
Abstract
A classical acute porphyria model in rats consists of combined treatment with 2-allyl-2-isopropylacetamide (AIA) and 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC). The present work describes the effects of this treatment on the pentose phosphate (PP) pathway, glutahione metabolism and redox state and how they contribute to alter the glucose pool of hepatocytes and modulate porphyria, in Wistar rat livers. Our approach is based on the fact that glucose is a repressor of 5-aminolevulinic synthase (ALA-S), the rate-limiting enzyme of the heme pathway, and treatment with AIA/DCC causes oxidative stress. Different doses of the xenobiotcs were used. The results show that AIA (500 mg/kg body weight [BW])/ DDC (50 mg/kg [BW]) treatment increased glutathione peroxidase (GPx) activity by 46%, decreased both glutathione reductase (GR) and glutathione S-transferase (GST) activity by 69% and 52%, respectively, and reduced by 51% reduced glutathione (GSH) and increased by 100% glutathione disulfide (GSSG) concentrations, therefore lowering by four-fold the GSH/GSSG ratio. The activity of glucose-6-phosphate dehydrogenase (G6PD), the rate-limiting enzyme of PP-pathway, was increased by 129% as well as that of 6-phosphogluconate dehydrogenase. NADPH and the NADPH/NADP+ ratio were increased by 14% and 28%, respectively. These effects could be attributed to the generation of reactive oxygen species (ROS) elicited by the porphyrinogenic treatment, shown by enhanced DNA damage and ROS production. G6PD stimulation would decrease hepatic glucose concentrations and consequently exacerbate the porphyria. A decrease in glucose could stimulate ALA-S and this would add to the effect of drug-induced heme depletion. Since the key role of GST is to inactivate toxic compounds, the drastic fall in its activity together with the accumulation of ALA would account for the symptoms of this hepatic disease model. The present findings show the high metabolic interplay between pathways and constitute a relevant contribution to achieve a better treatment of acute human porphyria.
Keywords
Introduction
Porphyrias are hereditary disorders caused by de-regulation of the heme pathway due to enzyme deficiencies leading to a lower heme formation. This triggers the induction of the regulatory enzyme ALA-synthase (ALA-S). 1
Acute porphyria, a life-threatening condition, is biochemically characterized by the accumulation of heme precursors such as ALA, which generates reactive oxygen species (ROS) resulting in oxidative stress.
The porphyrinogenic drug 2-allyl-2-isopropylacetamide (AIA) promotes the destruction of liver heme, particularly cytochrome P-450. 2 Another porphyrinogenic drug, 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC), strongly depletes hepatic heme due to a combination of heme destruction and inhibition of heme synthesis. 3 AIA/DDC treatment results in an acute heme deficiency and a marked de-repression of ALA-S and, consequently, in an exacerbated production of ALA and other heme precursors in the liver. 4 This combined treatment has been reported to induce an experimental porphyria accurately resembling acute variegate porphyria, in rats. 4 An accumulation of ALA has been associated with iron-mediated oxidative damage to biomolecules and cell structures 5 through the generation of ROS. 6 ROS are able to oxidize nucleic acids, proteins, lipids and carbohydrates, impairing key cellular functions. 7 It has been demonstrated that AIA/DDC treatment promotes an oxidative environment with ROS increases. 4
Glucose administration is known to have beneficial effects on acute porphyria patients significantly improving their clinical and biochemical condition. 1 In animal models, the prevention of acute experimental porphyria through high carbohydrate and/or protein intake 8 illustrates the effect of glucose on ALA-S, with carbohydrates preventing the induction of this heme-pathway regulatory enzyme. 9
On the other hand, it has been reported that AIA/DDC treatment promotes gluconeogenic and glycogenolytic blockages leading to a reduced glucose availability in hepatocytes. In this respect, hepatic phosphoenolpyruvate car-boxykinase and glycogen phosphorylase activities have been found to be impaired in this experimental porphyria, in rats.4,10
The pentose-phosphate (PP) pathway is primarily an anabolic pathway that uses the six carbon atoms from glucose to generate five carbon sugars and reducing equivalents. In this pathway glucose can be completely oxidized to CO2 and water. In fact, 30% of glucose oxidation in the liver occurs via the pentose pathway. Glucose-6-phosphate de-hydrogenase (G6PD, EC1.1.1.49) is the regulatory enzyme of the PP pathway supplying riboses for DNA and RNA synthesis. 11 However, its main function is the production of NADPH, the main cytoplasmic reducing agent. It has been shown that G6PD function is essential in the oxidative stress defense-NADPH dependent. 12 The NADPH pool regulates the levels of reduced glutathione (GSH) which, in turn, is in charge of eliminating harmful compounds such as peroxides from cells. GSH is in fact an antioxidant, and prevents damage to important cellular components caused by ROS. 13 It reduces disulfide bonds formed within cytoplasmic proteins to cysteines by acting as an electron donor. In the process, GSH is oxidized to glutathione disulfide (GSSG). Glutathione is found almost exclusively in its reduced form, since glutathione reductase (GR, EC 1.8.1.7), the enzyme that re-generates GSH from its oxidized form, is constitutively active and inducible. 14 GR is thus a critical enzyme to maintain a proper redox status. It is a FAD-containing enzyme that catalyzes the NADP-dependent reduction of GSSG to GSH. Other GSH metabolism enzymes are glutathione peroxidase (GPx) (EC 1.11.1.9) and glutathione S-transferase (GST, EC 2.5.1.18) (Figure 1).
