
Abstract
Trigeminal nerve pathology can lead to sensory and motor dysfunction to structures of the head that are easily recognized. The trigeminal nerve is a conduit for the distribution of postganglionic parasympathetic innervation to structures of the head. Parasympathetic innervation to the salivary glands is provided by preganglionic parasympathetic neurons of the facial and glossopharyngeal nerves. Postganglionic axons course with branches of the mandibular branch of the trigeminal nerve to reach the salivary glands. Denervation of the salivary glands impacts glandular function, leading to a reduction in the volume and composition of the saliva produced. Saliva plays an important role in oral health. Poor oral health has widespread systemic implications. This article describes a group of dogs with unilateral or bilateral dysfunction of the trigeminal nerve and/or its branches. In all dogs, an accumulation of thick, foamy saliva was observed accumulating in the dorsal aspect of the caudal oral cavity on the ipsilateral side to the affected nerve. In dogs with magnetic resonance imaging (MRI), there was a reduction in size based on the largest cross-sectional area measurement and an increase in mean signal intensity of the salivary glands ipsilateral to the affected nerves compared to the glands on the normal side. The authors hypothesize that the abnormal saliva and MRI changes observed were consequent to parasympathetic denervation of the salivary glands. The recognition of this clinical observation is the first step in understanding the impact that denervation has on salivation and ultimately on overall oral and systemic health in dogs.
Introduction
The trigeminal nerve, or the fifth cranial nerve, has 3 major branches: the ophthalmic, maxillary, and mandibular nerves. All 3 main branches of the trigeminal nerve provide somatic sensory function to the majority of the head. Only the mandibular nerve provides somatic motor innervation to the muscles of mastication (masseter, temporalis, lateral and medial pterygoids, and rostral belly of the digastricus), mylohyoideus, and tensor veli palatini muscles. 1,2 Disease involving the trigeminal nerve or its main branches often leads to dramatic clinical signs that are easily recognized in dogs. Most conspicuous is muscle atrophy secondary to denervation of the temporalis and masseter muscles that is observed with dysfunction of the trigeminal and/or mandibular nerve. Atrophy of the pterygoid muscles likely results in enophthalmos and consequent elevation of the third eyelid. Bilateral dysfunction of the mandibular nerve results in an inability to close the mouth with subsequent loss in the ability to prehend and chew food or drink water normally. This is colloquially referred to as a dropped jaw. Dysfunction of the trigeminal nerve or its main branches may also result in loss of cutaneous sensation that is appreciated through testing the autonomous zones for each main branch of the trigeminal nerve. 3 Beyond sensory and motor function, the mandibular branch of the trigeminal nerve provides an important conduit for the postganglionic parasympathetic axons to be distributed to the major salivary glands (parotid, mandibular, zygomatic, and sublingual). 1
The origins of the parasympathetic innervation to the salivary glands are provided by the cell bodies for the preganglionic parasympathetic neurons of the facial and glossopharyngeal nerves. 1 As a conduit, the mandibular nerve simply provides a means by which parasympathetic axons are distributed to the salivary glands. Although parasympathetic axons travel along with the mandibular nerve, the trigeminal nerve itself does not, per se, provide parasympathetic innervation. For the mandibular and sublingual glands, the cell bodies for the preganglionic parasympathetic neurons are located in the parasympathetic nucleus of the facial nerve. 1 Preganglionic axons course through the facial nerve, where they eventually join with the lingual nerve, a branch of the mandibular nerve, to synapse in the mandibular ganglion or sublingual ganglion. Postganglionic axons innervate the mandibular and sublingual glands, respectively. 1 For the parotid and zygomatic glands, the cell bodies for the preganglionic parasympathetic neurons are located in the parasympathetic nucleus of the glossopharyngeal nerve. 1 Preganglionic axons course through the glossopharyngeal nerve to synapse at the otic ganglion. 1 Postganglionic axons join with the auriculotemporal nerve, a branch of the mandibular nerve, to innervate the parotid and zygomatic glands. 1
Little information exists on the clinical consequence of loss of parasympathetic innervation with dysfunction of the trigeminal or mandibular nerve in dogs. Commonly encountered disorders causing trigeminal nerve dysfunction in dogs include trigeminal neuropathy of inflammatory, infectious, and neoplastic etiologies. 4 Beyond thorough physical and neurological examinations, the diagnostic evaluation of affected dogs often includes a minimum database (a complete blood count, serum biochemistry, and urinalysis), magnetic resonance imaging (MRI) of the head, and analysis of cerebrospinal fluid.
Disorders involving the trigeminal or mandibular nerve are anticipated to result in autonomic dysfunction of the organs such as the salivary glands. While diagnostic testing is aimed at establishing an etiology from which to direct therapy and prognosticate recovery, the loss of parasympathetic innervation to structures of the head likely goes unrecognized in dogs with dysfunction of the trigeminal nerve or its main branches. In human beings, loss of normal salivation results in a significant detrimental impact on oral heath 5 as well as quality of life. 6 The impact of autonomic dysfunction to the salivary glands may be challenging to detect in dogs that are not able to self-report symptoms. As in human beings, loss of normal salivation likely negatively impacts oral health in dogs. Poor oral health has widespread consequences in dogs. 7 -10
While the detection of abnormalities directly related to loss of parasympathetic innervation may be challenging to perceive in dogs, the loss of normal physiological processes such as salivation likely affects the character of saliva and therefore may serve as a biomarker for autonomic nervous system dysfunction. The purpose of the following report is to document the observation of an accumulation of thick, foamy saliva that adheres on the mucosa of the dorsocaudal aspect of the oral cavity in dogs with clinical signs related to dysfunction of the trigeminal or the mandibular nerves. Moreover, in dogs with unilateral dysfunction of the trigeminal or the mandibular nerve, the abnormal saliva accumulates on the side ipsilateral to the affected nerve in the area of the openings of the ducts of the parotid and zygomatic salivary glands. The authors hypothesize that trigeminal or mandibular nerve dysfunction leads to denervation of the ipsilateral salivary glands related to loss of parasympathetic postganglionic axons that travel with branches of the mandibular nerve, resulting in the accumulation of abnormal appearing saliva in the oral cavity. Additionally, there is a reduction in size of the salivary glands based on the largest cross-sectional area measurement and an increase in mean signal intensity of the salivary glands on MRI suggestive of structural changes within the denervated salivary glands.
