
Abstract
The study aimed to evaluate the effectiveness of two cleaners (Tergazyme and PowerFoam Plus) and two sanitizers (Decon 7 and BioDestroy) against biofilms formed by a variety of bacterial species obtained from meat processing plants (n = 44). Biofilms were developed on polystyrene pegs at alternating temperatures of 8°C for 16 h, followed by 15°C for 8 h, over a duration of 6 days. Bacteria within biofilms were enumerated by plating, and biomass was quantified by crystal violet staining and measuring absorbance (A570). Thirty of the 44 meat plant bacterial strains formed measurable biofilms, of which 43% (n = 19) were weak biofilm formers, 9% (n = 4) were moderate biofilm formers, and 16% (n = 7) were strong biofilm formers. Of the two sanitizers, Decon 7 was the most effective at biofilm eradication (100%). The effectiveness of BioDestroy for biofilm removal was improved by a pretreatment step of PowerFoam Plus and Tergazyme. These findings show that pretreatment with cleaning products is important for improving the effectiveness of some sanitizers, and soaking equipment with cleaners prior to application of sanitizers may improve biofilm removal in meat processing environments.
Introduction
Biofilms in food processing environments present a significant challenge because of their ability to serve as reservoirs for foodborne pathogens and spoilage microorganisms. Cross-contamination can occur when bacteria disperse from biofilms and consequently contaminate handling surfaces and food products (Marouani-Gadri et al., 2009; Silagyi et al., 2009; Wang et al., 2020). As such, the food industry has a strong interest in controlling biofilms to produce safe food with a long shelf life. Despite daily cleaning and disinfection of meat processing plants, there is increasing evidence of food contamination as a result of biofilm formation (Møretrø et al., 2013; Yang et al., 2017, 2018).
Biofilm formation is the process by which microorganisms adhere to surfaces, produce extracellular polymeric substances (EPS), and form complex, structured communities (Donlan, 2002; Hall-Stoodley and Stoodley, 2009). Biofilm communities provide mechanical, chemical and biological protection, rendering them more resistant to environmental stresses including biocides (Van Houdt and Michiels, 2005; Wang et al., 2020). Furthermore, bacteria in biofilms can colonize new niches by various dispersal mechanisms, thereby becoming a perpetual source of contamination either by survival of regular cleaning and sanitation processes or by colonizing difficult to access areas (Møretrø et al., 2013; Visvalingam et al., 2019).
Biofilms typically consist of mixed communities of microorganisms which contribute to their structure and function (Donlan, 2002). Within meat processing plants, biofilms can include aerobic and lactic acid-producing spoilage bacteria, Enterobacteriaceae as well as pathogenic bacteria (Koti et al., 2024; Wang et al., 2018). Various microorganisms such as Escherichia coli, Carnobacterium spp., Raoultella spp. and Pseudomonas spp. have been isolated from beef-processing conveyor belts after cleaning and sanitation (Beauchamp et al., 2012; Ma et al., 2019; Rangel et al., 2005; Wang et al., 2018). Consequently, these bacteria can combine to form mixed biofilms, creating a protective shield against conventional cleaning and sanitation (Visvalingam et al., 2017).
A previous study by Wang et al. (2018) noted that, Powerfoam Plus, a cleaner containing potassium hydroxide and sodium hypochlorite did not consistently reduce bacteria on the surface of “cleaned” conveyor belts. It was hypothesized that organic matter on the belt surface reduced the oxidizing power of chlorine, the active agent in the product (Wang et al., 2018). This study aimed to evaluate whether applying Tergazyme, a concentrated anionic detergent with protease, prior to applying Powerfoam Plus could eliminate single species biofilms of bacteria commonly found on meat fabrication equipment. We also compared this procedure to the effectiveness of two hydrogen peroxide based, food industry disinfectants: Biodestroy and Decon 7.