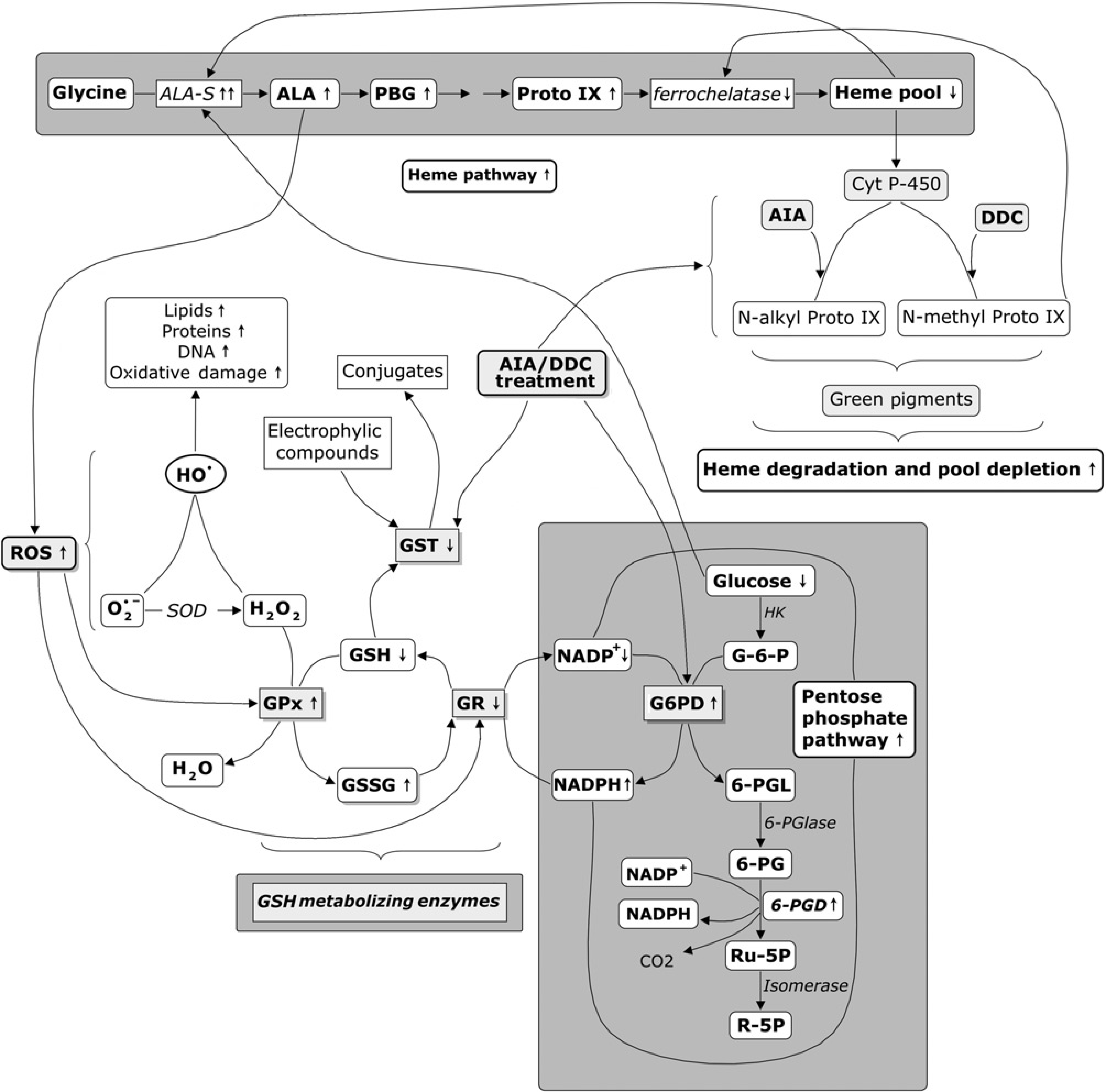
Scheme on the effect of AIA/DDC treatment on different metabolic routes (pentose phosphate pathway, heme pathway, ROS generation and GSH metabolism) leading to oxidative stress, toxic response, glucose decrease and porphyria exacerbation. AIA, 2-allyl-2-isopropylacetamide; ALA, 5-aminolevulinic acid; ALA-S, ALA-synthase; DDC, 3,5-diethoxycarbonyl-1,4-dihydrocollidine; Cyt P-450, cytochrome P-450; GSH, reduced glutathione; GSSG, glutathione disul-fide; G-6-P, glucose-6-phosphate; G6PD, glucose-6-phosphate dehydrogenase; GPx, glutathione peroxidase; GR, glutathione reductase; GST, glutathione S-transferase; GSH enzymes, enzymes of glutahione metabolism; HK, hexoquinase;
GPx are a family of selenium-containing enzymes that use GSH to scavenge peroxides when converting GSH to GSSG, thus protecting tissues from oxidative damage. GPx1 is the most abundant form and is expressed in cell types where the preferred substrate is hydrogen peroxide.
The GST family represent a major group of detoxification enzymes. All eukaryotic species possess multiple cytosolic and membrane-bound GST isoenzymes, each of which displays distinct catalytic as well as non-catalytic properties. The cytosolic isoenzymes are encoded by at least five distantly related gene families. 15 GSTs are up-regulated by xenobiotics, drugs, cytokines and endotoxin. 16 At least 100 chemicals have been identified as GST inducers. 15 On the other hand, hematin, bilirubin, biliverdin, biliary acids and halogenated compounds, among others, have been found to inhibit hepatic GST.17,18
As stated above, AIA/DDC treatment (model of acute porphyria) results in oxidative stress with increased ROS production. AIA/DDC treatment also promotes gluco-neogenic and glycogenolytic blockages leading to a lower glucose availability in hepatocytes. In this respect, it has been reported that glucose plays a key role in the regulation of the heme pathway, as well as in the treatment of human and experimental porphyrias. Considering all this, it seems interesting to study how the PP-pathway could be affected in the AIA/DDC porphyria model. This pathway consumes glucose and generates NADPH involved in redox reactions. It is also relevant to investigate the GSH metabolizing enzymes GR, GPx and GST, responsible for maintaining the status of GSH, the major endogenous cell antioxidant. The ultimate purpose is to establish the contribution of alterations in the glucose pool and the redox state of hepatocytes to this type of experimental hepatic porphyria, as well as the inter-relations of these metabolic pathways and their impact on the regulation of the heme pathway. This is the first report on the pentose pathway and glutathione metabolism in the AIA/DDC hepatic porphyria model.
Materials and methods
Drugs and chemicals
AIA (99% pure) was a gift from Roche Co. (Basel, Switzerland). DDC (99% pure) was purchased from Aldrich Chemical Company Inc. (Milwaukee, WI, USA). Bovine serum albumin (BSA), 1-chloro,2,4-dinitrobenzene (CDNB), dextrose, dinitrophenyl hydrazine, 2’,7’-dichloro-dihydrofluorescein diacetate (DCFH-DA), 1,2-dithiobis-nitrobenzoic acid (DTNB), ethylenediaminetetraacetic acid (EDTA), GR, guanidine hydrochloride, horseradish peroxidase, nicotinamide adenine dinucleotide phosphate (NADP), reduced nicotinamide adenine dinucleotide phosphate tetra sodium salt (NADPHNa4), ortho-phosphoric acid, oxidized glutathione (GSSG), phenol red, reduced glutathione (GSH), sodium dithionite, sulfosalicylic acid, Glucose (HK) Assay Kit and thiobarbituric acid were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). Bioxytech 8-OHdG-EIA Kit was obtained from Oxis Health Products, Inc. (Portland, OR, USA). All other chemicals were of analytical grade.
Animal treatments
Animals were treated according to International Guidelines (Guide for Care and Use of Laboratory Animals, National Research Council, USA, 1996, the Council of the European Communities Directive, 86/609/ECC) and also to guidelines from the Animal Care and Use Committee of the Argentine Association of Specialists in Laboratory Animals (AADEALC). Adult female Wistar rats (200–250 g body weight [BW]) purchased from the National Committee of Atomic Energy (CONEA Argentina) were used. Animals were fasted for 24 h (8 h before treatment and 16 h after treatment) before euthanasia. They were kept on rat chow and water ad libitum and housed in stainless-steel individual metabolic cages under conditions of controlled temperature (25°C) and light (12 h light-dark cycle from 06:00 to 18:00) for collection of 24-h urine. All in vivo experiments were carried out between 09:00 and 11:00. AIA was dissolved in saline solution (1.5 w/v)
19
and DDC (10 mg) was dissolved in 2.5 mL corn oil.
20
Both solutions were prepared just before use. Three different AIA doses were injected subcutaneously, whereas a single DDC dose was injected intraperitoneally. Rats were randomly divided into the following groups: group L (100 mg AIA
Blood samples were collected with a heparinized capillary tube from the ophthalmic venous plexus just before euthanasia and then centrifuged at 1000 × g for 10 min at 4°C. Plasma samples were kept at −70°C until ROS determination.