Methods
Affected dogs were examined from 2013 through 2017 at Red Bank Veterinary Hospital (RBVH) and the Veterinary Teaching Hospital at the University of Georgia (UGA). Dogs with clinical signs related to dysfunction of the trigeminal nerve or the mandibular nerve were eligible for inclusion. Signs consistent with dysfunction of the trigeminal nerve or its main branches included atrophy of the muscles of mastication (primarily the temporalis and masseter muscles), loss of cutaneous sensation as evidenced by absent or reduced response to tactile stimulation of the mucosa of the nasal septum (response to stimulation of the mucosa with the closed end of a forceps), and absent or reduced palpebral and corneal reflexes with preservation of the menace response and the voluntary ability to close the palpebral fissure. For the purpose of classifying cutaneous sensory loss, a reduced or absent palpebral reflex when stimulating the lateral canthus of the eye was considered related to the maxillary nerve, a reduced or absent palpebral reflex when stimulating the medial canthus of the eye was considered related to the ophthalmic nerve, and a reduced or absent response to stimulation of the mucosa of the rostral most aspect of the nasal septum was also considered related to the ophthalmic nerve. Dogs with neurological dysfunction consistent with a loss of the facial nerve (ie, facial weakness or paralysis) or a loss of sympathetic innervation (ie, Horner syndrome) to the eye were excluded from consideration. Additionally, dogs with a history of cervical or cranial trauma and dogs with a historical or preexisting disease involving the salivary glands or their ducts (ie, sialoadenitis or a salivary mucocele), temporomandibular joints, or the oral cavity were excluded. Dogs receiving any topical, oral, or parenteral medications that have autonomic effects were excluded. Given the prevalence of dental calculus in dogs, dental calculus and/or periodontal disease were not specific exclusion criteria.
All dogs were examined by a board-certified neurologist. In addition to a neurologic examination, patients also had general physical and oral examinations performed. In all dogs, the presence, site, side, and visual characteristics of the accumulation of abnormal saliva within the oral cavity were documented. Additionally, the signalment, neurological findings, imaging results, and etiological diagnosis, when available, were recorded.
In 11 dogs, MRI of the head was performed under routine general anesthesia. Using a 1.5-T MRI unita and multichannel head coil (RBVH) or a 3.0-T MRIb and an extremity coil (UGA), a minimum of the following sequences was obtained in the transverse plane in all dogs: T2-weighted (T2W), fluid attenuated inversion recovery, and T1-weighted (T1W) sequences before and after the intravenous injection 0.1 mmol/kg of gadodiamidec or gadopentetate dimeglumined. Additional sequences were at the discretion of the neurologist supervising the imaging procedure.
The cross sectional area of the parotid salivary gland was measured using digital imaging and communications in medicine (DICOM) softwaree.
In 6 dogs with unilateral dysfunction of the trigeminal or mandibular nerves, the entirety of the parotid, zygomatic, and mandibular salivary glands were visible on the MRI. Using DICOM viewing softwaref, the largest cross-sectional area measurement was obtained for the parotid, zygomatic, and mandibular salivary glands bilaterally. A measurement was obtained for each salivary gland using images from a transverse T2W sequence. The T2W transverse image in which each individual gland from both sides of the head appeared the largest was chosen for area measurements. All area values were recorded in centimeter squared (cm2). The mean signal intensity within the largest cross-sectional area tracing was also recorded for each gland. Additionally, a circular shaped 0.5-cm2 region of interest (ROI) was placed in an area of each gland that subjectively displayed a homogenous signal intensity. The mean signal intensity within the ROI was recorded for each gland. The largest cross-sectional area measurement, the mean signal intensity within the largest cross-sectional area tracing, and the ROI for the parotid, zygomatic, and mandibular salivary glands on the side of the head with dysfunction of the trigeminal and/or the mandibular nerve was compared to the glands on the other side of the head. A linear mixed-effects model was used to compare the largest cross-sectional areas and mean signal intensities between glands on the affected sides of the head and the normal side. Each model had a fixed effect of the side of the head (affected compared to normal) and a random intercept of the individual dog and salivary gland nested within each dog to account for the within dog and within gland correlations. Statistical analysis was performed using commercially available softwareg. A value of P < .05 was considered significant.
In 3 dogs, gross and microscopic evaluation of the abnormal trigeminal nerve was performed as part of a routine necropsy. In 2 of these dogs, gross and histological evaluation of the salivary glands from both sides of the head also was performed. Within 24 hours of humane euthanasia, the parotid, zygomatic, mandibular, and sublingual glands were dissected and removed. The unpreserved tissues were weighed. Weights were reported in grams. The salivary glands were preserved in 10% buffered formalin. Representative portions of the formalin-fixed glands and the affected trigeminal nerve and brain stem were routinely sectioned, processed, and stained with hematoxylin and eosin for microscopic evaluation.
Results
Eleven dogs were identified with unilateral or bilateral dysfunction of the trigeminal and/or its branches. The signalment, neurological examination findings, imaging modality used in diagnostic workup, and diagnosis are presented in Table 1. The age of the dogs ranged from 4 to 14 years. There were 3 spayed females, 7 castrated males, and 1 intact male. The breeds included 2 Labrador retrievers, 2 Cavalier King Charles spaniels, and 1 each of American Eskimo, Boykin spaniel, English bulldog, husky mix, pit bull, pug-beagle mix, and West Highland white terrier.
Signalment, Signs, Imaging Tests, Imaging Findings, and Presumptive Diagnosis.