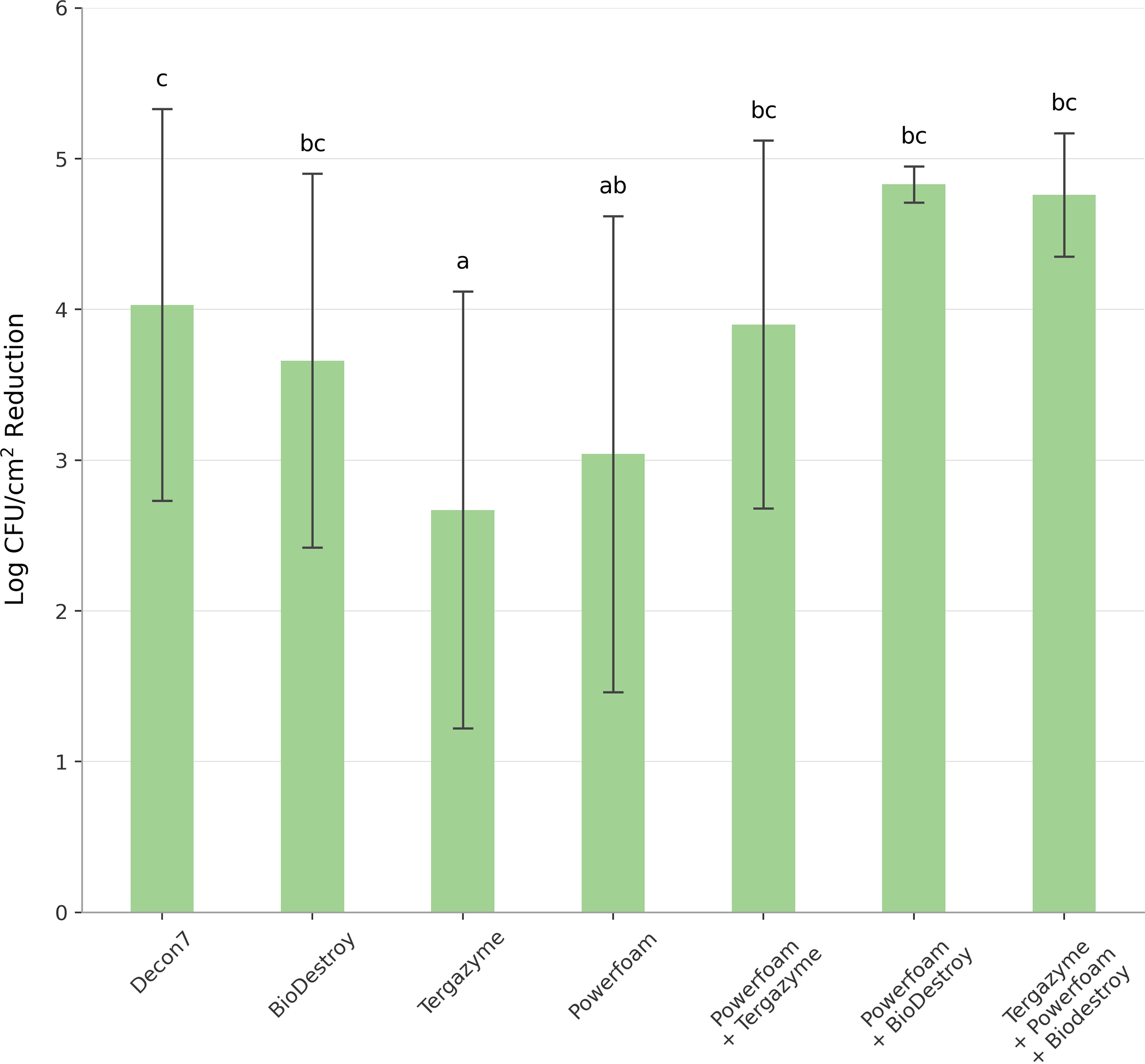
Overall biofilm reduction by treatment type. Mean values denoted by the same letters were not significantly different (p > 0.05).
Materials and Methods
Bacterial isolates
A total of 44 strains (Table 1), 40 of which were recovered from conveyor belts in a beef packing plant, before, during, and after routine sanitization, were included in this study (Wang et al., 2018). TStrains included 18 Gram-negative aerobes (GNA), 8 Gram-positive aerobes (GPA), 5 lactic acid bacteria (LAB), and 9 Enterobacteriaceae (EB) (Visvalingam et al., 2019; Wang et al., 2018). The other four meat plant isolates were generic E. coli (GEC; 136, CX10, AW1.7, CT02) of distinct genotypes (Visvalingam et al., 2017; Yang et al., 2021, 2023).