Rats were euthanized by decapitation 16 h after AIA/DDC administration. All assays were performed in animals with a total fasting period of 24 h. Fasting period enhances assay sensitivity and facilitates the standardization of the results obtained with respect to the control. On the other hand, since the effect of AIA depends on the previous diet, the eight-hour starvation period prior to intoxication guaranteed that the effect of the drug was not altered.
Hepatic tissue was quickly removed, perfused with ice-cold saline (0.85%, w/v NaCl). Small pieces of all the hepatic lobes were taken and homogenized in the adequate buffer according to the assay.
ALA-S activity
A fraction of liver (1 g) was immediately homogenized (1:3 w/v) in a solution containing 0.9% NaCl, 0.1 mmol/L Tris-HCl, pH 7.4 and 0.5 mmol/L EDTA to determine hepatic ALA-S activity by using the method of Marver et al. 22 Incubation mixtures containing 0.1 mol/L glycine, 0.01 mol/L EDTA, 0.08 mol/L Tris-HCl buffer pH 7.2, and 0.5 mL homogenate, final volume 2 mL, were incubated at 37°C for 60 min. The product was determined spectrophoto-metrically at 553 nm as described by Mauzerall and Granick. 23
Urinary 8-hydroxy-2’-deoxyguanosine concentrations
8-Hydroxy-2’-deoxyguanosine (8-OHdG) concentrations, in the 24 h urine samples were determined using the Bioxytech-8-OHdG-Q2 G-EIA Kit, Oxis Health Products, Inc. (Portland, OR, USA). This kit is a competitive enzyme-linked immunosorbent assay for quantitative measurement of 8-OHdG in urine employing the 8-OHdG monoclonal antibody and a secondary antibody conjugated to horseradish peroxidase. 3,3’,5,5’-Tetramethylbenzidine was used as an electron donor for the reduction of hydrogen peroxide. The resulting blue color was read on a Stat Fax Microplate reader (Awareness Technology, Inc., Palm City, FL, USA) at 450 nm.
Because of the variability among animals and the significative difference in the urinary volume excreted, the 8-OHdG concentrations were normalized in relation to the amount of creatinine. Creatinine concentrations in the 24 h urine samples were determined using the creatinine Direct-Test (Wiener Lab. Group, Rosario, Argentina).
Plasmatic ROS concentrations
To reduce the variation in ROS over time, the plasma was immediately stored at −70°C and analyzed as soon as possible (within 10 min). For plasmatic ROS determination the cell-free assay for DCFH oxidation was followed as described by Foucaud et al. (2010). 24 Immediately before use, DCFH-DA was chemically hydrolyzed at basic pH in the presence of 0.01 N NaOH for 30 min, in the dark and at room temperature. 25 The mixture was neutralized with 0.1 mol/L phosphate-buffered saline pH 7.4. The reaction mixture was prepared by adding 40 μL DCFH solution to 160 μL of 0.9% NaCl containing 0.1% BSA and 0.02 units of horseradish peroxidase to obtain a final concentration of 10 μmol/L DCFH. Plasma samples were added (20 μL) to reaction mixture. The fluorescence generated by the DCFH oxidation was measured after 400 s at 485 nm excitation and 530 nm emission in a FLUORstar Optima, fluorescence plate reader (Ortenberg, Germany).
GR activity
The supernatant fraction from a 1:4 liver homogenate in 0.1 mol/L potassium phosphate buffer and 1 mmol/L EDTA (pH 7.5) was prepared centrifuging at 11.000 × g for 20 min. GR activity was measured by the method of Mohandas et al. 26 The incubation mixture consisted of 1.65 mL phosphate buffer (0.1 mol/L, pH 7.6), 0.1 mL EDTA (0.5 mmol/L), 0.05 mL GSSG (1 mmol/L), 0.1 mL NADPH (0.1 mmol/L) and 0.1 mL enzyme, total volume 2.0 mL. Enzyme activity was determined at 25°C measuring the disappearance of NADPH at 340 nm, and was calculated as nmol NADPH oxidized per min mg protein using a molar extinction coefficient of 6.223 × 103 L/(mol cm).
GST activity
The supernatant fraction from a 1:4 liver homogenate in phosphate buffer (0.1 mol/L, pH 7.4) that contained KCl (1.17%, w/v) was prepared in a Potter Elvehjem homogenizer centrifuging at 11.000 × g for 20 min. GST activity was determined by the method of Haque et al. 27 The assay mixture consisted of 1.6 mL sodium phosphate buffer (0.1 mol/L, pH 7.4), 0.2 mL reduced glutathione (1 mmol/L), 0.1 mL of CDNB (1 mmol/L) and 0.1 mL of enzyme, total volume 2 mL. The absorbance change was recorded at 340 nm and enzyme activity was calculated as nmol CDNB conjugates formed per min mg protein using a molar extinction coefficient of 9.6 × 103 L/(mol cm).
GPx activity
Supernatant fraction from a 1:4 liver homogenate was obtained according to section 2.4.3.2. GPx activity was measured following the method of Haque et al. 27 The assay mixture consisted of 1.44 mL sodium phosphate buffer, 0.1 mL EDTA (1 mmol/L), 0.1 mL sodium azide (1 mmol/L), 0.05 mL of GR (1 IU/mL), 0.1 mL GSH (1 mmol/L), 0.1 mL NADPH (0.02 mmol/L), 0.01 mL H2O2 (0.25 mmol/L) and 0.1 mL of enzyme, total volume 2 mL. NADPH oxidation was recorded spectrophotometrically at 340 nm at room temperature. Enzyme activity was calculated as nmol NADPH oxidized per min mg of protein, using a molar extinction coefficient of 6.22 × 103 L/ (mol cm).
Thiol profile
Total thiols content (non-protein and protein-bound sulfhy-dryl groups) (T-SH) in rat liver was determined in homogenate 1:5 in 0.1 mol/L, Tris-HCl (pH 8.2) using Ellman's reagent, according to the method of Sedlak and Lindsay. 28 Values were expressed as μmol/g of tissue.
Total glutathione content (GSH plus GSSG) was determined spectrophotometrically at 412 nm in 11.000 × g supernatant by yeast-GR recycling method at room temperature using 5,5’-dithio-bis-(2-nitrobenzoic acid) (DTNB) and NADPH. Reference rate was established using a freshly prepared GSH standard curve Tietze. 29 GSSG was determined by the same method in the presence of 2-vinylpyridine, and GSH was calculated from the difference between total glutathione and GSSG. 30 GSH and GSSG contents are expressed as nmol/mg protein.
G6PD and 6-phosphogluconate dehydrogenase activities
The supernatant fraction from a 1:4 liver homogenate in 0.15 mol/L KC1 was prepared centrifuging at 105,000 × g for 60 min. G6PD activity was assayed according to a Bottomley et al., 31 which corrects for the 6-phosphogluconate dehydrogenase (6-PGD) present in liver homogenates. The incubation mixture contained 120 μmol Tris-ClH buffer (pH 8), 2 μmol glucose-6-P, 0.9 μmol NADP+, 10.4 μmol MgC12, 0.6 μmol 6-P-gluconate and the enzyme (final volume 1.0 mL). The activity of 6-PGD was determined eliminating glucose-6-P from the above mixture.
Then the activity of G6PD was obtained by the difference between the two measurements.
The activity of 6-PGD was determined eliminating glucose-6-P from the assay mixture. Changes in absorbance were recorded at 340 nm at room temperature, and enzymes activities calculated as nmol NADP+ reduced per min mg protein using a molar extinction coefficient of 6.223 × 103 L/(molcm). Enzymes activities were expressed as nmol NADPH produced per minute.