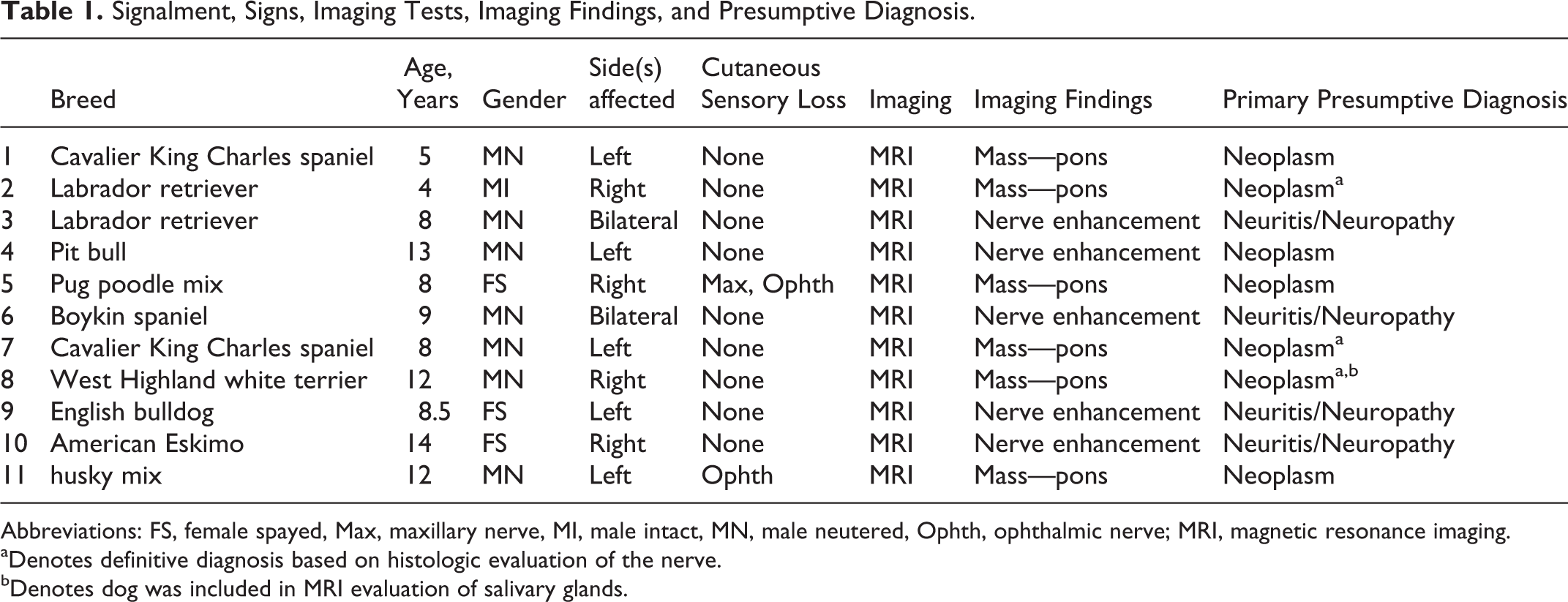
Abbreviations: FS, female spayed, Max, maxillary nerve, MI, male intact, MN, male neutered, Ophth, ophthalmic nerve; MRI, magnetic resonance imaging.
aDenotes definitive diagnosis based on histologic evaluation of the nerve.
bDenotes dog was included in MRI evaluation of salivary glands.
Eleven dogs had mandibular nerve dysfunction (4 dogs with right-sided and 5 dogs with left-sided dysfunction), and 2 dogs had bilateral dysfunction. In all dogs, neurological examination findings disclosed severe atrophy of the temporalis and masseter muscles on the affected side(s) of the head. Other neurological examination findings included decreased cutaneous sensation in the distribution of the ophthalmic nerve (1 dog) and ophthalmic and maxillary nerves (1 dog), unilateral enophthalmos (8 dogs), corneal ulceration (2 dogs), and an inability to voluntarily close the mouth (2 dogs). In one dog (West Highland white terrier), tongue ulceration and excessive dental calculus build up were also present on the ipsilateral side to the affected nerve (Figure 1).
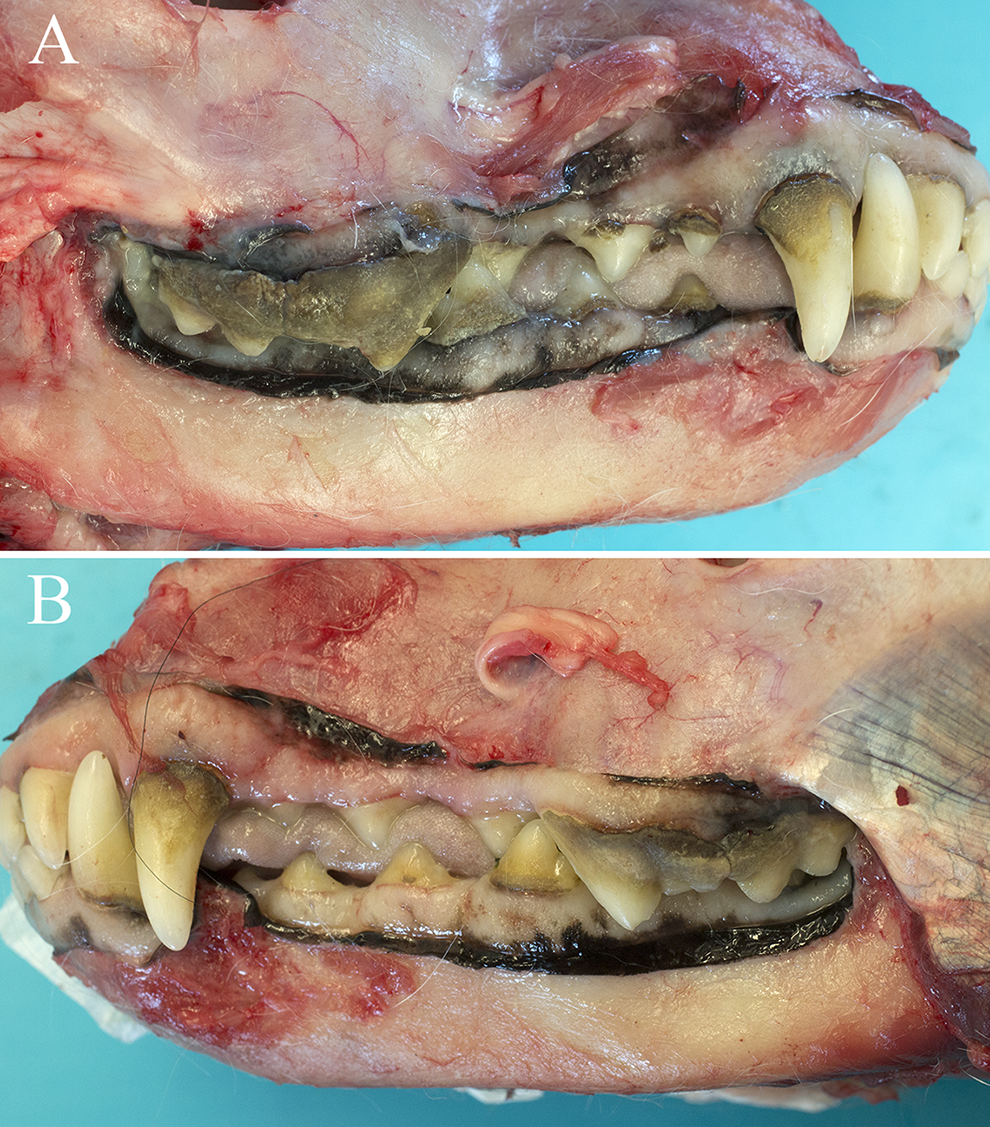
Lateral view of a post mortem preparation of a 12-year-old neutered male West Highland white terrier (dog 8) with right-sided dysfunction of the trigeminal nerve secondary to a nerve sheath neoplasm. (A) Note the accumulation of calculus over the buccal surface of the right maxillary fourth premolar and right maxillary first molar, right maxillary first and second premolars at the gingival margin, and the right