Meat Plant Bacterial Isolates Used in This Study

Strains selected for biocide experiments.
Culture conditions and inoculum preparation
Bacterial isolates were stored at −80°C in Tryptic Soy broth (Becton Dickinson, Mississauga, ON, Canada) containing 20% glycerol (ThermoFisher Scientific, Ottawa, ON, Canada), except for one Microbacterium strain, which was stored in Brain Heart Infusion (BHI) broth (Oxoid, Mississauga, ON, Canada) containing 20% glycerol. Cultures were maintained on Tryptic Soy Agar (TSA; Oxoid) at 4°C, and re-streaked onto fresh TSA every 2–4 weeks. Streaked plates were incubated at 25°C for 72 h. A single colony was inoculated into 5 mL of half-strength BHI and incubated at 25°C with shaking at 0.14 g for 72 h.
Stationary phase cultures grown in half-strength BHI were diluted 100-fold by adding 100 µL of culture to 9.9 mL sterile Meat Juice Medium (MJM; Gill, 1976) to obtain bacterial suspensions containing 107 colony-forming unit (CFU)/mL. The MJM was used to resemble the meat juices that are continuously wetting meat plant equipment surfaces and creating conditions conducive for bacterial biofilm formation (Giaouris et al., 2014).
Biofilm development and quantification
Biofilms were developed in 96-well round bottom microtiter plates (Nunc Immuno TSP lid; Fisher) fitted with a 96-peg polystyrene lid (Nunc; Fisher) (Ceri et al., 1999; Visvalingam et al., 2017). Suspended inoculum (160 µL) was added into duplicate wells for each of the 44 strains. Duplicate wells of uninoculated MJM were included as a blank control. Each plate was prepared in duplicate; one fitted with a 96-peg lid and the other with individual pegs (surface area 1.09 cm2) placed aseptically into each well. To mimic temperatures during beef processing, plates were incubated at 8°C for 16 h, followed by 15°C for 8 h, over 6 d (Yang et al., 2018). After 6 d, biofilms on peg-lids were quantified using the crystal violet method (Visvalingam et al., 2017). Peg-lids were washed for two 1 min periods in phosphate buffered saline (PBS; Hardy Diagnostics, Santa Maria, CA, USA), and then in a 0.1% aqueous (wt/vol) CV solution for 20 min, followed by two more PBS washes. Biofilms on pegs were de-stained in 80% (vol/vol) ethanol (Greenfield Globa, Brampton, ON, Canada) for 20 min. Absorbance (A570) of the CV solution was measured at 570 nm using a BioTek Synergy HT plate reader (Agilent, Mississauga, ON, Canada). Three independent experiments were performed and the mean A570 was calculated from six readings. Biofilm-forming ability was estimated using optical density cutoffs (ODc) as described by Yang et al. (2018).
Enumeration of bacteria in biofilms
Enumeration experiments were conducted using pegs aseptically removed from lids (Nunc Immuno TSP lid) prior to biofilm development (Visvalingam et al., 2017). Following 6 d of incubation, each peg was aseptically withdrawn and washed twice in 200 µL PBS for 1 min in a microtiter plate (Nunc, ThermoFisher Scientific, Ottawa, ON, Canada). Each washed peg was then placed in a 2 mL microcentrifuge tube (Eppendorf, Mississauga, ON, Canada) containing 300 mg of 0.5 mm sterile zirconia beads (ThermoFisher Scientific) and 1.2 mL of PBS. Each tube was vortexed for 2 min and the suspension was serially diluted 10-fold in PBS and plated on TSA. Plates were incubated at 25°C for 24–72 h. After incubation, plates containing 20–300 colonies were enumerated.
