
Abstract
Campylobacter is a major foodborne pathogen of significant public health concern, as it is one of the leading causes of gastroenteritis worldwide. This bacterium has specialized growth requirements, such as specific temperature ranges, a microaerophilic environment (low oxygen levels), and nutrient-rich media. Consequently, the growth and detection of Campylobacter can be challenging, particularly for inexperienced laboratory personnel. Over time, researchers have developed various selective media to recover Campylobacter from diverse food and environmental matrices. However, the variable conditions of these media, along with ongoing efforts to optimize Campylobacter growth and detection, remain active areas of research. This review will focus on the factors that impact Campylobacter growth and recovery. By understanding the unique characteristics and growth requirements of this pathogen, researchers and laboratory personnel can improve their ability to identify and monitor Campylobacter, ultimately contributing to enhanced food safety and public health.
Introduction
Campylobacter species cause over 20% of bacterial foodborne illnesses in the United States each year (Batz et al., 2012; Scallan et al., 2011; Shah et al., 2024; Sodagari et al., 2025). In 2021, the European Union (EU) reported that campylobacteriosis accounted for more than 62% of all confirmed zoonotic cases in humans (European Food Safety Authority; European Centre for Disease Prevention and Control, 2022). In addition, Campylobacter species continue to develop antimicrobial resistance to broad-spectrum antibiotic agents such as florfenicol (Tang et al., 2020,2021, 2022; Yang et al., 2019). The illness caused by these bacteria is known as campylobacteriosis, a form of gastroenteritis characterized by diarrhea, fever, and abdominal pain (CDC, 2021). The illness is usually self-limiting and resolves within 2–5 days; 5–10% of cases may require hospitalization, and the fatality rate can be anywhere from 1 to 5 per 10,000 cases according to different estimates (Galanis, 2007). In rare instances, post-infection autoimmune disorders known as Guillain–Barré syndrome (GBS) and Miller-Fisher syndrome may occur (Chiba et al., 1992; van Belkum et al., 2009). These diseases are characterized by weakness of the limbs and, in some cases, paralysis due to nerve damage (Keithlin et al., 2014). Campylobacteriosis is most often caused by consuming contaminated poultry (Chlebicz and Śliżewska, 2018).
Campylobacter spp. are Gram-negative bacteria with a curved or spiral shape; most species are motile, with a flagellum at one or both ends, allowing them to move in a corkscrew-like manner (Lastovica et al., 2014). However, under stress, Campylobacter can change shape from a filamentous to a coccoid form (Gaynor et al., 2005; Tresse et al., 2017). Campylobacter possesses considerable diversity at the species, subspecies, and strain levels (Gaynor et al., 2005; Vidal et al., 2016). This diversity includes differences in genetic and phenotypic characteristics and extends into growth requirements (Vidal et al., 2016). The genus Campylobacter consists of 69 species and 18 subspecies (LPSN, 2026). Most human infections are attributed to C. jejuni and C. coli, but emerging Campylobacter spp. have been associated with clinical presentations such as gastroenteritis or bacteremia (Costa and Iraola, 2019; Kaakoush et al., 2015; Man, 2011). Some of these have been associated with meat products. For example, in addition to isolation of the better-known Campylobacter foodborne spp., numerous isolates from meat samples were recovered by Lynch et al. (2011) and identified. Of the chicken meat Campylobacter spp. isolated by Lynch et al. (2011), fastidious spp. C. concisus and C. mucosalis were identified, and minced beef isolates included C. concisus, C. curvus, C. mucosalis, C. sputorum, and C. upsaliensis. Thermotolerant species of Campylobacter (able to grow at 42°C), particularly C. jejuni, C. coli, C. lari, and C. upsaliensis, are the most probable species of Campylobacter to cause disease (Mikulić et al., 2016). The optimum growth temperature for most Campylobacter is either 42°C (chicken body temperature) or 37°C (human body temperature), and no Campylobacter can grow below 30°C (Hakeem & Lu, 2021). Campylobacter cannot survive under ambient oxygen levels due to (1) limited tolerance against reactive oxygen species, (2) inability to produce adequate antioxidant enzymes, (3) low respiratory rate, and (4) presence of essential enzymes that are destroyed by oxygen (Velayudhan et al., 2004).
