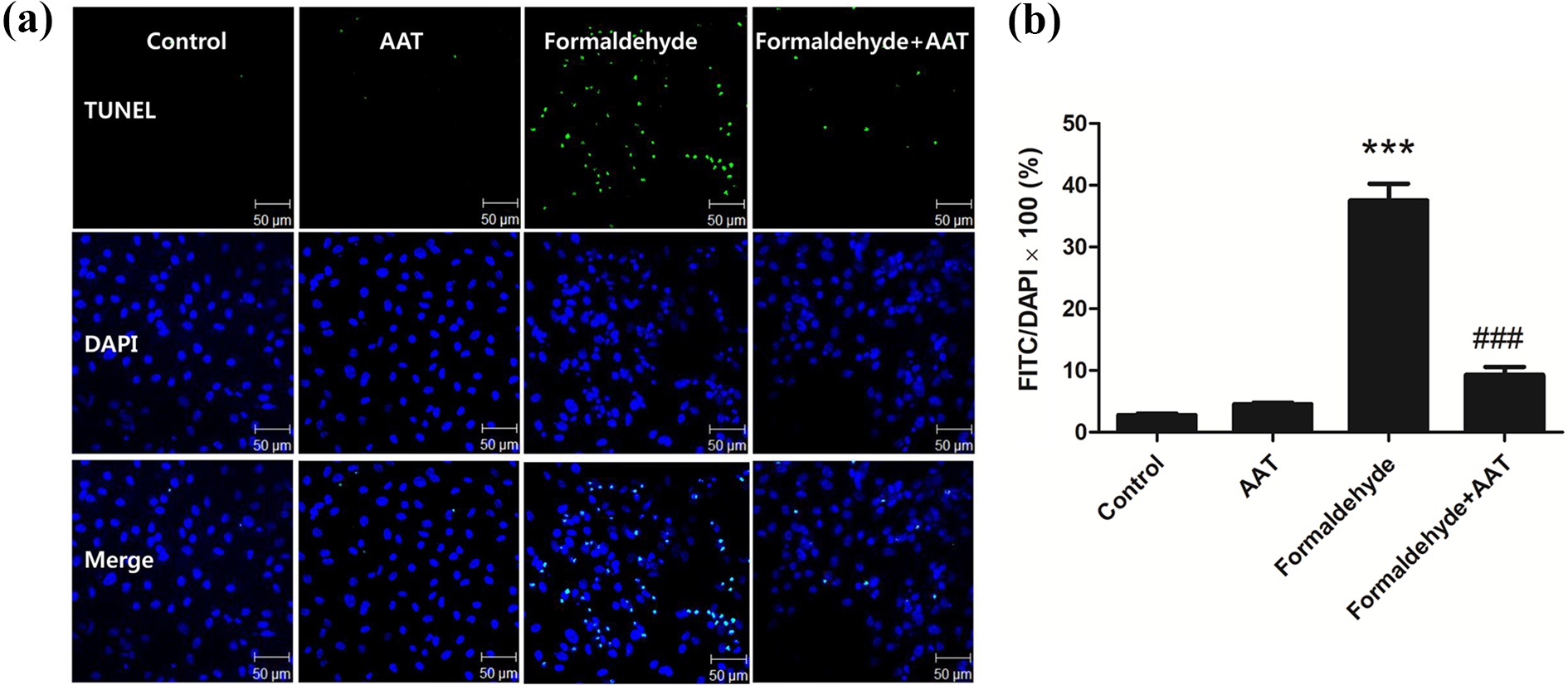
HPMC apoptosis detected by TUNEL. HPMCs were pretreated with AAT (2 mg/mL) for 1 h and subsequently treated with formaldehyde (250 µM) for 72 h. The cells were then stained with TUNEL reagent and DAPI. (a) The results were visualized using a confocal microscope (×200 magnification). (b) The apoptosis rate was calculated as the percentage of TUNEL-positive cells following AAT and/or formaldehyde treatment. Data are presented as means ± standard errors of the means of three or more independent experiments; ***p < 0.001 compared to the control; ###p < 0.001 compared to the formaldehyde-only group. HPMC: human peritoneal mesothelial cell; TUNEL: terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling; AAT: alpha-1 antitrypsin; DAPI: 4′,6-diamidino-2-phenylindole.