Determination of pyridine nucleotide phosphates
Liver was homogenized (10% w/v) in cold 5% HClO4 and stored for no less than 30 min in ice to complete the denaturalization of proteins. The samples were centrifuged at 1300 × g for 10 min and the clear supernatants were neutralized with 2 N KOH in the presence of bromo thymol blue 0.1% in 95% ethyl alcohol until a pale yellowish-green color was obtained. After 30 min in an ice bath for complete precipitation of KClO4, NADP+ and NADPH concentrations were determined by the enzymatic method described by Zhang et al. 32
Protein determination
Protein concentration was measured following Lowry et al., 33 using bovine serum albumin as standard.
Liver glucose concentration
Glucose was extracted by treatment of liver tissue samples with an ice-cold 0.5 mol/L perchloric acid solution. After centrifugation and neutralization, aliquots of the extracts were used. 34 Glucose concentrations were estimated by a commercial Glucose Assay Kit in accordance to the manufacturer's instructions.
Statistical analysis
Results are the means ± SEM of nine animals per group, since three different experiments including three animals per group were performed. Data were subjected to one-way analysis of variance (ANOVA). When the overall F-statistic was significant, multiple comparisons between treatment groups were performed using Tukey–Kramer's test The level of significance used was 0.05. Statistical analyses were performed with GraphPad Prism 4.0 for Windows (GraphPad Software, San Diego, CA, USA). 35
Results
Hepatic ALA-S activity (porphyria marker)
ALA-S, the key regulatory enzyme of the heme biosynthetic pathway, was used as a porphyria marker. ALA-S hepatic activity showed dose-dependent and statistically significant increases in the three groups studied (Figure 2). These increases were 68% in group L, 209% in group M and 355% in group H when compared with group C. Enzymatic activities in groups H and M were also statistically different from those of group L.
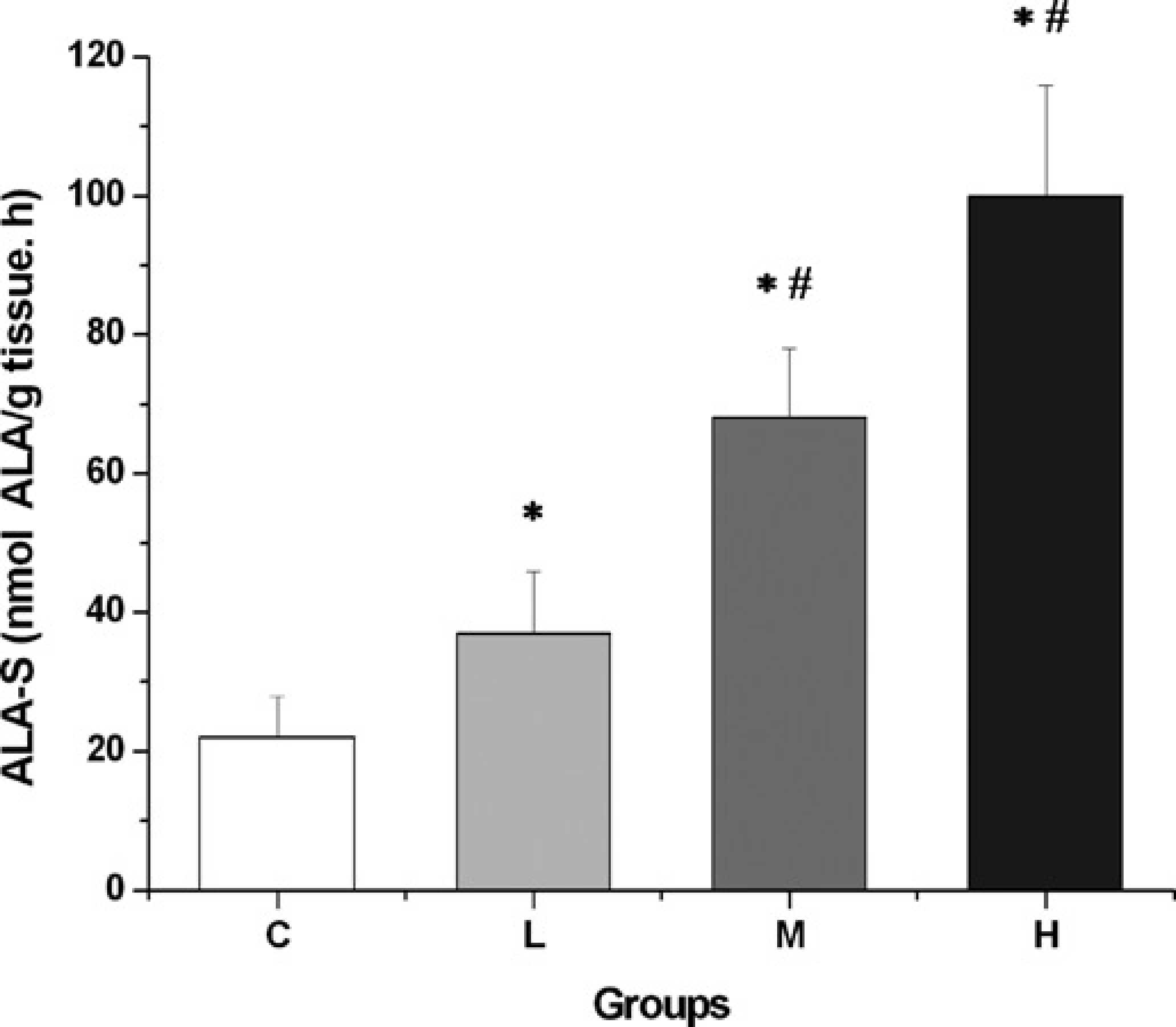
Effect of AIA/DDC treatment on ALA-S activity according to AIA dose. Animals were injected subcutaneously with three different AIA doses and intraperitoneally with a single DDC dose. Then they were divided into the following groups: group
Oxidative stress markers
In order to evaluate oxidative stress and the damage that it causes to biomolecules, DNA damage and ROS production were determined.
As it can be seen in Figure 3a, 8-OHdG was evaluated in urine in relationship with creatinine concentrations. Results obtained (Figure 3a) showed a marked DNA damage with high, significant increases of 595%, 1451% and 2,971% for groups L, M and H respectively when compared with controls. Levels of 8-OHdG corresponding to groups M and H were also statistically higher than group L and the values from group H were also significantly different from those of group M.
