
Abstract
Experiences of adversity in the early years of life alter the developing brain. However, evidence documenting this relationship often focuses on severe stressors and relies on peripheral measures of neurobiological functioning during infancy. In the present study, we employed functional MRI during natural sleep to examine associations between a more moderate environmental stressor (nonphysical interparental conflict) and 6- to 12-month-old infants’ neural processing of emotional tone of voice. The primary question was whether interparental conflict experienced by infants is associated with neural responses to emotional tone of voice, particularly very angry speech. Results indicated that maternal report of higher interparental conflict was associated with infants’ greater neural responses to very angry relative to neutral speech across several brain regions implicated in emotion and stress reactivity and regulation (including rostral anterior cingulate cortex, caudate, thalamus, and hypothalamus). These findings suggest that even moderate environmental stress may be associated with brain functioning during infancy.
Prominent ideas about how the environment shapes development rest on an understanding that brain plasticity during the first years of life confers vulnerability for key neural systems involved in stress and emotion-related functioning. The consequences of early life stress have been investigated by examining its impact on these systems (Sánchez, Ladd, & Plotsky, 2001). Research with infants and young children has employed peripheral indicators of neuroendocrine functioning (e.g., cortisol; Loman & Gunnar, 2010) and direct measures of brain electroencephalographic activity (Nelson & McCleery, 2008) to increase understanding of how early adversity affects neurobehavioral development.
The high spatial resolution of functional MRI (fMRI), commonly used with older children and adults, has facilitated precise identification of neural networks linking early adversity with subsequent socioemotional functioning. Consistent with animal models examining the consequences of early adversity (Sánchez et al., 2001), this work reveals the involvement of brain regions tied to initiation and regulation of the hypothalamic-pituitary-adrenal (HPA) axis stress response, including limbic (Tottenham et al., 2011) and medial prefrontal regions (Treadway et al., 2009). However, the existing knowledge base in this area derives from fMRI research involving older children and adults. This makes it difficult to distinguish effects of early stress from subsequent processes of recovery or development of psychopathology. Moreover, a predominant focus on severe stressors, such as institutional rearing or maltreatment (Hart & Rubia, 2012), leaves a gap in the empirical literature regarding effects of more moderate early adversity.
Nonphysical interparental conflict is a more moderate source of early adversity that nevertheless appears to be associated with alterations in stress hormones, behavioral symptoms, and socioemotional problems during childhood (Cummings & Davies, 2010; Davies, Sturge-Apple, Cicchetti, & Cummings, 2007). Although more sparse than research with older children and adults, research with infants indicates that interparental conflict is associated with differences in physiological and behavioral indices of emotional reactivity and regulation as early as 6 months of age (Crockenberg, Leerkes, & Lekka, 2007; Moore, 2010). Interparental conflict may have an impact on early emotional development through decreases in sensitive caregiving (Krishnakumar & Buehler, 2000), as well as direct exposure to aggressive interactions between caregivers (Crockenberg et al., 2007). Basic research suggests that 5-month-old infants discriminate between other people’s different emotional states, with expressions of anger eliciting greater attention and arousal than happy or neutral expressions (Balaban, 1995; Grossmann, Oberecker, Koch, & Friederici, 2010; Grossmann, Striano, & Friederici, 2005). Additionally, Moore (2009) showed that infants who witnessed vocal anger toward their mother demonstrated altered parasympathetic nervous system responses to an immediately subsequent stressful interaction with their mother. Specifically, they showed increased withdrawal of vagal tone and decreased recovery, both of which are indicative of greater physiological reactivity, after this brief exposure to anger (Moore, 2009).
Early exposure to interparental conflict may also increase risk for later emotional and psychological problems. In 6-month-old infants, higher levels of interparental conflict are associated with lower baseline vagal tone (C. Porter, Wouden-Miller, Silva, & Porter, 2003) and greater withdrawal of vagal tone during a stressful interaction-and-recovery period (Moore, 2010), indicative of lower parasympathetic tone and greater stress reactivity, respectively. Variation in vagal reactivity acts as a moderator of risk for school-age children exposed to conflict (El-Sheikh et al., 2009; El-Sheikh & Whitson, 2006). Despite the implication that some aspects of nervous-system functioning may be shaped by family conflict during infancy, and subsequently increase risk for school-age children, the ties between early exposure and subsequent vulnerability remain poorly understood. The autonomic and behavioral measures utilized to date represent outputs from multiple neural networks. Candidate neural networks linking early adversity with subsequent risk for psychopathology have not yet been identified.