Preparation of cleaners/biocides
A hydrogen peroxide based disinfectant, BioDestroy (Sani Marc, Victoriaville, QC, Canada) and a multi-component disinfectant Decon7 (Decon7 Systems Inc., Coppell, TX, USA) were assessed for their ability to eradicate biofilms. BioDestroy was diluted, as per the manufacturer’s instructions, 1:100 in sterile water to 600 ppm. Decon7 consisted of: surfactant quaternary ammonium compounds (benzyl and dimethyl chlorides), an oxidizer (hydrogen peroxide) and an accelerator (diacetin). The 3 components of Decon7 were mixed 49:49:2 as per the manufacturers’ instructions. Chlorinated alkaline cleaner, PowerFoam Plus (Epsilon Chemicals Ltd., Edmonton, AB, Canada), and the protease detergent cleaner, Tergazyme (Alconox Inc., Jersey City, NJ, USA) were diluted in sterile water according to manufacturers’ instructions to obtain 2.5% (vol/vol) and 1% (10,000 ppm), respectively. All biocide solutions were used within 30 min after preparation.
Effect of single or sequential biocide treatment on biofilms
Biofilms were developed over 6 d in duplicate plates by incubating individual, unattached pegs in 160 µL of bacterial suspension under the dynamic temperature cycle. Pegs were withdrawn and washed twice with 200 µL sterile water and transferred to a microtiter plate well containing 180 µL of each of the individual biocide solutions (BioDestroy, Decon7, PowerFoam Plus, or Tergazyme). Plates were incubated at ambient temperature for 10 min, with the exception of Tergazyme which was incubated for 1 h. Pegs were then transferred to a 1.5 mL microcentrifuge tube (Eppendorf) containing 300 mg of 0.5 mm diameter sterile zirconia beads and 1.2 mL neutralizing buffer (Becton Dickinson) and vortexed for 2 min (Visvalingam et al., 2017). The suspension was serially diluted in PBS and enumerated on TSA plates after incubation at 25°C for 24–72 h. Further successive testing of biocides included: Tergazyme for 1 h followed by Powerfoam Plus for 10 min, Tergazyme for 1 h followed by Powerfoam Plus for 10 min followed by BioDestroy for 10 min, or Powerfoam Plus for 10 min followed by BioDestroy for 10 min.
Data analysis
Bacterial counts were log10 transformed prior to analysis and biofilm mass (A570) and log10 counts of bacteria in biofilms were analyzed using R version 4.5.2 (R Core Team, Vienna, Austria). Outliers were identified within each strain using the interquartile range (IQR) method where values below (Q1-1.5*QR or above Q3 + 1.5*QR were excluded. Biofilm forming ability was calculated, as described by Stepanović et al. (2007). A one-way analysis of variance (ANOVA) was used to analyze reductions in biofilm log counts. Strain differences within each biocide type were evaluated with strain as fixed factor while differences between biocide treatments were assed with biocide treatment as fixed factors, followed by post hoc group comparison using a Sidak test (p < 0.05).
Results
Biofilm formation
Thirty out of the 44 strains developed measurable biofilms (Table 2). The extent of biofilm formation among meat plant strains ranged from A570 values of 0.07 ± 0.02–2.02 ± 0.15. Measurable biofilms determined by biomass assessment were observed in 100%, 89%, 72%, 50%, and 20% of GEC, EB, GNA, GPA, and LAB, respectively. Of the biofilm forming strains, 43% (n = 19) were weak, 9% (n = 4) were moderate, and 16% (n = 7) were strong biofilm formers (Table 2). Biofilm growth as determined by enumeration ranged from 3.36 ± 1.72 to 8.25 ± 0.15 CFU/cm2. Biofilm growth of strong biofilm formers ranged from 6.46 ± 1.12 to 8.25 ± 0.15 CFU/cm2, and biofilm mass (A570) ranged from 0.29 ± 0.28 to 2.02 ± 0.15. For moderate biofilm formers, growth ranged from 5.48 ± 2.4 to 6.9 ± 0.23 CFU/cm2, and mass (A570) ranged from 0.15 ± 0.09 to 0.26 ± 0.28. Weak biofilm formers had growth ranging from 3.36 ± 1.53 to 7.23 ± 0.37 CFU/cm2, and mass (A570) ranging from 0.07 ± 0.02 to 0.14 ± 0.13. Non-biofilm formers had growth ranging from 1.7 ± 0.0 to 6.06 ± 0.43 CFU/cm2 and mass (A570) ranging from 0.06 ± 0.0 to 0.07 ± 0.02.