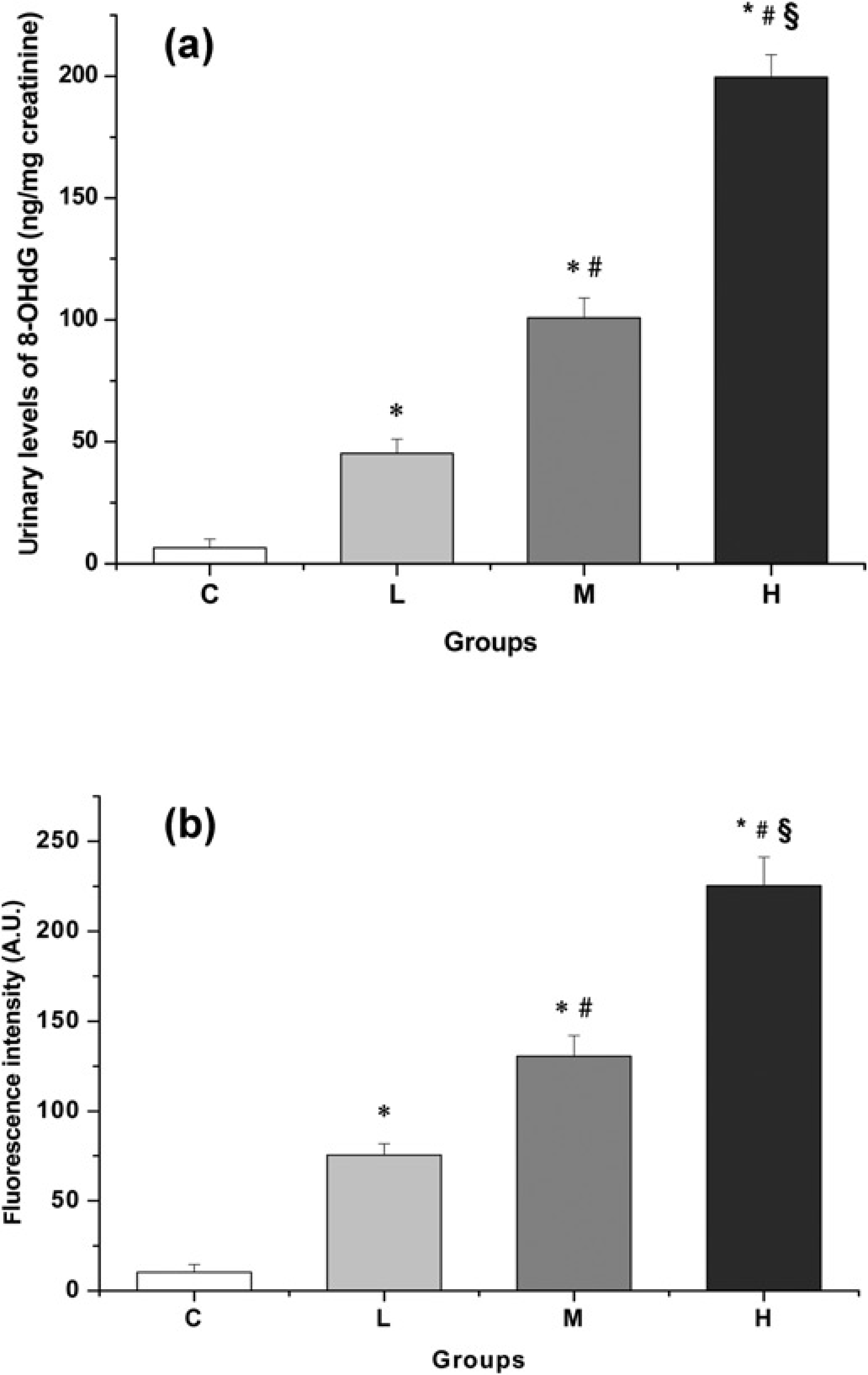
Effect of AIA/DDC treatment on oxidative stress markers: (a) urinary levels of 8-HO-dG and (b) plasmatic ROS concentrations according to AIA dose. Animals were injected with the same AIA and DDC doses as described in Figure 2. Each bar represents the mean ±
ROS concentrations were determined in plasma and as Figure 3b shows their production was greatly augmented in all treated animals. The remarkable ROS increases observed were AIA dose-dependent. Statistically significant increases of 633%, 1168% and 2087% were obtained for the groups corresponding to low, medium and high AIA doses respectively when compared with the control group. All the differences between groups were statistically significant.
Glutathione metabolizing enzymes: GPx, GR and GST
The activity of hepatic GPx, the enzyme that removes peroxides resulting from free radical action and thus protects tissues from oxidative damage, is described in Figure 4a. In response to the drug treatment, enzyme activity increased gradually with the dose of AIA. The increases attained were 17%, 33% and 46% in groups L, M and H respectively, when compared with group C. GPx activities in these three groups were statistically different from those of group C. Activity values from group H were also significantly different from those of group M.
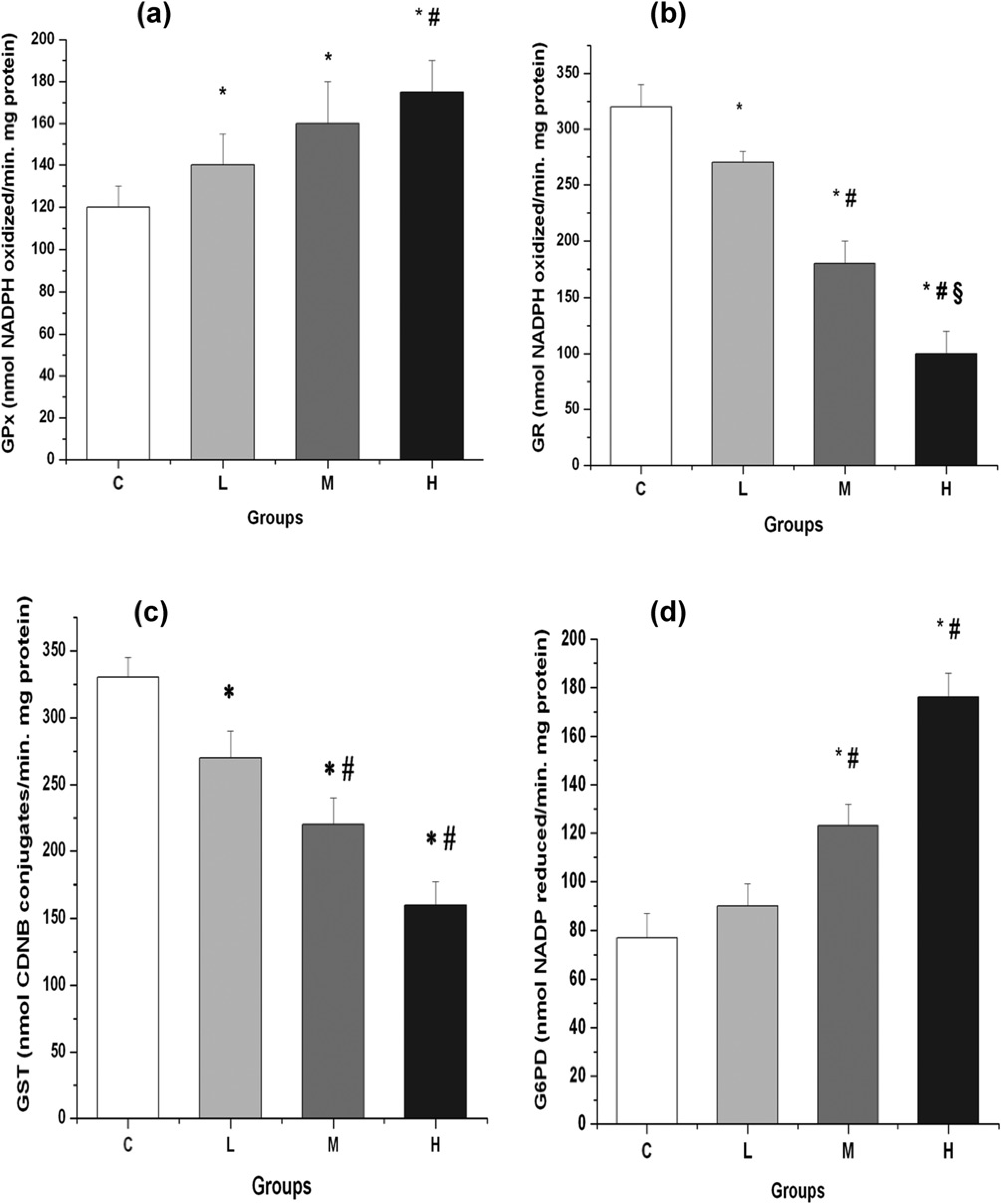
Effect of AIA/DDC treatment on (a) GPx, (b) GR, (c) GST and (d) G6PD activities according to AIA dose. Animals were injected with the same AIA and DDC doses described in Figure 2. Each bar represents the mean ±
Figure 4b shows the activity of GR, a critical enzyme in the maintenance of the cellular redox status. As the AIA dose increased, a significant drop was observed in the activity of this NADPH-reducing enzyme in liver. Levels decreased 16%, 44% and 69% in groups L, M and H, respectively, with respect to Group C. Values observed in groups M and H were significantly different from each other and also significantly different from those of group L.