Recent work demonstrates the feasibility of conducting fMRI research with infants during natural sleep (Redcay, Kennedy, & Courchesne, 2007), which allows for examination of specific neural regions and networks during the first years of life. This work also draws attention to the sensitivity of infants to environmental stimuli during sleep by documenting distinct patterns of neural activation depending on properties of speech (Dehaene-Lambertz, Dehaene, & Hertz-Pannier, 2002; Redcay, Haist, & Courchesne, 2008) and emotional tone (Blasi et al., 2011). The present study builds on these methodological advances to characterize infants’ neural responses to emotional stimuli in the context of varying levels of interparental conflict.
Method
Participants
Families were recruited through flyers posted at local human-services agencies and advertisements on the local Craigslist.org Web site. Twenty-four infants (8 females, 16 males) aged 6 to 12 months (M = 8.33, SD = 1.90) completed an auditory fMRI paradigm during natural sleep; 20 infants had usable fMRI data. Infants had no known neurological disorders and lived with both biological parents. Exclusion criteria included referrals or investigations by a public child-protective-services agency. To obtain sufficient range in the sample, we assessed interparental conflict during screening using the Problem-Solving Communication subscale from the Marital Satisfaction Inventory, Revised (Snyder, 1997) and selected families based on established norms for distressed versus nondistressed couples (see the Supplemental Material available online for further information about the participants).
Interparental-conflict measures
Mothers rated nonphysical interparental-conflict levels since the birth of the child on the Psychological Aggression subscale of the Revised Conflicts Tactics Scale (Straus, Hamby, Boney-McCoy, & Sugarman, 1996) and the O’Leary-Porter Scale (B. Porter & O’Leary, 1980). The measures were highly reliable (α = .936 and .823, respectively) and correlated, r(22) = .744, p < .001, which allowed for creation of an average composite score of maternal report of interparental conflict (see the Supplemental Material for more information about the administration of these measures and the creation of the composite score).
Auditory stimuli
Auditory stimuli consisted of previously validated nonsense sentences spoken in very angry, mildly angry, happy, and neutral tones of voice by a male adult (Pell, Paulmann, Dara, Alasseri, & Kotz, 2009). Nonsense sentences possessed phonological and grammatical properties of English, but content words were replaced by semantically meaningless sound strings (see the Supplemental Material for more information).
fMRI data acquisition
Infants came in for scanning at their regular bedtime. Neuroimaging data were collected on a Siemens Allegra 3.0T scanner with a phased-array coil. Consistent with previous neuroimaging research using auditory stimuli with sleeping toddlers (Redcay et al., 2007), the paradigm consisted of 20-s blocks separated by 15-s rest periods. Blocks for each emotion condition (very angry, mildly angry, happy, and neutral) were presented four times per run in a semicounterbalanced design based on a Williams’s Latin square. T2-weighted echo-planar functional scans (9 min, 28 s; 284 whole brain volumes) were acquired during presentation of auditory stimuli. Prospective acquisition correction was applied to adjust slice position and orientation, as well as to regrid residual volume-to-volume motion in real time during data acquisition for the purpose of reducing motion-induced effects (Thesen, Heid, Mueller, & Schad, 2000). See the Supplemental Material for more information on the scanning protocol and data-acquisition process.
fMRI data analysis
Neuroimaging data were converted to Neuroimaging Informatics Technology Initiative data format using the MRIConvert program (http://lcni.uoregon.edu/~jolinda/MRIConvert/). Brain images were extracted using the Brain Extraction Tool from the FMRIB Software Library (Beckmann et al., 2006; S. M. Smith, 2002; S. Smith, Bannister, Beckmann, & Brady, 2001) and the Brain Surface Extraction tool from BrainSuite09 (Sandor & Leahy, 1997; Shattuck, Sandor-Leahy, Schaper, Rottenberg, & Leahy, 2001). All other preprocessing steps, including realignment, registration, normalization, and smoothing with a 6-mm full-width half-maximum kernel, were accomplished using Statistical Parametric Mapping (SPM) software (SPM8; Wellcome Department of Cognitive Neurology, London, England). Images were normalized to a standard template for the 8- to 11-month age range from the MRI Study of Normal Brain Development (Fonov et al., 2011; Fonov, Evans, Mckinstry, Almli, & Collins, 2009). Images with severe motion artifacts (greater than 2 mm of motion or evidencing visual signs of motion artifacts) were removed from runs, which resulted in less than 2 mm of motion per run (maximum = 1.07 mm). At least three (out of four) blocks of each condition were retained from each run.