mandibular third premolar. (B) In comparison to the right side, there is subjectively less calculus present on the buccal surfaces of the left maxillary first premolar and maxillary first molar as well as the left mandibular premolars.
In all dogs with unilateral neurological deficits, there was accumulation of saliva on the mucosa that lines the dorsal aspect of the oral cavity at the caudal edge of the maxillary fourth premolar on the side ipsilateral to the neurological dysfunction (Figure 2). In comparison to saliva present in other areas of the oral cavity, the saliva adherent to the caudodorsal region of the oral cavity ipsilateral to the neurological deficits subjectively appeared thick, foamy, and more viscous. In the 2 dogs with signs related to bilateral neurological deficits, abnormal accumulation of saliva was observed bilaterally in the same area of the oral cavity as was seen in dogs with unilateral deficits. Additionally, the owners of both dogs had commented on the presence of ropey, foamy saliva that was present in the dogs’ water bowls following drinking. Both owners reported that this began coincident with the onset of neurological signs.
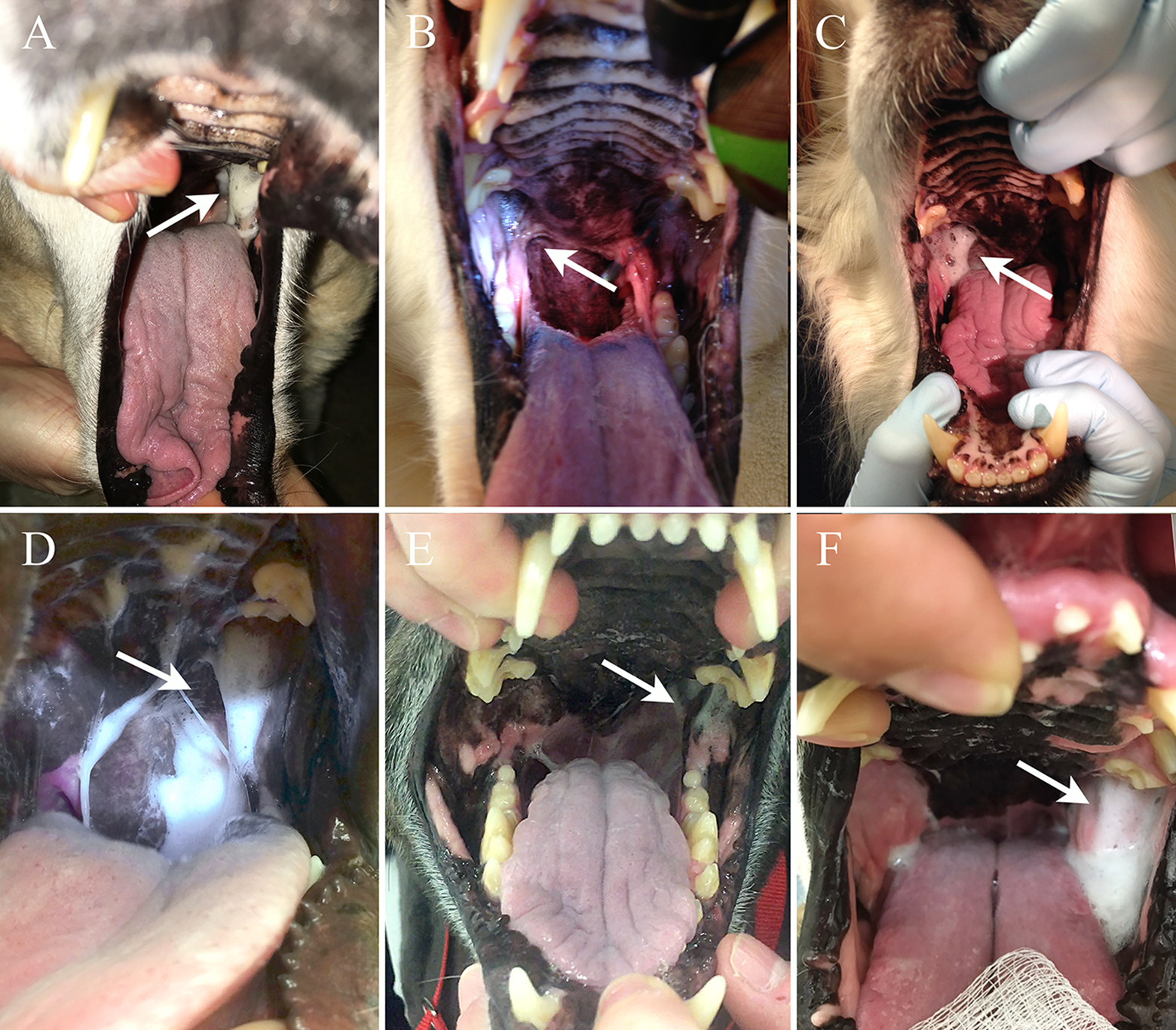
(A-F) Intra-oral views of 6 dogs with dysfunction of the trigeminal and/or mandibular nerve. The dogs in panels (A) and (D and F) have left-sided dysfunction. Dogs in panel (B and C) have right-sided dysfunction. Note the accumulation of thick, foamy saliva just caudal to maxillary molar 2 on the side ipsilateral to the nerve dysfunction.