The specificity of these growth requirements has presented challenges to the isolation and cultivation of Campylobacter spp. Consequently, considerable effort has been focused on optimizing enrichment and selective culture media to support consistent Campylobacter recovery and growth from various environmental matrices (Harrison et al., 2022). Optimizing media remains critical, as Campylobacter physiology and genetics must be better understood, particularly in specific environments they inhabit, such as the chicken gastrointestinal tract (GIT). Even with the emergence of molecular-based detection methods for Campylobacter, some culturing may still be required if pre-enrichment is required. Given the continued importance of Campylobacter as a foodborne pathogen and the need to cultivate it for further characterization, the development of media for its growth and detection is essential. This review will focus on Campylobacter metabolism, extrinsic factors that influence growth, and ecological considerations. It will also discuss a comprehensive understanding of Campylobacter, including its metabolism and subsequent requirements for growth and detection in culture media. A better understanding of Campylobacter growth requirements offers opportunities to refine and optimize growth media for characterization from various environmental matrices. The relationships among Campylobacter physiology, culture media design, sample matrix effects, environmental stressors, and detection outcomes are summarized in Figure 1, which provides a conceptual framework for understanding how each factor can influence recovery and interpretation.

Conceptual framework linking Campylobacter physiology, media formulation, matrix effects, and detection outcomes. Because Campylobacter growth depends on microaerophilic conditions, amino acid-based metabolism, temperature, species-level variation, and sensitivity to environmental stress, media selection can strongly influence recovery from complex samples. Matrix-associated factors, including competing microbiota, oxygen exposure, osmolarity, water activity, and prior cell stress, can further alter lag phase, culturability, and detection reliability. This framework highlights the need for media optimization that is both biologically informed and matrix specific. GIT, gastrointestinal tract; qPCR, quantitative polymerase chain reaction; VBNC, viable but nonculturable. Images created using BioRender. Olson, E. (2026) https://BioRender.com/3ely6ac.
Campylobacter Metabolic Requirements and General Requirements for Cultivation
Any development and improvement of alternative detection methods must consider Campylobacter’s metabolism and its ability to survive in unfavorable conditions. Not all Campylobacter metabolic processes are known; therefore, characterizing its complete metabolic profile can lead to the development of culture media with optimized nutrient availability. Understanding Campylobacter’s metabolism could prevent misdiagnosis and false negatives caused by difficulties in culturing with currently available media.
Campylobacter species are microaerophilic and capnophilic (CO2-loving) microorganisms that generally require 5% O2, 10% CO2, and 85% N2 for growth. Campylobacter species cannot use glucose as a carbon source and utilize only amino acids and Krebs cycle intermediates for energy production (Line et al., 2010; Stahl et al., 2012). Line et al. (2010) evaluated the ability of C. jejuni 11168 to oxidize 190 different substrates as sole carbon sources at 37°C and 42°C. They determined that the amino acids
Core Biological and Cultural Requirements That Shape Campylobacter Recovery

While C. jejuni is associated with most human illnesses, causing over 80% of Campylobacter infections (Hofreuter, 2014), C. coli and C. lari can also cause disease, though less frequently (Shah et al., 2024; Sodagari et al., 2025). Research indicates that no single medium can selectively grow C. jejuni while inhibiting C. coli or other species (Barbosa et al., 1999). In addition, some Campylobacter-selective media can result in the recovery of non-Campylobacter species (Bodie et al., 2025). This lack of defined, selective media for species other than C. jejuni makes it difficult to accurately identify and differentiate among Campylobacter species, which is vital for epidemiological purposes and for understanding the changing epidemiology of Campylobacter infections (Steinbrueckner et al., 1999). While considerable effort has been exercised in developing isolation and enrichment media for Campylobacter from a wide range of environmental, food, and clinical sources, there is a continued need for improvement as well as standardization (Harrison et al., 2022). Likewise, developing defined media for C. coli, C. lari, and other Campylobacter spp. requires research to identify the essential nutritional and growth requirements that may differentiate among the species. Any new media developed must be tested against a sufficiently representative panel of Campylobacter isolates to confirm specificity and sensitivity. These defined media would improve the identification and differentiation of Campylobacter species, which are essential for epidemiological surveillance and understanding the changing patterns of Campylobacter infections. However, before these can be developed, general concepts for Campylobacter culture media have to be considered to establish the baseline requirements that would need to be included regardless of the Campylobacter spp.