At the individual-subject level, fixed-effects contrasts were computed to examine neural activation during presentation of each condition (very angry, mildly angry, happy, and neutral) versus rest, as well as the specific contrast of the very angry condition relative to the neutral condition. Motion parameters in six directions were included as regressors of no interest. Functional runs for which the contrast of all auditory conditions to rest did not evidence auditory-cortex activation at a relaxed threshold (p < .05, uncorrected) were excluded from analyses because it was not possible to ascertain whether basic sensory processing of stimuli occurred. Four of the 24 infants did not have at least one functional run for which clear auditory activation was detected and were thus excluded from further analyses. The resulting contrast images were entered into whole-brain random-effects group analyses. We report only results that exceeded a threshold of p < .05, family-wise-error (FWE) corrected for multiple comparisons across the whole brain (specifically, p < .05 and 75 contiguous voxels, as determined by the NeuroElf AlphaSim toolbox, http://neuroelf.net/). Regions of activation were identified based on anatomical landmarks, although infant template and Montreal Neurological Institute template coordinates are provided in the figure and tables for reference.
Results
Effect of interparental conflict on processing very angry tone of voice
The primary research question focused on the extent to which the composite interparental-conflict score was associated with infants’ neural responses to very angry auditory stimuli relative to neutral auditory stimuli. A whole-brain regression with interparental-conflict score as the independent variable and neural activity during very angry relative to neutral tone of voice as the dependent variable revealed a significant cluster in rostral anterior cingulate cortex (ACC) as well as a subcortical cluster encompassing parts of the caudate, thalamus, and hypothalamus (Table 1). Specifically, higher levels of interparental conflict were associated with greater activation in these regions during presentation of very angry compared with neutral speech (Figs. 1a and 1b). To depict this association graphically, we extracted mean parameter estimates of activity (averaged across all voxels in each cluster) for each participant from both the rostral ACC and subcortical cluster during the very-angry-versus- neutral contrast using the MarsBaR region-of-interest toolbox for SPM (Brett, Anton, Valabregue, & Poline, 2002). These mean parameter estimates for each participant in each cluster were then plotted as a function of conflict score. The graphs in Figures 1c and 1d do not show the results of an additional statistical analysis; rather, they illustrate the