Eleven dogs underwent cross-sectional imaging for evaluation of dysfunction of trigeminal nerve. Magnetic resonance imaging of the head was performed in 11 dogs. In comparison to the unaffected side, 5 dogs had unilateral trigeminal nerve enlargement within the trigeminal canal, which extended caudally to compress the ipsilateral side of the pons. Three dogs had unilateral trigeminal nerve enlargement within the trigeminal canal, which continued as enlargement of the mandibular nerve as it exited the oval foramen. In these 3 dogs, compression or invasion of the brain stem was not observed. In all dogs with unilateral trigeminal nerve and/or mandibular nerve enlargement, the affected nerves had similar signal intensities on T1W and T2W images as the same nerves on the unaffected side. Moreover, the degree of contrast enhancement of the affected nerves on T1W images was subjectively greater compared to the contrast enhancement of the nerves on the unaffected side. In 1 dog, the trigeminal nerve within the trigeminal canal and the mandibular nerve as it exited the oval foramen on the side with atrophy of the temporalis and masseter muscles was not enlarged; however, it displayed a greater degree of contrast enhancement compared to the same nerves on the unaffected side. In the 2 dogs with bilateral dysfunction of the trigeminal and/or mandibular nerve, the trigeminal nerve and the mandibular nerve were subjectively enlarged and displayed strong and uniform contrast enhancement greater than expected for a normal dog. 11
In all dogs with unilateral dysfunction of the trigeminal and/or mandibular nerve, atrophy and contrast enhancement of the muscles of mastication were present on the side ipsilateral to the affected nerve. Bilateral atrophy and contrast enhancement of the muscles of mastication were present in the 2 dogs with bilaterally enlarged trigeminal nerves. Three dogs had effusion in the tympanic cavity on the side ipsilateral to the affected nerve.
In 6 dogs, the entirety of the parotid, zygomatic, and mandibular glands were visible on both sides of the head on the transverse T2W images (Figure 3). Therefore, the largest cross-sectional area measurement was obtained in a total of 18 salivary glands. On the side ipsilateral to the affected trigeminal nerve and/or mandibular nerve, the largest cross-sectional area measurement of the salivary glands was significantly smaller compared to the glands on the unaffected side (P = .023). On the side ipsilateral to the affected trigeminal nerve and/or mandibular nerve, the largest cross-sectional area measurement was smaller in 12 glands, the same in 2 glands, and larger in 4 salivary glands. In all dogs, the parotid gland was smaller on the side ipsilateral to the trigeminal and/or mandibular nerve dysfunction. On the side ipsilateral to the affected trigeminal nerve and/or mandibular nerve, the mean signal intensity within the largest cross-sectional area tracing and the defined, circular 0.5-cm2 ROI was significantly higher compared to the mean signal intensity of the glands on the unaffected side (P = .022 and P = .024, respectively). On the side ipsilateral to the affected trigeminal nerve and/or mandibular nerve, the mean signal intensity within the largest cross-sectional area tracing was higher in all glands except for 4 glands; 4 dogs had a single gland (3 mandibular and 1 parotid) on the unaffected side of the head with higher mean signal intensity. On the side ipsilateral to the trigeminal and/or mandibular nerve dysfunction, the mean signal intensity of the ROI was higher in 14 glands. In all dogs, the parotid gland on the side ipsilateral to the trigeminal and/or mandibular nerve dysfunction had a higher mean signal intensity in the ROI. Subjectively, the zygomatic glands on the side ipsilateral to the trigeminal and/or mandibular nerve dysfunction often had tissue with signal intensity consistent with fat dissecting through it. Additionally, the ducts within the gland on the side ipsilateral to the trigeminal and/or mandibular nerve dysfunction were often larger than those of the contralateral zygomatic gland.