Culture Media: General Concepts
Over time, several culture media have been developed to recover Campylobacter from food matrices (Eberle and Kiess, 2012; El Baaboua et al., 2021; Gharst et al., 2013; Harrison et al., 2022; Ricke et al., 2019). The quality of culture media directly affects observations and inferences drawn. It can lead to confusing interpretations due to the cultural characteristics of competing microorganisms present in the selective media (Basu et al., 2015). Each medium has different parameters for metabolism and growth, which can lead to false-positive cell counts (Buss et al., 2019). Most Campylobacter species grow at approximately 37°C; however, some species, including C. jejuni, C. coli, C. lari, and C. upsaliensis, are thermotolerant and grow at 42°C (Hwang et al., 2014). For thermotolerant Campylobacter, several antimicrobial susceptibility tests and resistance breakpoints have been used in various studies and confirmed with genetic mutation detection assays (Guévremont et al., 2006; Luangtongkum et al., 2009). In addition, each selective medium has a unique battery of antimicrobials used for Campylobacter selection. As a result, the efficacy of antimicrobials will vary depending on the microbial communities present in complex samples and the Campylobacter species (Luangtongkum et al., 2009).
Media can influence the selectivity and detection of Campylobacter from samples; therefore, a better understanding of media can potentially improve future studies (Basu et al., 2015). One improvement could be the development of more rapid, efficient, and selective culture-based methods, such as the one described by Strakova et al. (2021) to improve detection rates for poorly filterable water. To accomplish this, Strakova et al. (2021) centrifuged water samples at high speed followed by resuspension of the resulting pellet and inoculation on a filter that was placed on Campylobacter selective modified charcoal cefoperazone deoxycholate agar (mCCDA). The concept is based on motile bacteria passing through the filter pores, while mCCDA suppresses background microbial growth. Another method would be to evaluate a broader range of enrichment broths and culture media to optimize the recovery of diverse Campylobacter species, as mentioned by Ricke et al. (2019). Hazeleger et al. (2022) suggested that determining the optimal enrichment broth and culture conditions for specific food matrices and Campylobacter strains would improve the sensitivity and reliability of detection, as well as the validation of new detection methods using a panel of well-characterized Campylobacter strains to ensure robust performance. Overall, a better understanding of how media composition and culture conditions impact Campylobacter detection can lead to the development of more rapid, sensitive, and comprehensive detection methods, an important goal given the public health significance of this pathogen.
When optimizing detection methods, the ability to reduce time and cost while limiting the growth of background microorganisms must be considered. Growth and detection of Campylobacter from foods can be problematic because high levels of competing organisms may be present (Bodie et al., 2025; Hill et al., 2017). Non-Campylobacter microorganisms present can outcompete Campylobacter for available nutrients. Consequently, enrichment media designed with nutrients that favor Campylobacter growth when a low number of Campylobacter cells is expected in the initial sample are critical. Campylobacter requires a microaerophilic atmosphere for active growth (Hill et al., 2017). Therefore, media must be nutrient-rich, contain oxygen-quenching agents, and, in the case of mixed populations, contain antibiotics to reduce background microorganisms (Solis-Soto et al., 2011). Variation across media makes it challenging to quantify Campylobacter, particularly to elucidate metabolic mechanisms (Tejera et al., 2020). For Campylobacter growth, direct plating and liquid-broth enrichment are the primary methods used, depending on the sampling technique and the number of Campylobacter cells present (Gonsalves et al., 2016). For example, when isolating C. jejuni from chicken fecal samples for quantification, C. jejuni is expected to be at population levels above 2.00 log10 CFU/mL; thus, these types of samples can be directly plated onto Campylobacter agar without prior enrichment (Musgrove et al., 2001; Sahin et al., 2003). However, Campylobacter spp. may not reach these levels when collected from processing samples. Consequently, Campylobacter samples from carcasses and environmental samples should be enriched in media before plating them onto agar for isolation (Rivoal et al., 2005). Since Campylobacter is a fastidious organism, challenges in detection persist. Sensitivity to environmental stress and external factors, such as temperature and competing microorganisms, can prevent growth and detection.