Biofilm Formation by Meat Plant Bacterial Isolates
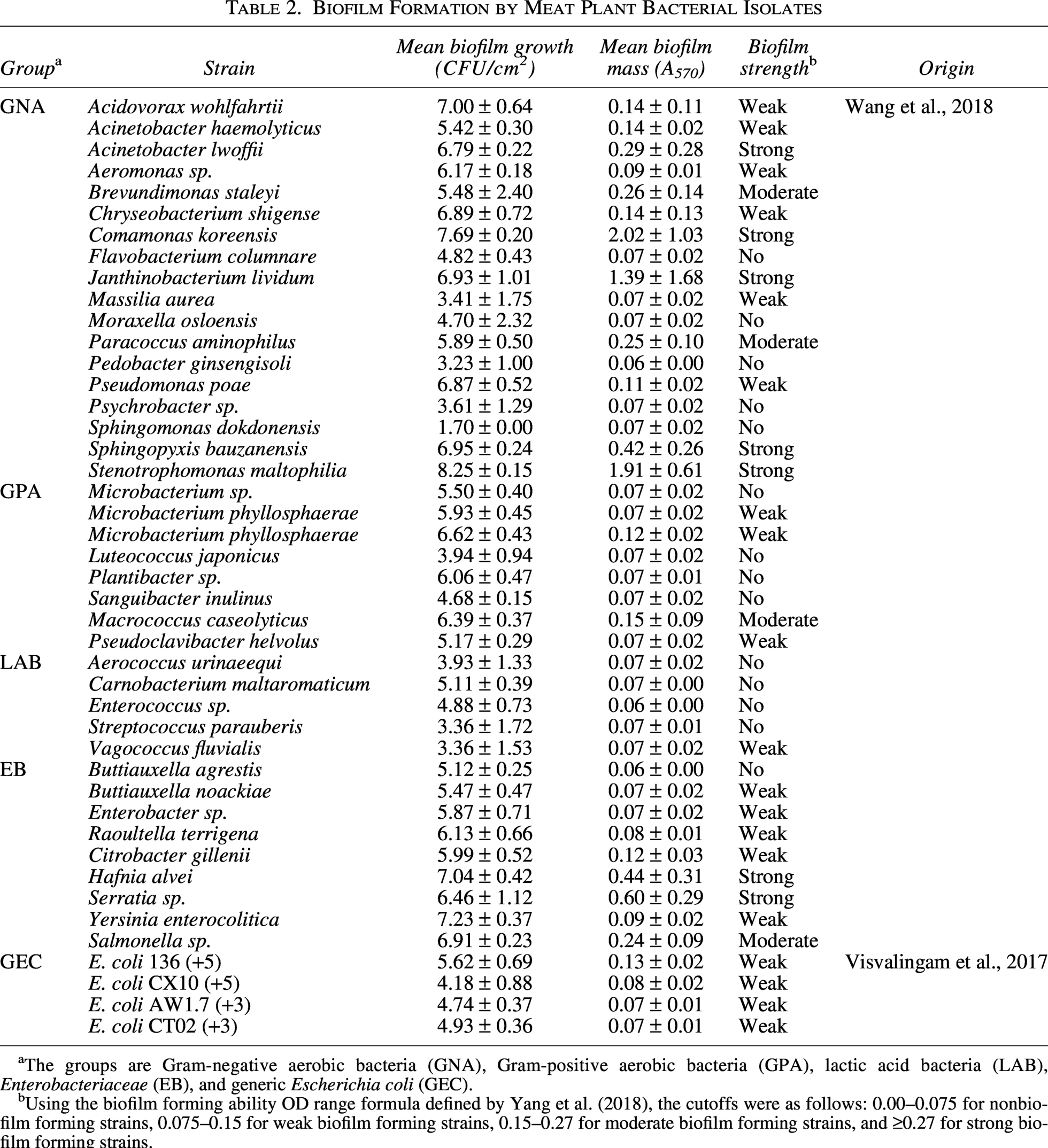
The groups are Gram-negative aerobic bacteria (GNA), Gram-positive aerobic bacteria (GPA), lactic acid bacteria (LAB), Enterobacteriaceae (EB), and generic Escherichia coli (GEC).