GST, an enzyme that plays an essential detoxifying role by linking toxic compounds to GSH to form less reactive substances, was assayed in the four groups. Figure 4c shows that the hepatic activity of this transferase decreased as the porphyrinogenic treatment was intensified. Values for groups L, M and H were 18%, 33% and 52%, respectively. Group C (control) was considered as 100%. Values from the three groups were statistically different from those of group C and values from groups M and H were also significantly different from those of group L.
Thiol compounds
Table 1 shows the results obtained when total (protein-bound and non-protein) sulfhydryl groups reacting with Ellmann reagent (T-SH) were estimated. The porphyrino-genic treatment gradually decreased the amount of thiol compounds in liver as AIA dose increased. Thus, levels were reduced 26%, 46% and 55% in groups L, M and H, respectively, when compared with the control. Values observed in groups M and H were significantly different from each other, and also significantly different from those of group L.
Effect of AIA/DDC treatment on thiol profile according to AIA dose
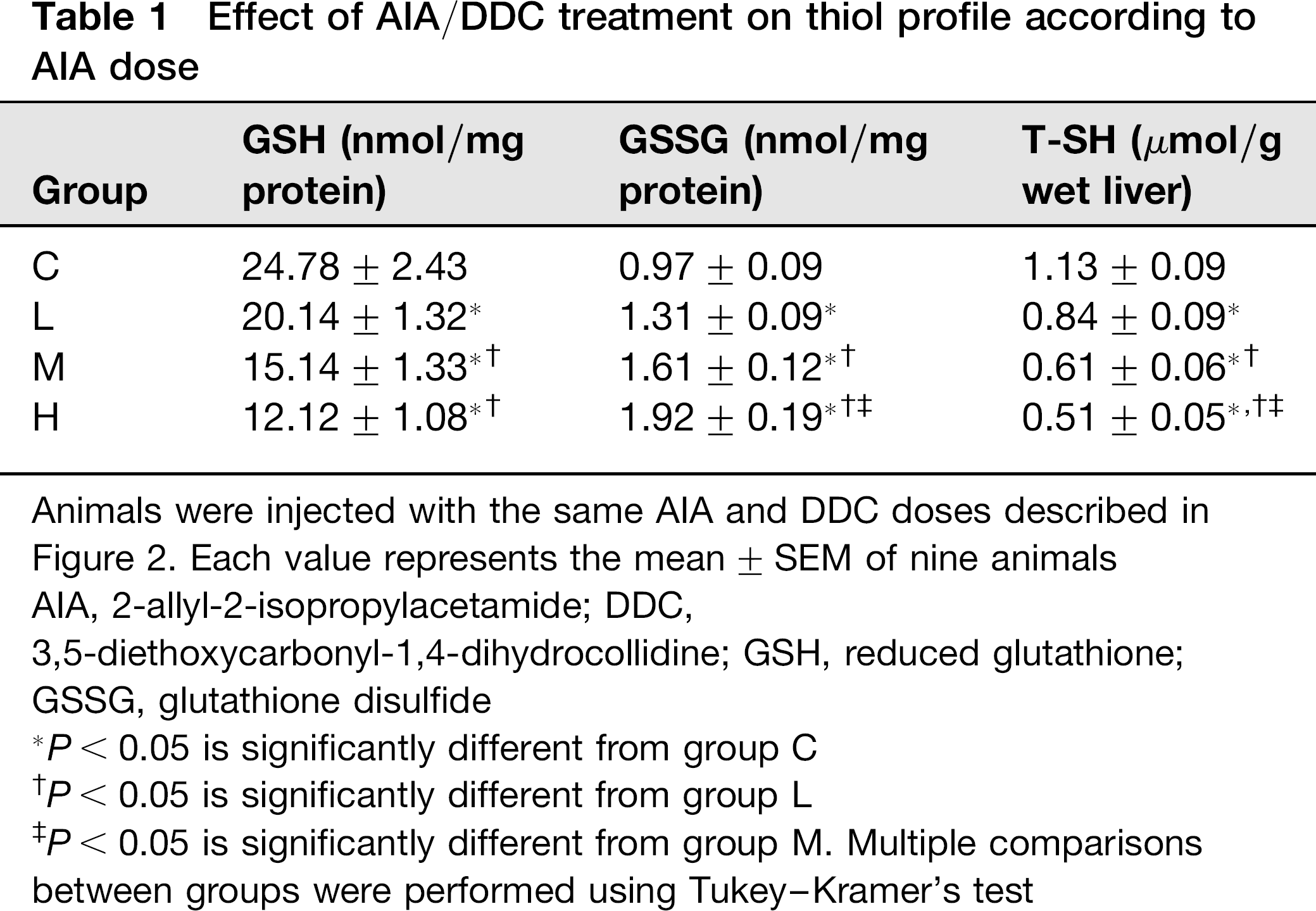
Animals were injected with the same AIA and DDC doses described in Figure 2. Each value represents the mean ± SEM of nine animals AIA, 2-allyl-2-isopropylacetamide; DDC,
3,5-diethoxycarbonyl-1,4-dihydrocollidine; GSH, reduced glutathione; GSSG, glutathione disulfide
P < 0.05 is significantly different from group C
P < 0.05 is significantly different from group L
P < 0.05 is significantly different from group M. Multiple comparisons between groups were performed using Tukey-Kramer's test
Glutathione: reduced and oxidized forms
Levels of hepatic GSH, an endogenous antioxidant produced by the cells, are shown in Table 1. An almost linear loss of this antioxidant agent was observed as AIA doses increased. GSH concentrations were reduced by 19%, 39% and 51% with respect to the control in groups L, M and H, respectively. Values from groups M and H were also significantly different from those of group L.
GSSG concentrations were also measured in the groups studied. GSSG showed a concomitant increase, in agreement with the GSH decrease (Table 1). The amount of hepatic GSSG increased with the AIA dose. GSSG increases in groups L, M and H were 35%, 67% and 100%, respectively, when compared with controls (Table 1). All the differences between groups were statistically significant.