positive association between conflict and activation of these regions to very angry relative to neutral speech that was demonstrated statistically with the fMRI analyses. Results remained consistent when we controlled for variation in infant age. These results were specific to the very-angry-relative-to-neutral contrast. Exploratory analysis of the association between interparental conflict and neural processing of happy speech are presented in the Supplemental Material.
Results of the Whole Brain Regression: Positive Association Between Interparental Conflict and Brain Activation in Response to Very Angry Relative to Neutral Tone of Voice

Note: The table reports only results that exceeded a threshold of p < .05, family-wise-error corrected for multiple comparisons across the whole brain (specifically, p < .05 and 75 contiguous voxels). The number of voxels within each cluster is indicated by k. Coordinates without voxel numbers indicate submaxima within the preceding cluster. MNI = Montreal Neurological Institute.
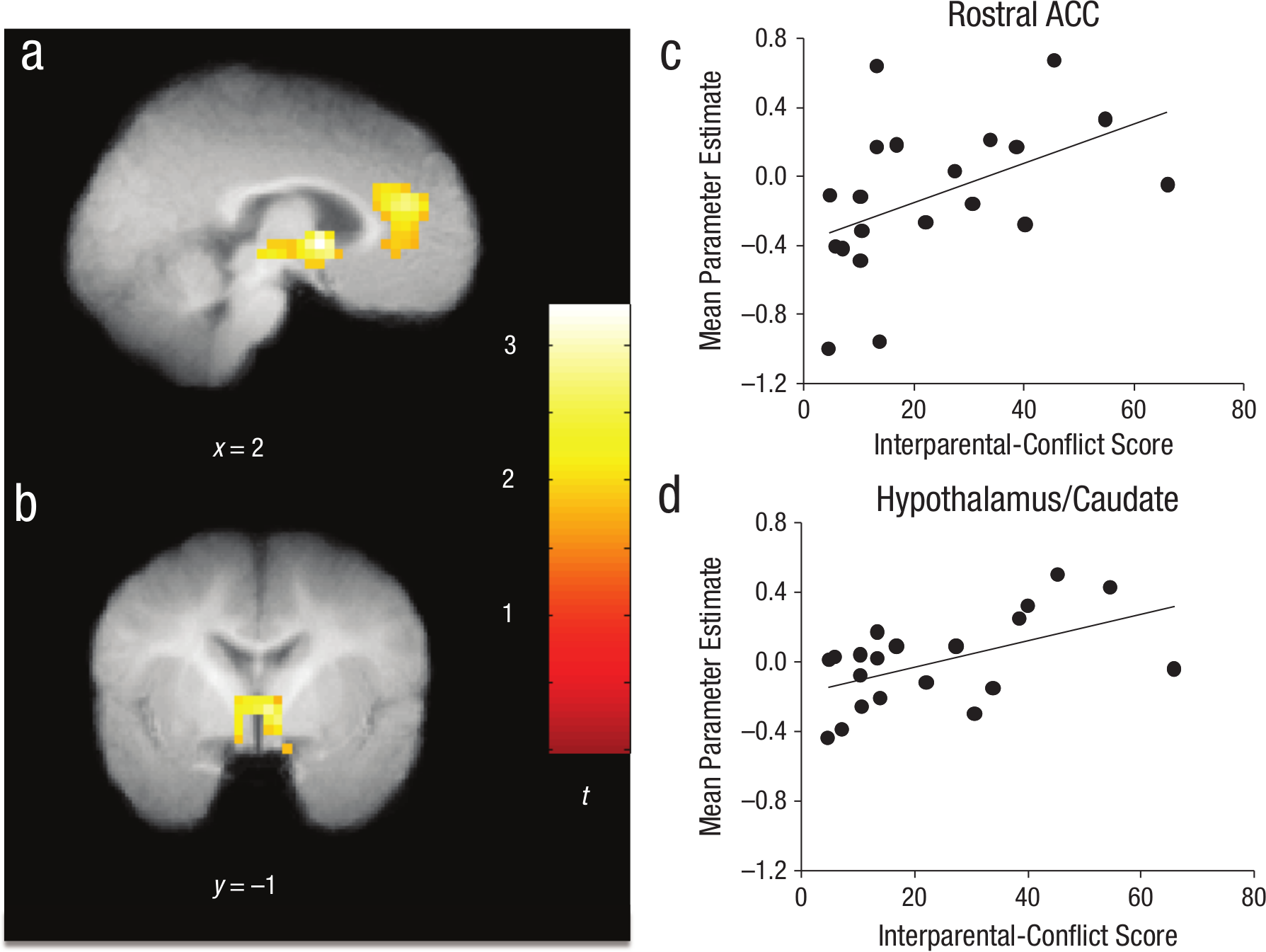
Association between interparental-conflict scores and neural reactivity to very angry speech relative to neutral speech. Activations that exceeded a threshold of p < .05, family-wise-error corrected for multiple comparisons across the whole brain (specifically, p < .05 and 75 contiguous voxels), are displayed on the group mean structural image. The brain image in (a) shows activation in the rostral anterior cingulate cortex (ACC; infant-atlas coordinates: x = 3, y = 29, z = 13; Montreal Neurological Institute coordinates: x = 4, y = 36, z = 17). The images in (a) and (b) show activation in a subcortical cluster including hypothalamus, caudate, and thalamus (infant-atlas coordinates: x = 3, y = −1, z = 1; Montreal Neurological Institute coordinates: x = 4, y = −1, z = 1), in which higher conflict scores predicted greater neural response to very angry than to neutral speech. The scatter plots (c and d; with best-fitting regression lines) reillustrate the association between conflict score and parameter estimates, separately for these two regions.
Effect of different emotional tones of voice
Because conflict was associated with neural responses to very angry speech, brain activation during presentation of each emotional tone of voice was examined after regressing out individual differences in conflict (Table 2). Direct comparison of activations in response to very angry relative to neutral stimuli did not reveal any clusters surviving FWE correction, although a cluster in the left temporal pole was just below this extent threshold. Comparison of activations in response to happy relative to neutral stimuli revealed significant areas in the left dorsolateral prefrontal cortex, putamen, and medial temporal and occipital cortices.