Campylobacter Growth Characteristics
Growth of bacteria such as Campylobacter implies the division of a bacterial cell, thereby increasing cell number (Margolin, 2005; Wang and Levin, 2009). Measuring and observing growth can be challenging because growth and death rates can be identical when other background microorganisms compete for the same nutrients (Axelsson-Olsson et al., 2007; Stahl et al., 2012). This challenge becomes evident with Campylobacter-selective media, in which non-Campylobacter organisms are recovered alongside Campylobacter (Bodie et al., 2025; Kim et al., 2017). This is supported by the occurrence of Campylobacter in the GIT of several food animals, and the synergistic relationship Campylobacter appears to have with other GIT organisms (Horrocks et al., 2009; Indikova et al., 2015; Olson et al., 2024a,b, 2025a).
There are a variety of laboratory approaches to measure growth. Direct cell counts are accurate and straightforward, performed by counting bacteria under a microscope (Marro et al., 2022). However, this method counts both living and dead cells and does not indicate the number of viable cells (Aryal, 2022). Further disadvantages include the potential to miss small cells and the requirement for a phase-contrast microscope unless staining is used (Aryal, 2022). Plate counts involve spreading a diluted bacterial sample on an agar plate and allowing it to grow into colonies. Theoretically, each colony represents a single bacterium from the original sample (Marro et al., 2022). However, clumps or chains of cells can form a single colony, leading to erroneous viable cell counts (Hazan et al., 2012). Plate counts are time-consuming and require a fixed amount of time for each stage, including dilutions, plating, incubation, and media preparation (Hazan et al., 2012). Campylobacter can be challenging to assess by plate counts because of its fastidious growth requirements and sensitivity to environmental stresses (Hakeem & Lu, 2021; He et al., 2024), and some Campylobacter cells may also enter a viable but nonculturable state, further complicating plate count methods (He et al., 2024).
Light scattering, based on the deflection of light by individual cells, essentially involves using a spectrophotometer to detect and quantify the level of turbidity resulting from the presence of bacterial cells blocking light transmission (Pirt, 1975). The advantage of using spectrophotometer-detected turbidimetry is that it is not only rapid but it is also nondestructive to the bacterial cells. Turbidity is most effective with suspensions of moderate density, as those with very high or very low density yield erroneous results (Maia et al., 2016). Bacteria exhibit typical growth curves comprising five phases: a lag phase, an exponential phase, a stationary phase, a death phase, and a long-term stationary phase (Baranyi and Roberts, 2000; Navarro Llorens et al., 2010; Pirt, 1975). Turbidimetric measurement of thermotolerant Campylobacter growth can require 6–7 h of incubation at 41.5°C before a sufficient detectable change in turbidity occurs (Hazeleger et al., 2016). However, Bodie et al. (2023) demonstrated that, even when using the same growth media, lag-phase durations can vary between C. jejuni, C. lari, and C. coli. Campylobacter can also form viable but nonculturable (VBNC) cells in response to various stressors, including low temperature, oxygen, acid, and salt treatments (Silva et al., 2011). These VBNC cells cannot divide in conventional culture media but retain membrane integrity and metabolic activity.
Protocols for the isolation of Campylobacter generally involve an enrichment phase in broth media (International Organization for Standardization ISO 10272-1:2017, 2017; USDA FSIS, 2021). Regardless of the procedure chosen, growth begins with a lag phase (Zwietering et al., 1990). The length of the lag phase is affected by any stress the cell has encountered before enrichment and by differences in serotype or strain (Booth, 2002; Jasson et al., 2007). Lanzl et al. (2020) assessed the variability in lag duration of C. jejuni and C. coli during enrichment after different food-relevant stress treatments to evaluate the impact on growth kinetics and reliability of detection outcomes. They subjected 13 C. jejuni and 10 C. coli strains to cold stress by refrigerating or freezing. They determined that refrigerated storage did not significantly reduce final cell counts, but frozen storage reduced cell concentrations by 1.6 log10 CFU/mL for both species. The mean lag duration for C. jejuni and C. coli without stress was 2.5 and 2.2 h, respectively. Refrigerated storage increased the lag duration for C. jejuni to 4.6 h and for C. coli to 5.0 h, and frozen storage increased the lag duration to 5.0 and 6.1 h for C. jejuni and C. coli, respectively. They performed a scenario analysis on the growth kinetics of Campylobacter species. They determined that the limit for successful detection was reached within 32.5 h, even in a worst-case scenario starting with a single cell. The probability of achieving the detection limit within 48 h was 100%, lending credence to the theory of competitive microbiota in enrichment, which leads to nondetection of Campylobacter rather than to any stress the cells experience before enrichment.