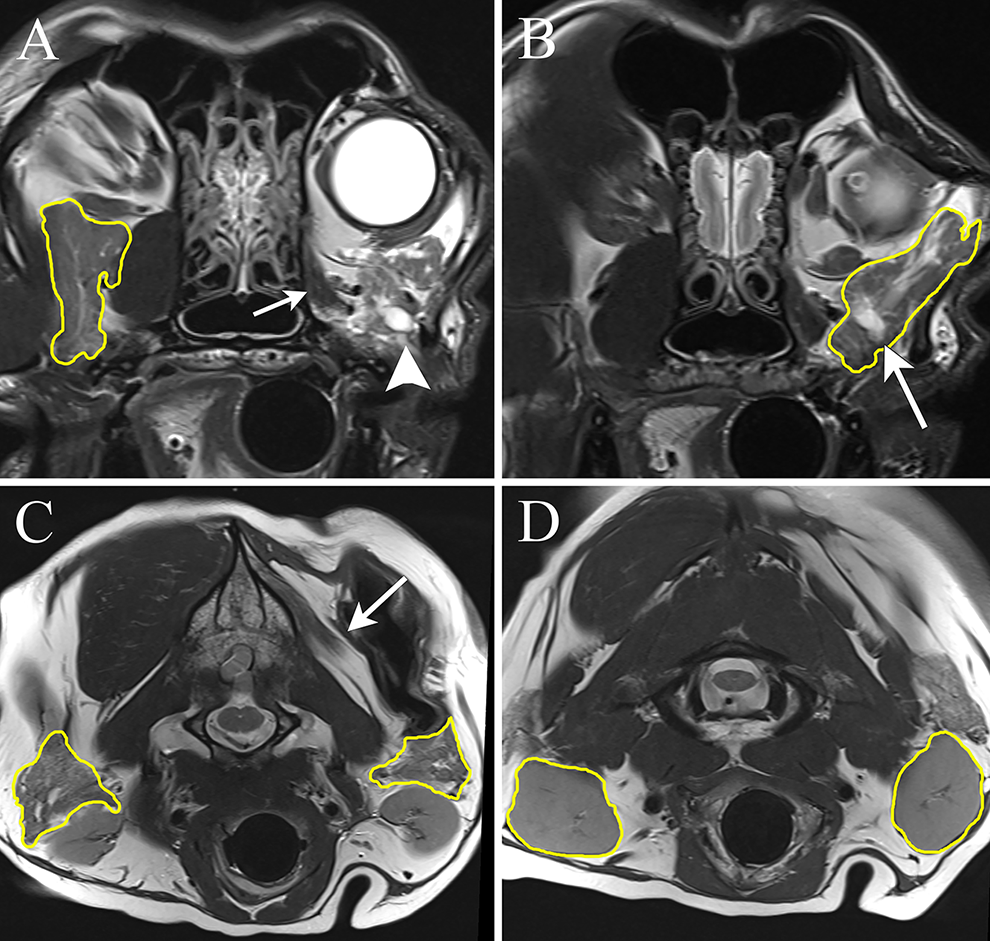
The transverse T2-weighted magnetic resonance imaging from a 14-year-old neutered male Husky-mix (dog 10) with left-sided trigeminal nerve dysfunction. In all the panels, the salivary glands are outlined in yellow as a representation of the largest cross-section area measurements. (A) The right zygomatic salivary gland has a homogenous signal intensity with a centrally positioned, dorsoventrally oriented linear T2-hyperintense structure that likely represents a duct within the gland. The left eye is visible due to enophthalmos on the side of the affected nerve. Just ventral to the left eye, the rostral aspect of the left zygomatic gland is visible (arrowhead). Note the atrophy of the pterygoid muscle (small arrow). (B) In comparison to the right side, the left zygomatic gland is hyperintense with more conspicuous lobulations (arrow). The largest cross-sectional area of the left zygomatic salivary gland was observed in a different image. The ducts within the salivary gland also appear dilated. C, The cross-sectional area of the left parotid salivary gland is smaller than that on the right side. Note the atrophy of the temporalis muscle (arrow) D, The largest cross-sectional area for the mandibular salivary glands appear similarly sized. Note the atrophy of the temporalis muscle.
In 2 dogs (dog 2 and dog 8), gross and histological evaluation of the salivary glands was performed. The weight of the unpreserved salivary glands from the side of the head with trigeminal or mandibular nerve dysfunction and unaffected side of the head were recorded. Due to the small size and close anatomic relationship, the mandibular and sublingual glands were dissected and weighed as one structure in dog 8. In dog 8, all the salivary glands from the affected side weighed less than the same glands on the unaffected side. On the affected side, the weight of the glands were as follows: parotid = 1.3 g, zygomatic = 2.9 g, and mandibular/sublingual = 2.4 g, while the weight of the glands on the unaffected side were: parotid = 1.5 g, zygomatic = 3.3 g, and mandibular/sublingual = 3.2 g. In this dog, the MRI included the salivary glands, and therefore largest cross-sectional area measurements were obtained. The largest cross-sectional area measurements of the glands from the side of the head with trigeminal or mandibular nerve dysfunction were smaller than those of the unaffected side. In dog 2, only the zygomatic gland on the affected side weighed less (6 g) than the zygomatic gland on the unaffected side (8 g). The mandibular and sublingual glands weighed the same on both sides. The mandibular weighed 14 g and sublingual glands weighed 2 g. The parotid gland on the affected side weighed more (18 g) than that on the unaffected side (12 g). The gross and microscopic findings of the formalin- preserved glands from the affected and unaffected sides of the head were similar. Briefly, the glands from both sides had similar staining characteristics of the glandular epithelia, development of acini, and amount of connective tissue dissecting between lobules of the glands.
A definitive diagnosis of a nerve sheath neoplasm of the trigeminal nerve was established in 2 dogs based on microscopic evaluation of the affected trigeminal and mandibular nerves. In the remaining dogs, a presumptive diagnosis of a nerve sheath neoplasm or neuritis was based on previously established MRI criteria. 12
Discussion
Saliva plays an important role in maintaining oral health through lubrication, maintenance of pH, prevention of caries as well as in food digestion. An excess of saliva or hypersalivation can cause drooling and sialorrhea, which can lead to disturbance in hygiene and social stigma in human beings. Conversely, inadequate salivation, hyposalivation or xerostomia, can lead to a decline in oral health, profound discomfort, and difficulty with mastication and swallowing in human beings. 13 In dogs, excess or inadequate salivation is rarely considered a primary disease process. Despite this, normal saliva production likely plays an important role in oral health of dogs. Poor oral health in dogs can have systemic consequences. 7,8,10 The first step in establishing the role of salivation in the maintenance of oral health is an understanding of the anatomy and physiology of the salivary glands.