Using the biofilm forming ability OD range formula defined by Yang et al. (2018), the cutoffs were as follows: 0.00–0.075 for nonbiofilm forming strains, 0.075–0.15 for weak biofilm forming strains, 0.15–0.27 for moderate biofilm forming strains, and ≥0.27 for strong biofilm forming strains.
Of the 18 GNA strains, 28% (n = 5) did not form biofilms (Flavobacterium columnare, Moraxella osloensis, Pedobacter ginsengisoli, Psychrobacter sp., and Sphingomonas dokdonensis), whereas 33% (n = 6) were weak, 11% (n = 2) were moderate, and 28% (n = 5) were strong biofilm formers (Acinetobacter lwoffii, Comamonas koreensis, Janthinobacterium lividum, Sphingopyxis bauzanesis, and Stentrophomonas maltophilia). For GPA strains (n = 8), 50% (n = 4) did not develop biofilms, 37.5% (n = 3) and 12.5% (n = 1) were weak and moderate biofilm formers, respectively. The LAB strains (n = 5) were comprised of 80% (n = 4) nonbiofilm formers and 20% (n = 1) weak biofilm formers. Among the EB strains (n = 9), 56% (n = 5) were weak, 22% (n = 2) were strong (Hafnia alvei and Serratia sp.), 11% (n = 1) were moderate, and 11% (n = 1) did not form biofilms. The GEC strains (n = 4) were all weak biofilm formers.
Effect of biocides on bacterial biofilms
Biocides differed in their ability to reduce biofilms, with Tergazyme and Powerfoam being the least effective (Figure 1). Following treatment of biofilms with Decon 7, none (0/13) of the strains were cultivable (Table 3). After treatment with BioDestroy, PowerFoam Plus, or Tergazyme, 23% (3/13), 46% (6/13), and 54% (7/13), respectively, of the strains were recovered (Table 3). When Tergazyme was followed by PowerFoam Plus, the number of cultivable strains was reduced to 15% (2/13), with the two surviving strains being Serratia sp. and H. alvei (Table 3). When these two strains were pretreated with either PowerFoam Plus or Tergazyme then PowerFoam Plus, followed by treatment with BioDestroy, neither strain could be cultivated (Table 3).
Log Reduction and Log Surviving (CFU/cm2) of Biofilms for Each Biocide Treatment
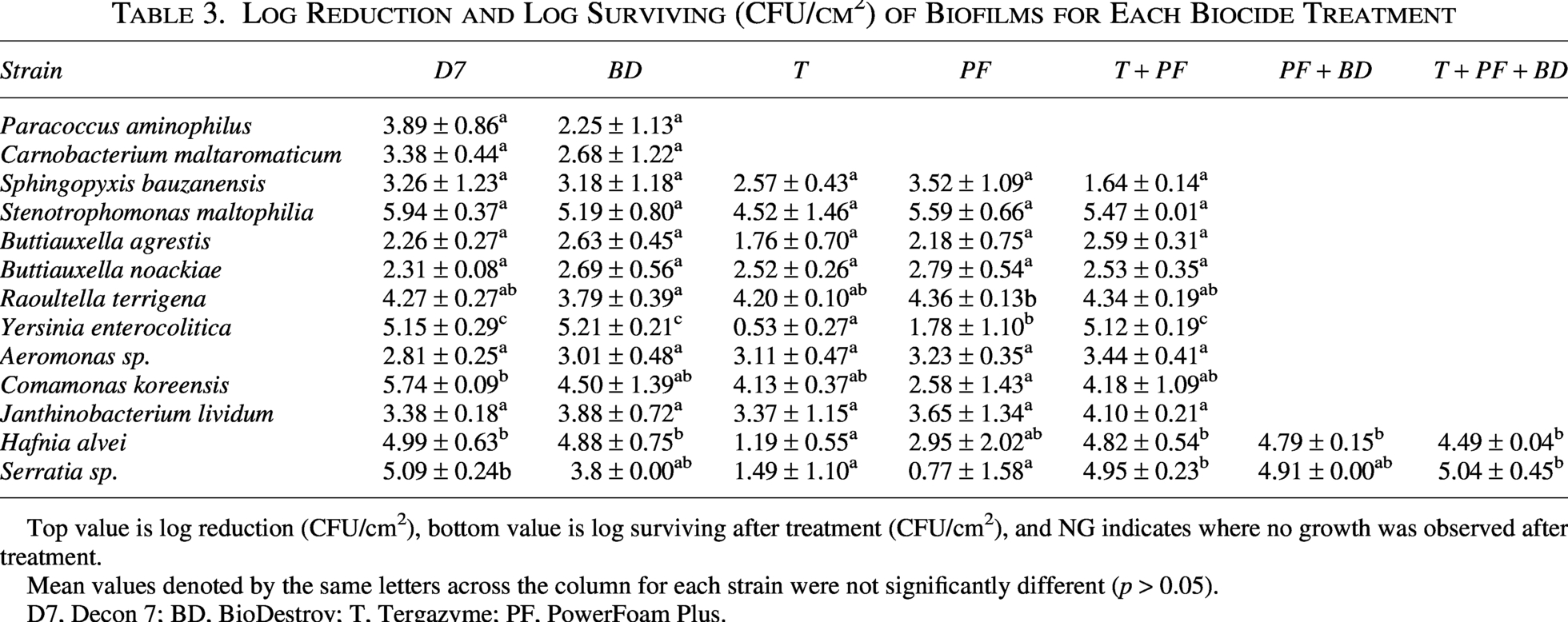
Top value is log reduction (CFU/cm2), bottom value is log surviving after treatment (CFU/cm2), and NG indicates where no growth was observed after treatment.