Numerous molecular methods for identifying and detecting Campylobacter, as well as for quantification, have been developed (Bodie et al., 2024; Eberle and Kiess, 2012; Gharst et al., 2013; Ricke et al., 2019). Polymerase chain reaction (PCR) uses differences in genome sequences among organisms to differentiate them (Ricke et al., 2019). Multiplex PCR targets multiple genes within a sample (Ricke et al., 2019). For example, Zhao et al. (2001) adapted previously published primers for a multiplex PCR to differentiate C. jejuni and C. coli after growth on blood agar plates. They tested 5% of colonies on the blood agar plates and found that 54% were C. jejuni and 41% were C. coli. The remaining 5% of colonies tested may have been other Campylobacter species not identified by the species-specific multiplex PCR, and the researchers concluded that multiple colonies must be tested with this method. Real-time PCR, or quantitative PCR, involves using fluorescent dyes that accumulate with each cycle, with an intensity directly proportional to the amount of target template DNA, and has been used for Campylobacter for a variety of sample sources as well as part of a multiplex assay for multiple foodborne pathogens including Campylobacter (Chapela et al., 2015; Klena et al., 2004; Park et al., 2011; Ricke et al., 2019).
Each of these methods has its advantages and disadvantages, and the choice of method often depends on the specific requirements of the experiment. For example, direct cell counts and plate counts methods directly measure the number of bacteria. In contrast, turbidity measurements provide a rapid and easy way to estimate bacterial numbers. Using traditional culture methods allows viable cell recovery, which researchers can use for phenotyping through plating and imaging. Some molecular approaches also require these methods, as Campylobacter cells must multiply in enrichment media before reaching sufficiently high levels for molecular assays such as PCR. Conventional PCR only detects the presence