Activation to Emotion Stimuli After Regressing Out Interparental Conflict
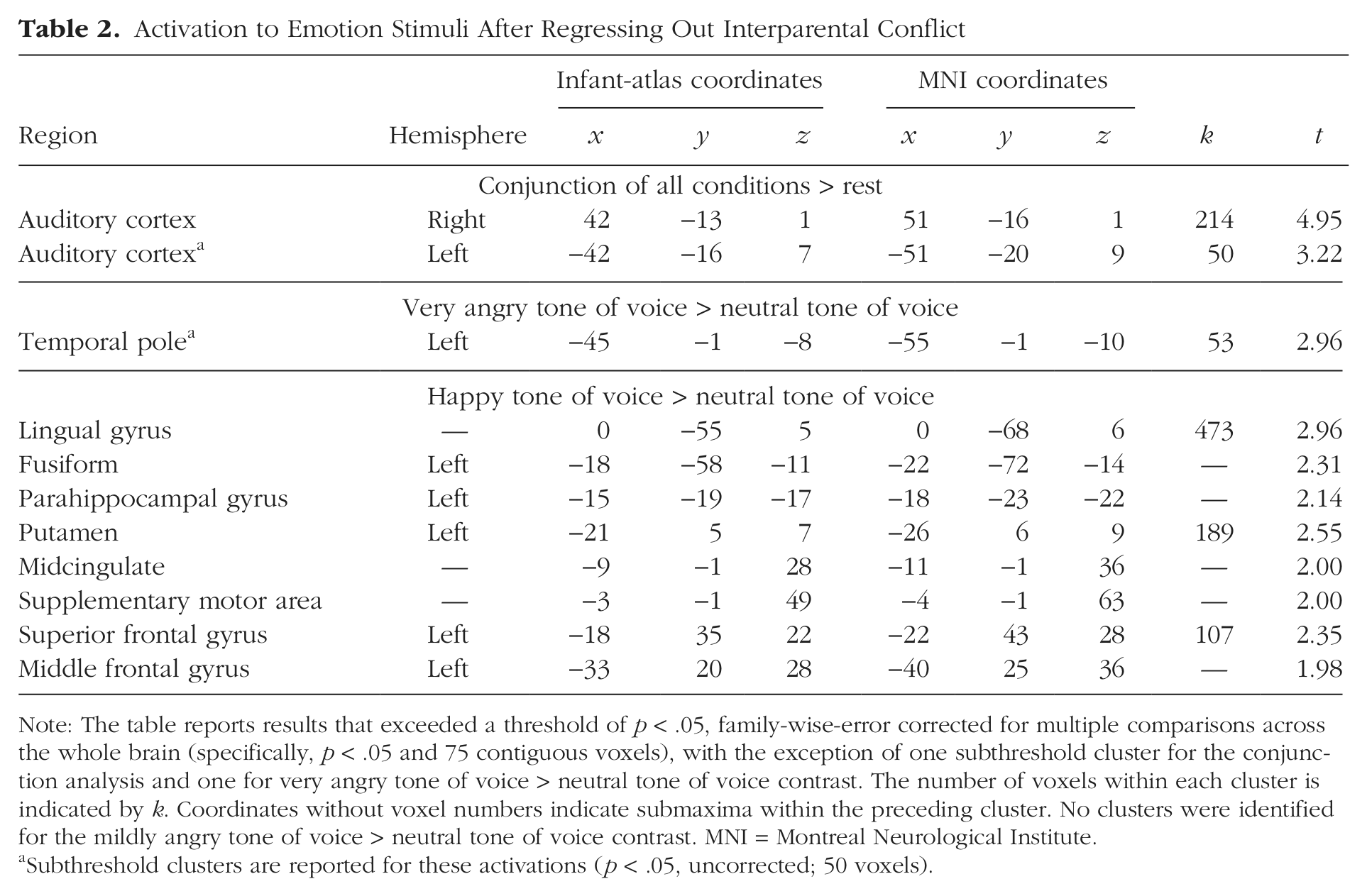
Note: The table reports results that exceeded a threshold of p < .05, family-wise-error corrected for multiple comparisons across the whole brain (specifically, p < .05 and 75 contiguous voxels), with the exception of one subthreshold cluster for the conjunction analysis and one for very angry tone of voice > neutral tone of voice contrast. The number of voxels within each cluster is indicated by k. Coordinates without voxel numbers indicate submaxima within the preceding cluster. No clusters were identified for the mildly angry tone of voice > neutral tone of voice contrast. MNI = Montreal Neurological Institute.
Subthreshold clusters are reported for these activations (p < .05, uncorrected; 50 voxels).
Discussion
Although unusually adverse experiences such as institutional rearing or maltreatment are known to affect development of key neural networks, the present study suggests potential effects of a more moderate environmental stressor, nonphysical interparental conflict. By taking advantage of recent methodological advances that allow for investigation of neural functioning during infancy with the high spatial resolution afforded by fMRI (Blasi et al., 2011; Dehaene-Lambertz et al., 2010), this study provides novel evidence of associations between interparental conflict and patterns of infant brain functioning elicited by processing emotional speech during natural sleep.