There are 4 major salivary glands in the dog that contribute to oral secretions. These include the parotid, zygomatic, mandibular, and sublingual glands. 1 Each of these salivary glands has a distinct innervation and secretion profile. The salivary glands receive parasympathetic and sympathetic innervations. Salivary flow is primarily evoked by the parasympathetic nervous system mediated by acetylcholine. 14
Parasympathetic innervation to the salivary glands is provided through the parasympathetic nuclei for the facial and glossopharyngeal nerves in the medulla (Table 2). The parasympathetic preganglionic cell bodies for the parotid and zygomatic glands are located in the parasympathetic nucleus of the glossopharyngeal nerve. 1 Preganglionic axons course through the glossopharyngeal nerve to synapse at the otic ganglion. 1 The postganglionic axons course through the auriculotemporal nerve, a branch of the mandibular nerve, to innervate the parotid and zygomatic glands. 1 The parasympathetic preganglionic cell bodies for the mandibular and sublingual glands are located in the parasympathetic nucleus of the facial nerve. 1 Preganglionic axons course through the facial nerve where they project rostrally through the tympanic cavity to join the lingual nerve, a branch of the mandibular nerve, at the level of the oval foramen. 1 Preganglionic axons then course in the lingual nerve to synapse in the mandibular ganglion and sublingual ganglion to ultimately innervate the mandibular and sublingual glands, respectively. 1
Parasympathetic Innervation to the Major Salivary Glands.a
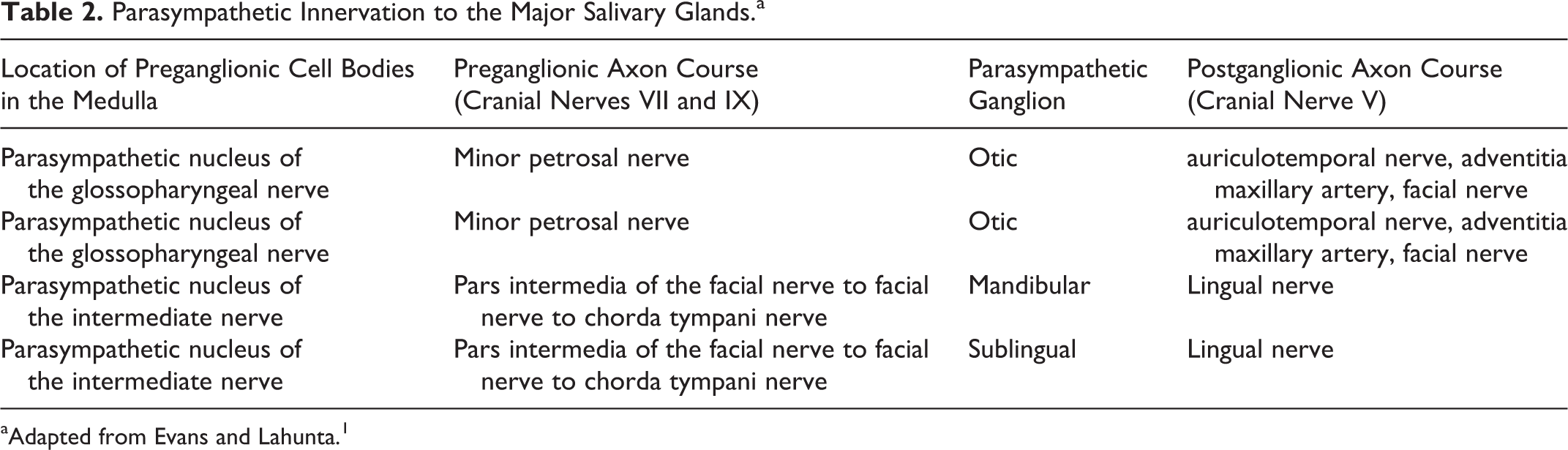
aAdapted from Evans and Lahunta. 1
The salivary glands also receive sympathetic input from preganglionic sympathetic cell bodies located in the lateral horn of the gray column of the first few thoracic spinal cord segments. The respective spinal nerves exit the vertebral column, course via the segmental ramus communicans, and ascend the vagosympathetic trunk. 15,16 The preganglionic axons synapse in the cranial cervical ganglion and have their postganglionic sympathetic axons travel along the arteries that supply each salivary gland. 1
Salivary flow is largely mediated by the parasympathetic nervous system through stimulation of the M1 and M3 muscarinic cholinergic receptors that not only increases the volume of saliva produced but also results in an increased viscosity through exocytosis of stored protein granules. 14,17 Despite an increase in total protein content with parasympathetic stimulation, the resultant saliva has a lower protein concentration than saliva produced under basal (resting) conditions due to the larger volume of fluid that is secreted. 18
Stimulation of the sympathetic nervous system, mediated by noradrenaline, can further modulate the composition of saliva by increasing degranulation from salivary cells. 19 Additionally, sympathetic stimulation of the blood vessels decreases blood flow through the salivary glands. 19,20
In addition to the autonomic nervous system, nonadrenergic and noncholinergic neurotransmitters such as vasoactive intestinal peptide and substance P also result in increased saliva production. 20,21 Referred to as secretory nerves, this input courses along axons associated with the maxillary artery. 22 Ultimately, disturbance anywhere along the pathway from the brain stem nuclei, the nerves providing autonomic and nonautonomic input, the neurotransmitters to the salivary glands, and ducts can alter the production and composition of saliva.