or absence of the target; multiplex PCR requires complex primers and may lead to both false positives and false negatives; qPCR is vulnerable to inhibitors that may be present in the matrix, needs specific primers, and does not differentiate live from dead cells (Ricke et al., 2019). Thus, consistency in culture approaches for enrichment to support microbiological and molecular detection methods is essential. In addition to baseline growth requirements, several extrinsic and methodological factors can interfere with Campylobacter culturability and interpretation; these factors and their implications for detection are summarized in Table 2.
Factors That Interfere with Culture-Based Recovery and Interpretation of Campylobacter Growth
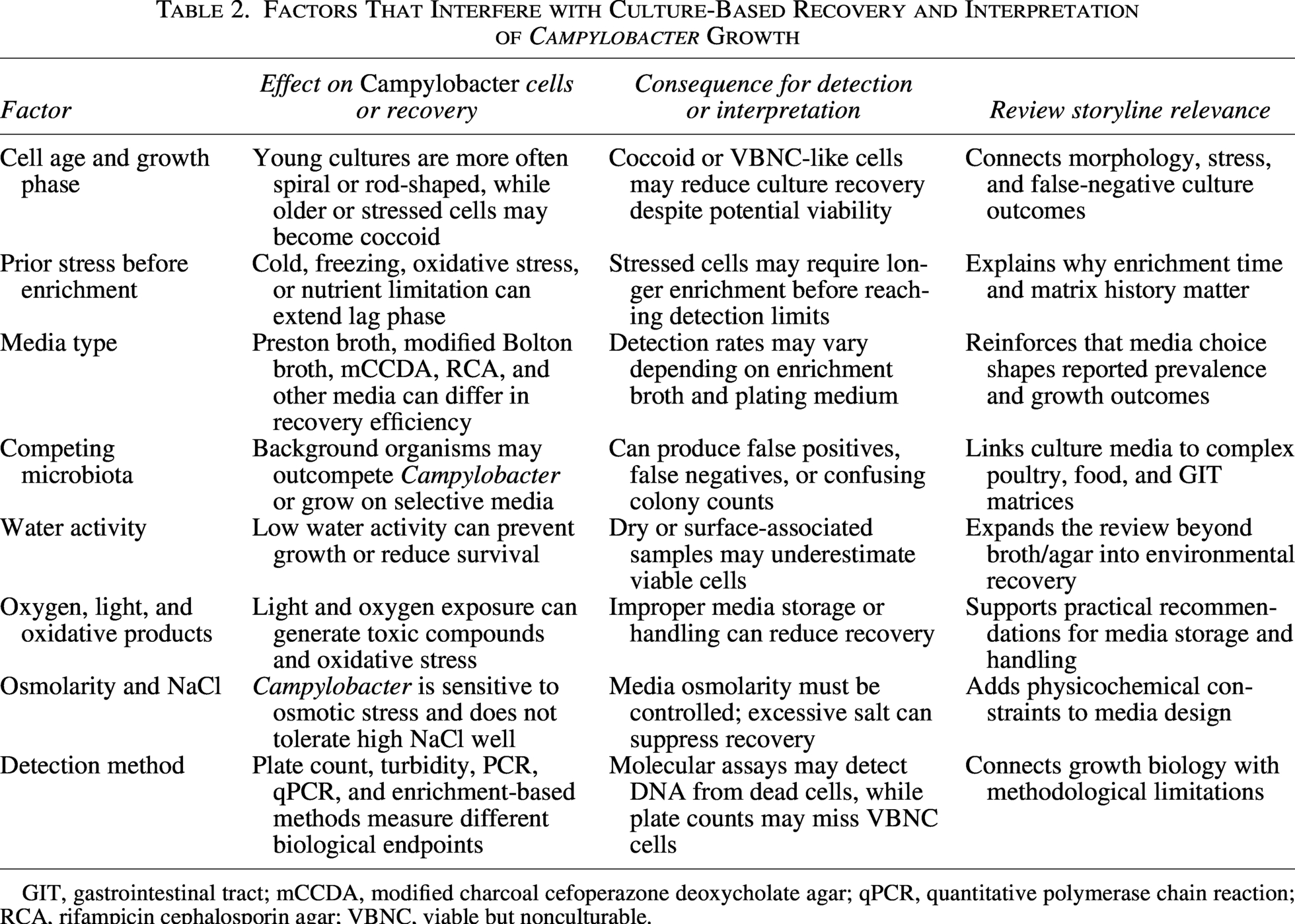
GIT, gastrointestinal tract; mCCDA, modified charcoal cefoperazone deoxycholate agar; qPCR, quantitative polymerase chain reaction; RCA, rifampicin cephalosporin agar; VBNC, viable but nonculturable.
Factors Affecting the Growth of Campylobacter Species
Cell age
When culturing Campylobacter, several morphological forms can be generated and identified visually. When pathogenic bacteria such as Campylobacter are released from their hosts into various environments, they are often challenged by environmental stresses, including nutrient availability, osmotic shock, temperature variation, pH, and oxidative stress (Lin et al., 2009; Pogacar et al., 2009). Young cultures, particularly in the early log phase, will form rod-shaped cells, including spiral- and S-shaped cells (Reezal et al., 1998). Coccoid forms occur in the stationary or death phase and when conditions are unfavorable for growth (Kelly et al., 2001). Studies have shown that as C. jejuni transitions from its helical to coccoid morphology, it enters a VBNC state (Beumer et al., 1992; Hazeleger et al., 1995). These nonculturable coccoid cells may still be infective to animals and humans and therefore still pose a health risk (Beumer et al., 1992; Hazeleger et al., 1995).