Caspase-3 activity during HPMC apoptosis was attenuated by AAT treatment
We examined the activity of caspase-3, the main effector caspase, in HPMCs collected after 72 h of formaldehyde and/or AAT treatment. Caspase-3 activity was significantly higher in the formaldehyde group than the control group (p < 0.001) and lower in the AAT pretreatment group than the formaldehyde group (p < 0.01) (Figure 5(a)).
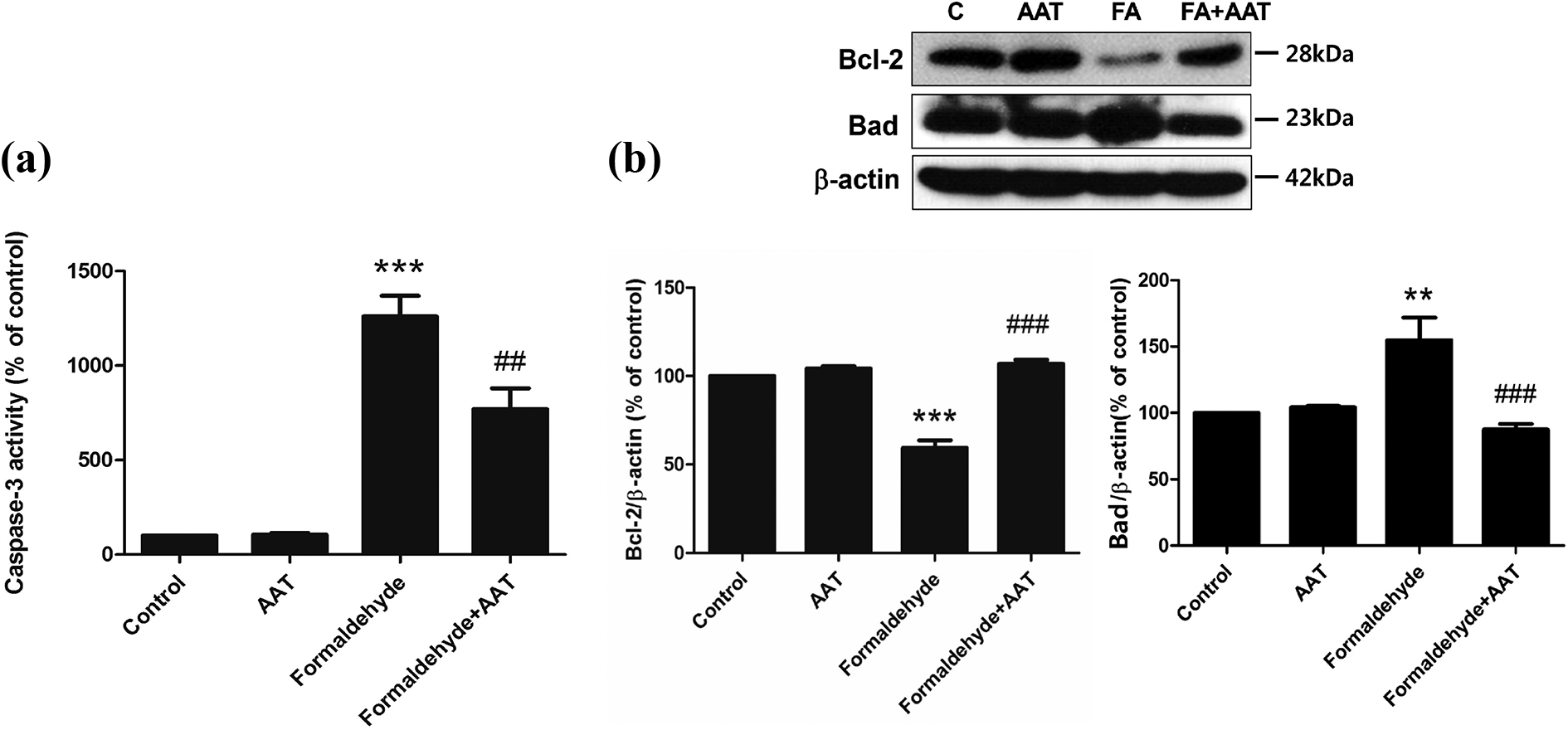
Analysis of caspase-3 activity (a), and Bcl-2 and Bad protein expression (b). (a) Formaldehyde treatment for 72 h increased caspase-3 activity in HPMCs; however, this effect was significantly diminished by pretreatment with AAT. (b) HPMCs were pretreated with AAT (2 mg/mL) for 1 h and subsequently treated formaldehyde (250 µM) for 72 h. The sample sizes of each group were all six. The upper figure represents Western blots of Bcl-2 and Bad. C: control; FA: formaldehyde. The lower results are expressed as percentages of the control values. All data are presented as means ± standard errors of the means of three or more independent experiments; **p < 0.01 and ***p < 0.001 compared to the control; ##p < 0.01 and ###p < 0.001 compared to the formaldehyde-only group. HPMC: human peritoneal mesothelial cell; AAT: alpha-1 antitrypsin.