Mean values denoted by the same letters across the column for each strain were not significantly different (p > 0.05).
D7, Decon 7; BD, BioDestroy; T, Tergazyme; PF, PowerFoam Plus.
Discussion
In this study we analyzed the biofilm forming ability of 44 bacterial strains isolated from meat processors under conditions that simulated the harvest environment. Many of these strains are frequently encountered in meat-processing environments and linked to product spoilage, such as Pseudomonas, Carnobacterium, Serratia, Acinetobacter, Aerococcus, Moraxella, Lactobacillus, Yersinia, and Escherichia (Ercolini et al., 2009; Giaouris et al., 2014; Koti et al., 2024; Møretrø et al., 2013; Stellato et al., 2016; Wang et al., 2018). Biofilm forming ability is known to be mostly strain dependent and closely related to strain properties like expression of cell surface structures (Van Houdt and Michiels, 2005; Wang, 2019). Similar to previous studies using the MBEC peg model (Keelara et al., 2016), isolates in the present study formed biofilms under laboratory conditions, supporting their potential to persist on food-contact surfaces.
The strongest biofilm formers included A. lwoffii, C. koreensis, J. lividum, S. bauzensis, S. maltophilia, H. alvei, and Serratia sp. Interestingly, A. lwoffii or C. koreensis did not form strong biofilms in Lennox broth no salt (LB-NS). H. alvei and Serratia likely formed the strongest biofilms due to their robust production of EPS, complex quorum sensing regulation, and surface adhesion mechanisms such as type 1 fimbriae, outer membrane proteins and various adhesins (Bruhn et al., 2004; Giaouris et al., 2014; Moons et al., 2006; Souza Viana et al., 2009). Biofilm formation is affected by pH, temperature, osmolarity, O2 levels, nutrient composition and the presence of other bacteria (Giaouris et al., 2005, 2014; Hood and Zottola, 1997; Stepanović et al., 2003). These conditions are difficult to model in the laboratory, but we did include MJM in our media and dynamic temperature cycling between 8°C and 15°C to mimic these factors. Meat juices generate a conditioning layer of carbohydrates, proteins, lipids, and minerals on abiotic surfaces (Ma et al., 2020). Ma et al. (2020) found that diluted beef juice enhanced biofilm and extracellular matrix formation on stainless steel by E. coli as compared to minimal medium M9.