Media type
Harrison et al. (2022) have extensively reviewed the various media used for isolation and detection of Campylobacter spp. Culture media play an essential role in Campylobacter’s detection by influencing growth. For example, Seliwiorstow et al. (2016) demonstrated the impact of culture media on the recovery of Campylobacter from fresh and frozen poultry meat samples. In their study, after sampling poultry products, Campylobacter was analyzed using enrichment in Preston broth and enrichment in modified Bolton broth (supplemented with potassium clavulanate, triclosan, and polymyxin B). The enrichment cultures were streaked onto both mCCDA and Rapid Campylobacter agar (RCA). Both fresh and frozen poultry meat samples yielded more Campylobacter when enriched with modified Bolton broth than with Preston broth (Seliwiorstow et al., 2016). When comparing media, RCA yielded a higher detection rate than mCCDA. The results suggested that the type of media used can affect the growth and detection of Campylobacter species.
Since Campylobacter is a facultative intracellular pathogen with limited catabolic pathways, media must be supplemented with nutrients to support growth. In general, Campylobacter spp. are asaccharolytic and are unable to utilize carbohydrates for growth, forcing them to rely on amino acids (Bronowski et al., 2014). Nutrient concentration influences Campylobacter survival in the media (Thomas et al., 1999). Limited nutrient availability can lead to reduced cell size, changes in morphology, and shifts in protein synthesis (Cesar and Huang, 2017; Westfall and Levin, 2018). Lack of nutrients is one of the most common environmental stresses that Campylobacter species encounter. Mihalievic et al. (2007) determined the impact of selected environmental stress factors, including temperature shift, starvation, and the concentration of atmospheric oxygen, on the subsequent ability to cultivate C. jejuni along with the viability of two C. jejuni isolates. The results of these studies show that nutrient deprivation led to fewer culturable Campylobacter cells than exposure to temperature shifts, such as heat stress (55°C) and oxidative stress (atmospheric oxygen exposure). Media are typically supplemented with peptones and dextrose to provide a source of energy for growth (Lagier et al., 2015). The peptones provide nitrogenous compounds, carbon, sulfur, and trace ingredients, while dextrose is utilized as an energy source. More recently, Olson et al. (2024a) demonstrated that gut-habitat-simulating media supported C. jejuni and its interactions with the non-Campylobacter population. Using such media demonstrated the reliance of C. jejuni on the other members of the gut microbial community and their metabolic products (Olson et al., 2024b, 2025a).
Water activity
Water activity has also been shown to affect Campylobacter growth, thereby reducing detection. For example, Cesare et al. (2003) attempted to estimate the length of time over which one could reasonably expect pathogen survival and, hence, the potential for cross-contamination. A five-strain pool of C. jejuni was suspended in either a phosphate-buffered saline solution or Trypticase soy broth and inoculated on samples of Formica® laminate, glazed ceramic tiles, polished stainless steel, and 100% cotton dishcloths. Samples inoculated with C. jejuni did not grow at water activities below 0.987 (Cesare et al., 2003).
Osmolarity and light
Hoffman et al. (1979) observed that the growth of Campylobacter spp. was substantially reduced when nutrient media plates were stored in light and air. These environmental changes can generate toxic compounds, free radicals, and hydrogen peroxide, which can injure cells and negatively affect Campylobacter growth (Hofreuter, 2014). Media for Campylobacter species usually contain ingredients to neutralize the toxic effects of substances formed when oxygen and light are present (Corry et al., 2003). Oxygen-quenching agents in the media can significantly improve Campylobacter recovery by protecting it from harmful agents such as light and oxygen (Oyarzabal et al., 2005). Oxygen-quenching agents, such as blood, hemin, charcoal, oxyrase, ferrous sulfate, sodium pyruvate, and sodium metabisulfite, can be added to Campylobacter media to improve growth.