Glucose-6-phosphate dehydrogenase (pentose phosphate pathway)
G6PD, the regulatory enzyme of the pentose phosphate (PP) pathway, was measured in liver with the double purpose of evaluating the consumption of glucose and the formation of NADPH, involved in redox reactions. The activity of this enzyme, responsible for maintaining NAD(P)H pools, increased with the porphyrinogenic treatment (Figure 4d). G6PD increases were 17%, 60% and 129% in groups L, M and H, respectively, when compared with Group C. G6PD activities in the groups M and H were also significantly different from those of group L.
6-phosphogluconate dehydrogenase (6-PGD) (pentose phosphate pathway)
The activity of 6-PGD, the third enzyme of PP pathway and also responsible of generating NADPH, was assayed. The hepatic activity of this enzyme showed dose-dependent and statistically significant increases in all the xeno-biotic-treated groups. Thus, values of 180 ± 28, 230 ± 21, 320 ± 32 (*#) and 402 ± 50 (*#) nmol NADP reduced/ minmg protein (*#) were obtained for group C, L, M and H respectively. These values represent the mean ± SEM of nine animals per group. *P < 0.05 is significantly different from group C; #P < 0.05 is significantly different from group L. Multiple comparisons between groups were performed using Tukey-Kramer's test.
Pyridine nucleotide phosphates: reduced and oxidized forms
As it can be seen in Table 2, the content of NADPH, an important cytoplasmic reducing agent, is increased in liver. Statistically significant increases ranged from 9% (group M) to 14% (group H) when compared with controls. The values of both of these groups also differ statistically from those of the group corresponding to the lower AIA dose, as well as H group does with respect to group M.
Effect of AIA/DDC treatment on NADPH, NADP+ and NADPH/NADP+ according to AIA dose
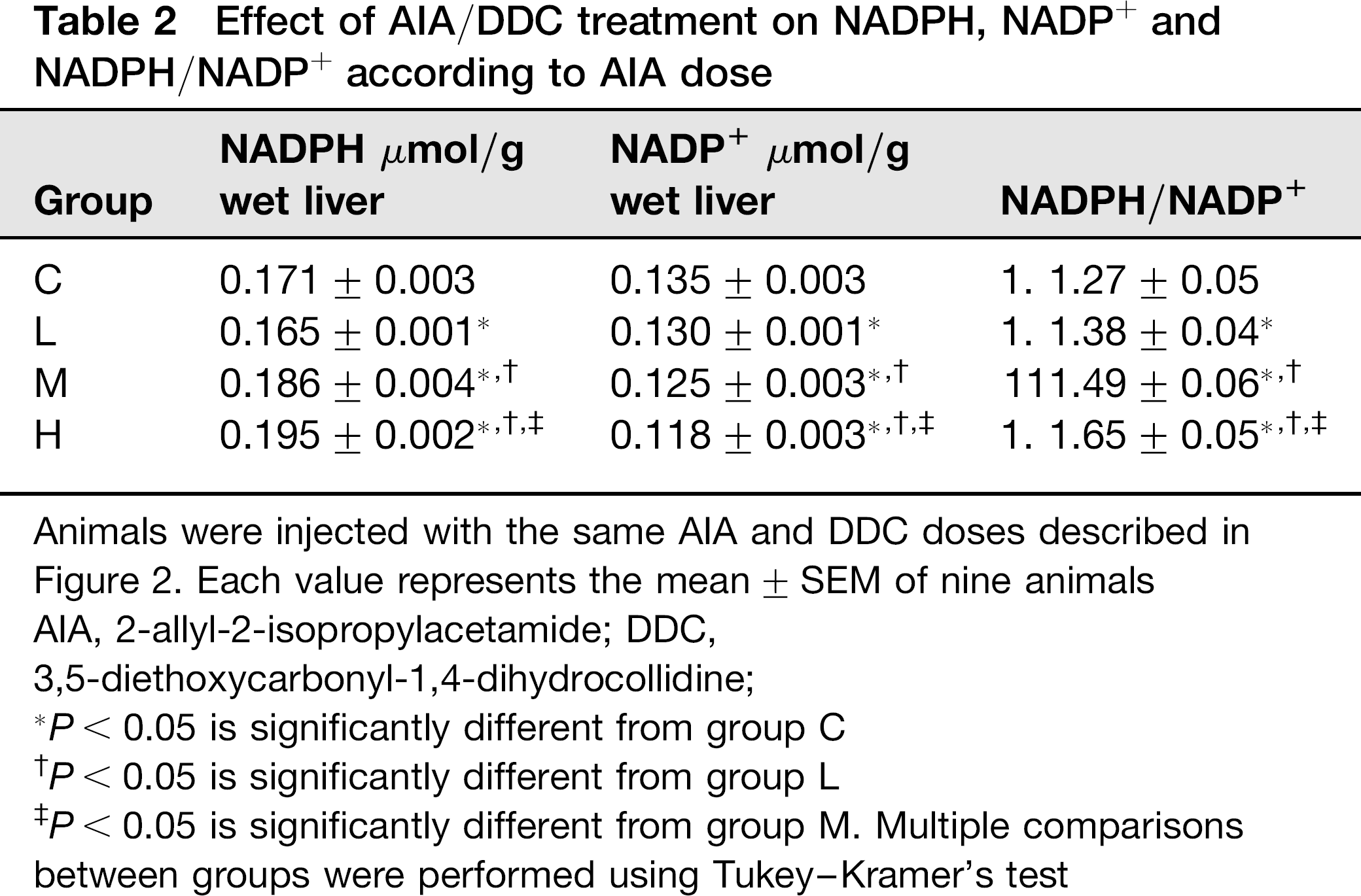
Animals were injected with the same AIA and DDC doses described in Figure 2. Each value represents the mean ± SEM of nine animals
AIA, 2-allyl-2-isopropylacetamide; DDC,
3,5-diethoxycarbonyl-1,4-dihydrocollidine;
P < 0.05 is significantly different from group C
P < 0.05 is significantly different from group L
P < 0.05 is significantly different from group M. Multiple comparisons between groups were performed using Tukey-Kramer's test
A concomitant loss in the levels of NADP+, the product of NADPH oxidation, was observed in livers of xenobiotic-treated rats (Table 2). Thus, statistically significant decreases of 4%, 8% and 13% in NADP+ contents were observed in the groups L, M and H respectively when comparing with controls. Statistically significant differences were also observed between groups.
As shown also in Table 2, the NADPH/NADP+ ratio was enhanced in the three groups of treated rats. The increase in this ratio was AIA-dependent. Thus, the percentages of 8%, 16% and 28% were obtained for groups L, M and H, respectively. All the differences between groups were statistically significant.
Liver glucose concentration
Hepatic glucose concentrations showed dose-dependent and statistically significant decreases in the three groups studied. Values of 7.9 ± 0.1, 7.1 ± 0.2 (*), 6.4 ± 0.2 (*#) and 5.8 ± 0.1 μmol glucose/g wet weight (*#§) were obtained for group C, L, M and H respectively. These values represent the mean ± SEM of nine animals per group. *P < 0.05 is significantly different from group C; #P < 0.05 is significantly different from group L; §P < 0.05 is significantly different from group M. Multiple comparisons between groups were performed using Tukey-Kramer's test.