In all the dogs presented here, an accumulation of abnormal saliva was noted in the caudal aspect of the oral cavity in proximity to the opening of the ducts of the parotid and zygomatic glands on the side ipsilateral to a loss of function of the trigeminal nerve and/or it branches. The lack of concurrent clinical examination findings involving the facial nerve or imaging evidence of neurological disease apart from those related to the trigeminal nerve or the mandibular nerve combined with the location of the saliva accumulation suggests denervation of the parotid and/or zygomatic glands. The mechanism by which the parasympathetic fibers are damaged in disease processes that involve the trigeminal nerve or its branches is unknown. While speculative, in dogs with trigeminal nerve neoplasms, invasion or compression of the parasympathetic axons coursing with the trigeminal nerve or its branches may result in loss of parasympathetic innervation. Similarly, in dogs with trigeminal neuritis, inflammation involving the trigeminal nerve or its branches may also affect the parasympathetic axons.
Experimentally, avulsion of the auriculotemporal nerve can reduce acetylcholine synthesis and result in a 95% reduction in saliva produced by the parotid gland. 20,22,23 Although the volume of saliva was not determined in the dogs in this report, the abnormal visual characteristics of the accumulated saliva suggest a change in saliva composition from its normal watery consistency to a more viscous state. While the physical appearance of saliva is rarely reported in experimental models, loss of parasympathetic innervation not only impacts the volume of saliva produced but also impacts the protein content as well as other constituents that likely change physical and visual characteristics of saliva. 24 -26 Interestingly, avulsion or sectioning of the auriculotemporal nerve, which affects postganglionic axons, can also result in marked salivary flow for several days. 19,27 Referred to as degeneration secretion, it is likely related to leakage of acetylcholine from the postganglionic nerve endings. 27 Degenerative secretion is the consequence of an acute lesion; therefore, it likely does not explain the accumulation of abnormal saliva in all the dogs reported here, given their chronic history of neurological dysfunction.
It is also possible that a loss of the sensory function involving the trigeminal nerve may have influenced salivation. The mandibular nerve specifically provides sensory function to the cheek, tongue, teeth of the mandible, the lower lip, and the skin of the head. 1 In human beings, anesthetic ointment applied to the oral mucosa reduces saliva flow secondary to decreased activity of the mechanoreceptors involved in the parotid–masticatory salivary reflex. 28 While it was difficult to discern sensory function to the oral cavity or skin of the mandible, loss of sensation in the territory of the maxillary or ophthalmic nerves in 2 dogs was noted. In these dogs, more widespread sensory loss may have been present.
Another consequence to denervation of the salivary glands in human beings is atrophy of the gland. 13 Atrophy is a consequence of denervation atrophy as well as non-use (disuse) atrophy secondary to a loss of reflex salivation involving trigeminal reflexes. 13 Experimentally, injury to the nerve supplying parasympathetic innervation to the parotid gland results in atrophy of the gland. 29 Specifically, experimental injury at the level of the oval foramen that involves the otic ganglion results in parotid gland atrophy, whereas partial injury of the otic ganglion, which spares some parasympathetic innervation, may not result in atrophy of the parotid gland. 20,23 Significant differences in the largest cross-sectional area measurements were observed on the MRI studies in a cohort of dogs in this report. In support of denervation atrophy, there was also a significant increase in the mean signal intensity of the glands on the side ipsilateral to the trigeminal and/or mandibular nerve dysfunction. Fatty infiltration secondary to denervation occurs with denervation of the salivary glands. 29 An increase in fat composition would be expected to increase the mean signal intensity on a T2W image. Despite this, subjective differences in the microscopic evaluation of the salivary glands were not noted.
The present study has limitations. A definitive etiologic diagnosis was not established in all dogs. Different disease processes may not affect the mandibular nerve as far distal as the otic ganglion or auriculotemporal and lingual nerves and, therefore, may not involve postganglionic axons to the salivary gland. Moreover, not all dogs had an MRI performed or were necropsied to best assess the distal extent and degree of involvement of the trigeminal and mandibular nerves. Specifically, the otic ganglion or the parasympathetic postganglionic axons in the auriculotemporal and lingual nerves were not evaluated. Magnetic resonance imaging is unlikely to resolve the otic ganglion and nerves conducting postganglionic axons. Likewise, histological assessment of the otic ganglion and nerves conducting postganglionic axons may not predict functional deficits. Additionally, microscopic evaluation may not enable determination of functional deficits such as the ability to initiate and conduct an action potential at the level of the ganglion or in the postganglionic parasympathetic axons, respectively, with compression or infiltration of the tissue by neoplastic or inflammatory cells.
Likewise, the largest cross-sectional area measurement may not accurately reflect the atrophy of the gland. Moreover, difference in imaging parameters affected spatial resolution, which likely impacted the accuracy of the tracing of the gland. Volumetric analysis would likely provide more accurate assessment of gland size. Finally, the mean signal intensity may not relate to gross or histological changes in the gland. Despite the significant difference observed in the mean signal intensity in the ROI, fatty infiltration was not observed histologically in the two dogs that underwent necropsy.
Conclusion
Dogs with dysfunction of the trigeminal or mandibular nerve accumulate abnormal appearing saliva in the caudodorsal aspect of the oral cavity suggestive of hypofunction of the parotid and/or zygomatic glands. Xerostomia negatively impacts oral health and leads to dental caries in human beings. 30 Although the possible impact that the abnormal saliva had on oral health was not investigated, the severity of the dental calculus present on the side ipsilateral to the dysfunction of the trigeminal nerve in one dog implies a relationship. Future studies should not only investigate salivary gland structure and its function through analysis of the volume and composition of saliva but also the potential impact xerostomia has on the oral and systemic health in dogs with dysfunction of the trigeminal nerve and/or it branches.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Materials
1.5-Tesla, GE Brivo MR 355 Inspire; GE Healthcare, Princeton, New Jersey, with multichannel head coil. 3.0-Tesla MRI; Siemens, Skyra, Malvern, Pennsylvania. Omniscan, Amersham Health, Princeton, New Jersey. Magnevist; Bayer Healthcare Pharmaceu-ticals, Wayne, New Jersey. eFilm, Merge Healthcare, Milwaukee, Wisconsin. NilRead Enterprise Viewer, Westlake, Ohio. SAS version 9.3, Cary, North Carolina.