Bcl-2 and Bad expression mediated formaldehyde-induced apoptosis of HPMCs
In order to investigate the downstream pathway involved in the mitigation of formaldehyde-induced HPMC apoptosis by AAT, we conducted Western blotting analysis of Bcl-2 and Bad expression (Figure 5(b)). This revealed that formaldehyde treatment downregulated expression of Bcl-2 and upregulated that of Bad. Moreover, Bcl-2 and Bad levels were found to be higher and lower, respectively, in the AAT pretreatment group than the formaldehyde-only group (p < 0.001).
Discussion
In our study, we demonstrated that in vitro administration of AAT attenuated formaldehyde-induced apoptosis of HPMCs. The number of apoptotic cells was significantly lower in the AAT pretreated group than the formaldehyde-only group. In addition, AAT inhibited several apoptotic pathways, as evidenced by its effects on caspase-3 activity and Bcl-2 and Bad levels.
Recent studies have revealed that AAT not only inhibits serine proteinases secreted by neutrophils but also controls the inflammatory process in the peritoneum. 18 AAT expression differs among chronic PD patients with different types of peritoneal membrane. 19 Moreover, AAT recovered from PD effluents has been shown to be biologically active, inhibiting elastase activity and the synthesis of platelet-activating factor. 20 However, whether AAT modulates cell apoptosis, especially that of human peritoneal cells, is not known. We have shown here that apoptosis of cultured HPMCs induced by formaldehyde is mitigated by AAT pretreatment.
Glucose, which constitutes the main osmotic factor in PD fluids, is unstable when exposed to heat. 21 Thus, glucose in PD fluids breaks down into other molecules during the heat sterilization procedure. 22 GDPs are one of the most critical influences on the biocompatibility of glucose-based PD solutions. The concentrations of diverse GDPs differ depending on the temperature and pH during the sterilization and storage processes. GDPs in PD fluid impair peritoneal mesothelial cell function and promote the formation of advanced glycation end-products (AGEs). Carbonyl compounds deriving from several degradation cascades are also well-known sources of irreversible AGEs. 23 AGEs might be associated with histopathologic changes leading to loss of ultrafiltration and, eventually, remodeling of the peritoneal membrane. 24,25 Long-term exposure to bioincompatible dialysate components during PD thus causes variable side effects due to loss of peritoneal membrane cell structure or function. 26 Formaldehyde, which is among the GDPs chemically verified as being present in PD fluids, passes easily through the cell membrane to immediately cross-link certain intracellular components. 11 Formaldehyde is associated with increased cellular injury and may contribute to the peritoneal membrane damage that occurs in patients undergoing PD. 27 Moreover, it may react with the mesothelial cells lining the peritoneal cavity and could be responsible for their disappearance during long-term treatment.