Discussion
As expected, AIA/DDC treatment impaired the heme pathway promoting the de-regulation of the rate-limiting enzyme ALA-S, the activity of which was enhanced. This up-regulation is due to the action of both drugs: AIA promoting heme destruction and DDC promoting combined heme destruction and heme synthesis inhibition. 3 The increase in ALA production as a consequence of ALA-S de-regulation would lead to oxidative stress. In this respect, it is known that ALA is able to generate free radicals and ROS such as superoxide anion (O2•-), hydrogen peroxide (H2O2), hydroxyl radical (•OH), and ALA enoyl-radical (•ALA), 6 which act as pro-oxidants both in vivo and in vitro. 36 Enhanced DNA oxidative damage and ROS production (Figure 1) confirmed the oxidative stress, indicating that ALA accumulation elicits an oxidative environment. Thus, DNA damage caused by AIA/DDC treatment could be attributed to ROS generation, which was found to be increased in plasma. This damage is in line with what has been reported regarding lipids and protein for these drugs. In this respect, Lelli et al. 4 demonstrated that the activity of the liver antioxidant enzyme SOD was induced by about 235% in AIA/DDC treated rats, whereas CAT activity diminished due to the heme depletion caused by both drugs.
The oxidative environment elicited by the porphyrino-genic AIA/DDC treatment impaired glutathione metabolism resulting in lower GSH and higher GSSG concentrations. Thus, the ratio GSH/GSSG reached almost 4:1 in the fraction containing cytosol and microsomes corresponding to the higher AIA dose. These effects are a consequence of both a decrease in the activity of GR, the enzyme responsible for GSSG reduction to GSH and an increase in the activity of GPx, which consumes GSH and generates GSSG (Figures 4a and b). In these oxidative conditions, GPx uses GSH to eliminate peroxides, so protecting the tissue against the aggression of the xenobiotics, but GSH cannot be adequately replaced due to an inefficient GR activity (Figure 1).
The decrease in GR activity might be attributed to the action of ROS, which would impair the enzyme's redox active site consisting of the dithiol Cys-58-Cys-63, 37 in agreement with reports showing that oxygen radical generating systems irreversibly inhibit GR. 38
The GST activity measured in the present work corresponds to microsomal (MGST) and cytosolic isoenzymes of GST, considering the cellular fraction used for the assays. MGST has a well known role in electrophile conjugation and oxidative stress protection, and a recently reported role in cell protection against cytostatic drugs. 39 On the whole, GST and GSH have a detoxifying action. Results show that there was a decrease in GST activity (Figure 4c). The possible causes for this decrease are: (1) the presence of porphyrins, and particularly of an increased protoporphyrin which is related to hematin, a well-known GST inhibitor, 17 and/or (2) the action of DDC, a methyl-containing compound which can methylate the pyrrolic nitrogen of His present in the GST active site 40 similarly to the methylation of the pyrrolic nitrogen of CYP proto to form [N-methyl-protoporphyrin IX]. 2 In fact, it has been reported that the enzymatic methylation of GST by methyl transferases inhibits GST activity. 41 Likewise, AIA, an allyl-containing acetamide compound, could alkylate the sulfhydryl group of Cys or the pyrrole nitrogen of His, both being amino acids of the active site of GST, 40 similarly to the alkylation of the pyrrole nitrogen of CYP proto to form [N-alkyl-protoporphyrin IX]. 2 In both cases, this would lead to a reduced enzyme activity (Figure 1). Regarding the decrease in GST activity, a similar effect has been reported for other xenobiotics such as chloroform, bromobenzene, carbon tetrachloride and N-nitrosodimethylamine, which caused a marked decrease in hepatic GST and a concomitant increase in the serum enzyme released from liver. 18 GST inhibition is important from a therapeutic point of view, since this enzyme seems to be involved in drug resistance and in the biosynthesis of a number of important arachidonic acid metabolites. As van Bladeren and van Ommen 42 proposed that an inhibition of this enzyme by xenobiotics has toxic consequences by making cells more vulnerable to alkylating agents. 42 Treatment with Sedormid (allylisopropylacetyl carbamide), an structural analogue of AIA which has been shown to cause similar chemical porphyrinogenesis, results in ataxia, paralysis, tremor and neurological symptoms.43,44 Since GST is a key enzyme in chemical defense, 15 the drastic 50% fall in its activity caused by the present xeno-biotic treatment together with the accumulation of ALA 45 would account for the expression of the toxic symptoms of this hepatic disease mentioned above. The decreased activity of GST in this model of hepatic acute porphyria is in line with the observation that alterations in GST expression are implicated in hepatic, cardiac and neurological disease. 46 The increase in G6PD activity observed in the present model is in line with what occurs in a chronic porphyria model 47 and in human hepatic porphyrias, where 6-PGD activity was also found to be enhanced.48,49 This reported stimulation of 6-PGD also agree with the present findings. G6PD is the regulatory enzyme of the pentose pathway. Its main function is the production of NADPH, which plays a central role in cellular ROS metabolism because it is the obligatory reduced equivalent for all major ROS-producing and detoxifying pathways. 12 Moreover, G6PD function is essential in the defense against oxidative stress-dependent NADPH. 12 The increase in NADPH, the decrease in NADP+ and the consequent rise in NADPH/NADP+ ratio here found to agree with the enhanced G6PD activity observed. The increase in hepatic G6PD activity and a more active PP-pathway lead to a decrease in liver glucose, the primary substrate of this carbohydrate pathway, as here reported in AIA/DDC treated rats (Figure 1). Taking into account that glucose represses the heme pathway regulatory enzyme ALA-S, 9 which is induced in porphyrias 1 as shown in the present work, the effect of AIA/DDC treatment on the pentose pathway through G6PD modulation would modulate the porphyria, causing ALA-S de-repression.
Regarding the PP-pathway and porphyria, it is worth mentioning that erythrocyte transketolase, the enzyme that converts ribose-5P into sedoheptulose-7P and glyceralde-hyde -3P, has been found to be decreased in hepatic por-phyrias and mainly in the acute intermitent disease. 50 Considering that PP-pathway is more active in the AIA/ DDC model of acute porphyria, a possible partial block at transketolase concentration would direct the ribose-5P flux toward the synthesis of nucleotides rather than to the glyco-lytic metabolization of the pentose.
In conclusion, our results show that the disruption in GSH metabolism and the PP-pathway is related to the ROS production elicited by the porphyrinogenic AIA/ DDC treatment. These observations have not been previously reported. The disruption involves the impairment of GPx-, GR- and G6PD-coupled oxido-reduction reactions, particularly at GR concentration, which in turn leads to a four-fold lower GSH/GSSG ratio. The stimulation of G6PD, the PP rate-limiting enzyme, would exacerbate the porphyria, since the consequent decrease in glucose would exert a de-repressor effect on the heme pathway regulatory enzyme ALA-S. The glucose depletion-induced effect on this key enzyme would be added to the de-regulatory effect of heme depletion during AIA/DDC treatment (Figure 1), exacerbating the porphyria. The exogenous administration of glucose for the treatment of acute por-phyria would counterbalance the de-regulations of the heme pathway promoted by the higher degradation of glucose into pentoses here reported.
All these results show the high metabolic interplay between the pathways analyzed. So they are a very important contribution to achieve a better treatment for acute human porphyria.
Footnotes
Acknowledgements
This work was supported by grants from the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) and the Universidad de Buenos Aires (UBACyT). LSMV is a Scientific Research Career Member of the CONICET.