As Campylobacter is removed from its environment and handled as a laboratory culture, it becomes susceptible to osmotic stress when transferred to an enrichment medium. Osmolarity is a measure of solute concentration in a solution (Cameron et al., 2012). When grown in liquid media, Campylobacter may be hyperosmotic or dehydrated, depending on the medium. These changes in osmolarity will cause substantial stress on cells, causing them to swell or shrink (Burgess et al., 2016). Furthermore, Campylobacter is more sensitive to NaCl than other pathogenic GIT pathogens, such as E. coli and Salmonella, which can survive for long periods when exposed to 30% NaCl (Cameron et al., 2012; Doyle and Roman, 1982; Kempf and Bremer, 1998; Wood, 1999). Campylobacter cannot survive when growth media are supplemented with NaCl above 2% due to osmotic stress; as water leaves the cell, cytoplasmic dehydration occurs, and turgor is reduced because Campylobacter cannot adapt to these conditions (Cameron et al., 2012). Consequently, NaCl is kept at minimal to absent levels in selective media such as Mueller–Hinton and mCCDA (Hofreuter et al., 2012; Trigui et al., 2015). Campylobacter can also become stressed when grown in liquid media lacking sufficient nutrients or in low-osmotic environments (Brandl et al., 2004; Reezal et al., 1998). Moran and Upton (1986) reported that low osmolarity in C. jejuni cells led to cytoplasmic leakage and the degradation of cellular components. These findings suggest that Campylobacter has an osmolarity threshold for survival in media.
Conclusions and Future Directions
Campylobacter cells can exhibit different morphological forms, influenced by environmental stresses such as nutrient availability, osmotic shock, temperature variation, pH, and oxidative stress. Rod-shaped Campylobacter cells in the early log phase can transform into coccoid forms when conditions are unfavorable for growth, such as a decrease in temperature or nutrient availability. Campylobacter cells can become reduced in size either through multiple divisions without an increase in size or due to insufficient nutrients for cell division, leading to a decrease in cell size until death. Environmental factors, such as exposure to light and air, can negatively affect Campylobacter growth, but the use of oxygen-quenching agents in the media can improve recovery.
In addition, Campylobacter is a diverse genus of bacteria found in various sources, including humans, animals, and environmental samples. While multiple media and methods are available for growing and detecting Campylobacter, including enrichment broths and selective agar plates, the resulting growth responses are not always equivalent across Campylobacter spp. The growth and detection of Campylobacter can be challenging due to its complex growth requirements and the presence of competing microorganisms. There is a need for further research to optimize enrichment procedures, improve detection methods, and understand the epidemiology and pathogenicity of different Campylobacter species. Advances in genomic-metabolomic modeling offer opportunities for further media refinement (Tejera et al., 2020). More recent developments in general anaerobic, less selective media that approximate GIT environments, such as chicken cecal contents, offer an opportunity to examine Campylobacter interactions with the indigenous GIT microbiota (Olson et al., 2024a). Preserving GIT ecology in vitro is essential for establishing both synergistic and antagonistic interactions between Campylobacter and indigenous GIT microbiota (Olson et al., 2024b, 2025a). Further characterization of GIT metabolites and the microbial community should improve the development of GIT-representative media (Chatman et al., 2024; Olson et al., 2025b). Alternative methods, such as molecular techniques, can provide faster and more accurate results for Campylobacter detection, but may still require some form of culturing before the molecular assay. In addition, most PCR- and qPCR-based molecular detection approaches cannot distinguish between living and dead bacteria (Ricke et al., 2019). However, recovery of RNA or employment of fluorescent dyes in qPCR assays such as ethidium and propidium monazide, that only penetrate dead cells can be used to differentiate live from dead cells (Ricke et al., 2019).
Further research and development of selective media that provide better selectivity while retaining Campylobacter optimal growth, especially in samples with high levels of competing microorganisms, is needed. Comparative studies should also be conducted to evaluate the performance of different enrichment media and selective agars for Campylobacter detection across a broader range of sample types, including the GIT of food animals. These studies may be critical for designing practical preharvest intervention strategies, such as bacteriophages and probiotics (Deng et al., 2020; Olson et al., 2022). Likewise, rapid and sensitive detection methods for Campylobacter should be developed and used in conjunction with enrichment media to improve the efficiency and accuracy of Campylobacter detection and quantitation in food samples. Standardizing culture media and detection methods would ensure more consistent and reliable results. Researchers, public health agencies, and food industry stakeholders should collaborate to develop strategies for reducing Campylobacter contamination in poultry and other food sources. Education and training of laboratory personnel is also a priority to ensure proper handling and detection of Campylobacter in food samples.
Authors’ Contributions
S.C.R.: Conceptualization, investigation, original draft preparation, graphics, review, and final editing. A.R.B.: Conceptualization, investigation, review, editing, and original draft preparation. C.A.O. and M.J.R.: Review and editing. E.G.O.: Review, graphics, and final editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.