The same five GNA strains that did not develop measurable biofilms after 6 d at 15°C in LB-NS (Visvalingam et al., 2019), also did not form biofilms in this study. Furthermore, the LAB, Carnobacterium maltaromaticum strain that previously formed biofilms (Visvalingam et al., 2019), did not develop a measurable biofilm in the present study. Conversely, E. coli strain 136, previously identified as a strong biofilm former, only formed weak biofilms in this study, likely due to the use of different media and incubation temperature.
Compared to other steps in the cleaning and sanitation process in commercial settings, the chlorinated foam cleaner, PowerFoam Plus inconsistently reduced bacteria on conveyor belts (Wang et al., 2018; Yadav et al., 2025). Powerfoam Plus is more efficacious with a pre- or post-Tergazyme treatment, which helps disrupt EPS and proteins within biofilms. The protective EPS layer of biofilms plays a central role in neutralizing or impeding the diffusion and contact of biocides with cells embedded within biofilms (Koti et al., 2024; Wang, 2019). Furthermore, the effectiveness of sanitizers can be compromised by the presence of organic matter, which must be cleaned from equipment surfaces so as to not compromise biocide efficacy (Koti et al., 2024). However, even when surfaces are visibly clean, sufficient organic matter may remain to neutralize the oxidizing power of PowerFoam Plus (Wang et al., 2018).
Among the two food industry disinfectants, Decon 7 was the most effective at eliminating biofilms. As a multi-component formulation, Decon 7 simultaneously disrupts the EPS matrix, enabling deep penetration of the biocide into the dense biofilms formed by Hafnia and Serratia. It has also been shown to be effective at removing E. coli O157:H7 and Salmonella biofilms on meat (Wang et al., 2020). Our findings that biofilms formed by all 13 meat plant strains were eradicated by Decon 7 agrees with previous reports. In contrast, BioDestroy relies on a single mechanism of action that is more easily neutralized by organic residues and EPS. The effectiveness of BioDestroy was improved by pretreatment with PowerFoam Plus and Tergazyme, which disrupt the integrity of the EPS matrix. PowerFoam Plus is an alkaline surfactant that removes organic residues, and weakens the surface adhesion of embedded cells, whereas Tergazyme contains proteases that degrade proteins within the EPS matrix. Biofilms formed by Hafnia and Serratia spp. were not effectively removed by cleaning agents alone, nor was Serratia removed by BioDestroy. Effective removal of these recalcitarant biofilms required application of a cleaning agent prior to BioDestroy. Even with prolonged exposure, use of a single sanitizer may have limited effectiveness against biofilms (Visvalingam et al., 2024; Wang, 2019).
It is important to distinguish between biofilm inhibition and biofilm reduction when interpreting antimicrobial efficacy. As highlighted by Thieme et al. (2019), many biofilm studies evaluate treated biofilms only relative to untreated endpoint controls, without considering the initial mature biofilm biomass prior to treatment. This can lead to inhibitory effects being misinterpreted as biofilm reduction or eradication. In the present study, sanitizer efficacy was therefore interpreted within the context of established biofilms and changes in viable cell recovery following treatment.
The use of multiple cleaning and sanitizing products remains the best approach to removing meat plant bacterial biofilms. It is important to acknowledge that the monoculture biofilms formed on polystyrene peg surfaces in this study do not fully represent the complexity of biofilms within industrial meat processing environments. Multispecies biofilms may exhibit altered structural characteristics, enhanced extracellular matrix production, and increased tolerance to biocides compared to single-species biofilms. Additionally, surface composition can influence bacterial attachment and sanitizer efficacy. Additional studies are needed to further examine the effectiveness of biocide treatments on non-contact surfaces of meat plant equipment and their action against multi-species biofilms.
Authors’ Contributions
C.C.: Microbiological laboratory experiments and writing original draft; B.Y.: Statistical analyses; X.Y. and T.M.: Conceived and designed the study and provided funding. All authors revised the article and gave approval for the final version to be published.
Footnotes
Acknowledgments
The authors thank Sujeema Abeysekara and Chloe Brennan for their assistance in the laboratory.
Data Availability
Data are available from authors upon request.
Disclosure Statement
No competing financial interests exist.
Funding Information
Funding for this study was provided by the Beef Cattle Research Council (FOS.01.21) and the Results Driven Agricultural Research (2023F139R).