In this study, we showed that the range of cellular targets of formaldehyde results in the induction of HPMC apoptosis and demonstrated that AAT can attenuate this effect. Aldehydes are associated with increased cellular injury and may contribute to peritoneal membrane damage among patients treated with PD. 27 In our study, the range of formaldehyde concentrations that induced apoptosis of HPMCs was found to be 30–500 µM, as determined by MTT assays. Legge and Thorell reported the concentration of formaldehyde in PD solution between 6 and 15 µM. 28,29 In fact, patients with PD are exposed lifelong to these low concentration solutions. However, in vitro experiment is difficult to conduct a long-term exposure at low concentration, so to create a similar condition, we applied high-concentration and short-term exposure of formaldehyde to make an apoptosis model. Our fluorescence-activated cell sorting and TUNEL assay revealed that the number of apoptotic cells was significantly lower in the AAT pretreatment group than the formaldehyde-only group. The protective effect of AAT demonstrated in the present study indicates that this protein may be used to reduce HPMC apoptosis and facilitate the management of peritoneal membrane damage in a variety of conditions.
Caspases act as initiators and effectors of apoptosis and are activated by intrinsic (mitochondrial) and extrinsic (death receptor) pathways. 30 Initiator caspases proteolytically activate effectors, such as caspase-3, which in turn cleave other intracellular proteins, ultimately leading to cell death. In a prior study, Buchel et al. demonstrated that the caspase-3 pathway is a major mechanism underlying cell apoptosis in a renal injury model and is upregulated by PD fluids. 31 Moreover, in animal studies, caspase inhibition has been shown to exert remarkable effects on organ injury by preventing apoptosis in renal ischemia. 32,33 In our study, we investigated the effect of AAT on caspase-3 activity by using a colorimetric analysis in HPMC apoptosis. The colorimetric analysis in the measurement of caspase-3 activity has some advantages: (1) In our previous studies, 34 –36 we confirmed that the colorimetric analysis reduced time spent when compared to Western blot assay for caspase-3 cleavage; (2) the colorimetric analysis does not require fluorescence or luminescence readers or X-ray film to detect of cleaved caspase-3 band; (3) the colorimetric analysis is more high-throughput experiment than Western blot by quantification of caspase-3 activity by fluorescent in a 96-well plate. Caspase-3 activity was significantly increased in the formaldehyde-only group compared with the control. However, pretreatment with AAT significantly decreased this effect. This result supports the notion that AAT pretreatment may decrease HPMC apoptosis through caspase-3 inactivation.
To clearly assess the AAT effect and mechanism on apoptosis, we also compared the protein levels of Bcl-2 and Bad. Bcl-2 family members, including Bcl-2 and Bad, reside upstream of apoptotic pathway and regulate the mitochondrial pathway in the cell death pathways. 30,37 The viability of cells is largely determined by the interplay between proteins of the Bcl-2 family 38 and the ratio of pro-apoptotic (Bad) to anti-apoptotic (Bcl-2) molecules is a major checkpoint in the apoptotic pathway. 37 We analyzed the protein expression by Western blotting, finding that formaldehyde treatment downregulated the Bcl-2 expression and upregulated the Bad expression. However, compared with formaldehyde treatment only, pretreatment with AAT significantly increased expression of Bcl-2 and decreased that of Bad. As a result, the ratio of Bcl-2 to Bad expression was higher in the AAT pretreatment group than the formaldehyde group. These results indicate that AAT treatment inhibit apoptosis by also affecting the Bcl-2 family of mitochondrial apoptosis pathway.
Conclusions
In the present study, formaldehyde, which is present in PD fluids, induces apoptosis of HPMCs. Furthermore, pretreatment with AAT inhibits formaldehyde-associated HPMC apoptosis and this protective effect is related to caspase activity and the Bcl-2 family signaling pathway. This indicates that AAT might serve as a therapeutic agent to inhibit the pathway responsible for peritoneal mesothelial cell apoptosis during PD. Further animal and clinical studies are needed to confirm the effects of AAT in inhibiting apoptosis of peritoneal mesothelial cells and preserving peritoneal membrane function.
Footnotes
Author contributions
Jeong-Hoon Lim and Sang Mi Park contributed equally to this study.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute, funded by the Ministry of Health & Welfare, Republic of Korea (HI15C0001) and supported by Medi-Start Up Program funded by the Daegu Metropolitan City (Project Name: Development of the candidate agents for acute kidney injury through the reduction of oxidative stress). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.