Higher levels of interparental conflict were associated with greater activation to very angry tone of voice in the rostral ACC and subcortical structures, including the hypothalamus. Although we cannot be certain about the meaning of the activation patterns in these brain regions, many studies indicate their involvement in emotion and stress processing and regulation (Kober et al., 2008). The rostral ACC is implicated in emotion processing and regulation in typical populations (Kober et al., 2008), and its functioning is frequently altered in stress-related disorders (Fonzo et al., 2010; Kim et al., 2008). Research also demonstrates associations between early adversity and decreased volume of the ACC for adults with (Treadway et al., 2009) and without symptoms of psychopathology (Cohen et al., 2006), although the developmental pathway through which these structural differences emerge remains unknown.
The hypothalamus initiates activity of the HPA axis. Activity of the HPA axis in response to psychosocial stress is controlled by limbic brain structures involved in emotion processing and memory, including the amygdala, hippocampus, and ACC (Pruessner et al., 2010; Ulrich-Lai & Herman, 2009). The hypothalamus is thus viewed as a key link between emotional input, neuroendocrine functioning, and stress reactivity (Kober et al., 2008). Extensive research has focused on alterations in the functioning of the HPA axis (as indexed by the hormone cortisol) as a result of early life stress, including more normative stressors, such as interparental conflict (Davies et al., 2007), and more extreme events, such as neglect and abuse (Bruce, Fisher, Pears, & Levine, 2009). Specific patterns of HPA-axis functioning have also repeatedly been associated with mood disorders in adolescence and adulthood (Lopez-Duran, Kovacs, & George, 2009; Parker, Schatzberg, & Lyons, 2003).
These findings also converge with extensive research using animal models, which points to the ACC and hypothalamus as part of neural networks that link early psychosocial adversity to subsequent difficulties with regulation of emotions and stress (Loman & Gunnar, 2010). However, this study is the first to document an association between an environmental stressor and the functioning of these specific brain regions during infancy. These regions were identified based on a whole-brain regression as opposed to a priori specification as regions of interest. This allowed for a more independent test of whether the findings in this study converge with existing knowledge about the role of these brain regions based on animal models and research with older children and adults (Hart & Rubia, 2012).
This study also provides novel evidence regarding infants’ neural processing of happy and angry emotional speech during sleep, regardless of the level of interparental conflict. The findings are broadly in line with those of a recent fMRI study indicating differentiation of sad relative to neutral vocalizations in sleeping 3- to 7-month-olds (Blasi et al., 2011), although this study did not find differentiation between happy and neutral stimuli. We may have been better able to observe the latter pattern because of differences in the age ranges sampled and the stimuli (nonsense speech vs. emotional vocalizations).
Limitations of the present study include the lack of observational assessment of interparental conflict and of a high-intensity positive-affect condition (e.g., very happy) to test whether the effects are specific to anger rather than high-intensity emotion more generally. Additionally, recruitment through Craigslist and human-services agencies may have skewed the sample toward individuals of lower socioeconomic status. We also were unable to monitor and control for sleep state, which is an important issue to be addressed in future work (see the Supplemental Material). Future research will also benefit from longitudinal investigations and inclusion of behavioral measures to assess whether changes in neural functioning mediate between exposure to environmental stress and socioemotional development.
Despite these limitations, the present findings indicate that during a period when infants are particularly vulnerable because of their complete dependence on caregivers and high levels of neural plasticity, moderate sources of environmental stress may be related to neural functioning in areas central to emotion and stress-related processes. Moreover, far from being oblivious to parents’ conflict, infants’ processing of stressor-relevant stimuli, such as angry tone of voice, may occur even during sleep.
Footnotes
Acknowledgements
Special thanks are due to Scott Watrous at the University of Oregon’s Lewis Center for NeuroImaging, Kyndal Howell at the Oregon Social Learning Center and the University of Oregon Stress Neurobiology and Prevention Laboratory, and Weili Lin and Kathy Wilber at the Biomedical Research Imaging Center, University of North Carolina School of Medicine.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
Support for this work was provided by the Center for Drug Abuse Prevention in the Child Welfare System (1-P30-DA023920); the Early Experience, Stress, and Neurobehavioral Development Center (1-P50-MH078105); a Ruth L. Kirschstein National Research Service Award (F31-10667639); and the Lewis Center for NeuroImaging at the University of Oregon.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.