
Abstract
Since its discovery in 2000, WW domain-containing oxidoreductase (WWOX, FOR or WOX1) has been considered as a tumor suppressor protein. Global research focus has been aimed mainly toward this direction. In this thematic issue, updated information has been collected regarding the structure, function and signaling of WWOX, along with its critical role as a tumor suppressor and participation in metabolism, neurodegeneration, ataxia, epilepsy, neural disorders, neuronal damages, and interactions with oncogenic viruses. WWOX is not a driver of cancer initiation. Chromosomal alterations in the WWOX gene enhance cancer progression. Importantly, a homozygous nonsense mutation of WWOX gene in humans leads to neural pathologies and early death, rather than spontaneous cancer development. These findings suggest new physiological functions of WWOX in metabolism and neural diseases, and these areas require further investigation.
Specific contents
In the cancer area, there are eight major articles, including contributors from the groups of David Smith, Kay Huebner, Andrzej Bednarek, Yu-Jen Chen, Peijun Liu, and Yao Chang. Essentially, every article provides a comprehensive review on the role of WWOX in cancer. In particular, Yao Chang has discussed the interaction of WWOX with proteins of oncogenic viruses. Also, Robert Richards and Rami Aqeilan brought up the critical issue of WWOX in cancer and metabolism. Ora Schueler-Furman, Amjad Farooq, and Yosef Shaul discussed the tandem WW domain repeats and associated functions, protein/protein interactions, and protein tyrosine phosphorylation in the WW domain areas. Li-Jin Hsu reviewed the role of WWOX in signaling, apoptosis, autophagy, and stress response. Yu-Jen Chen has shown the participation of WWOX in the Sonic hedgehog signaling. Amal Hashem reviewed the emerging role of WWOX in neural diseases. Finally, Pei-Jung Lu discussed the single WW domain-protein Pin1 in the regulation of the cell cycle.
Brief history
This section will be written in lay language to refresh my memory. It has been 15 years since the groups of CM Aldaz and Robert Richards and my group had independently discovered WWOX (also named FOR and WOX1) in 2000.1–3 My team isolated and functionally characterized the mouse Wwox cDNA at the Guthrie Research Institute. The Institute is located in a very small, deserted railroad town called Sayre in PA. Shortly after our isolation and characterization of Wwox, we had surprising visitors, including Robert Richards from the University of Adelaide, Adam Paige from the Imperial Cancer Research Fund UK, David Smith from the Mayo Clinic, and Udo Oppermann from the Karolinska Institute. A surprising wakeup call to me was from Kay Huebner of the Ohio State University right after my sinus surgery in 2002. In 2005, Marius Sudol, the father of WW domain, relocated to our neighborhood at the Geisinger Clinic in Danville, PA. It was then a great touch with the real WW domains. And, again in 2014, we had another E.T.-style touch from Taiwan via the WW domains with Marius Sudol at Singapore. To us, we cherished the sunshine from a world out there. I recalled numerous pleasant conversations and discussions with many friends and competitors in the field. Now, WWOX is a real “Wild Wild OX”.
In 2002, David Smith isolated the mouse Wwox genome. 4 This has paved the road to make gene knockout mice. Meanwhile, under the encouragement of my most enthusiastic classmates Su (Shur-Tzu) Chen in Tainan and Chun-I Sze in Denver, we together first explored the role of Wwox in the mouse brain development, stress response, and neurological diseases such as Alzheimer’s disease starting in 2001.5–7 In addition, Feng-Jie Lai and Hamm-Ming Sheu had first pioneered the role of WWOX in the skin cancer development both in mice and humans. 8 Importantly, outstanding cancer researchers Carlo Croce, Kay Huebner, Rami Aqeilan, CM Aldaz, and many global investigators have greatly contributed to establish the WWOX field in cancer, especially in the areas of gene alterations, epigenetic modifications and relationship with cancer development. The updated knowledge about WWOX in cancer has been well documented in this thematic issue. Furthermore, recent development of Wwox knockout mice, along with studies in genetic linkage in humans, has led to the discovery that WWOX participates in metabolism, such as in the regulation of the metabolism of steroid, high-density lipoprotein cholesterol, glucose, and bone homeostasis. Most recently, WWOX was shown to play a role in the regulation viral proliferation, suggesting that WWOX participates in the activation of innate immune system against invading microorganisms.
WWOX in bubbling death
Over the years, my group has been pioneering the mechanisms of WWOX signaling pathways under various stress stimuli, including transforming growth factor beta (TGF-β), tumor necrosis factor (TNF), hyaluronan/hyaluronidase Hyal-2, complement C1q, UV irradiation/cold shock, chemotherapeutic drugs, and p53.1,8–13 In our most recent study, we have reported temperature-regulated “bubbling death” and its transition with apoptosis. 12 Bubbling death is defined as “formation of a bubble from the nucleus and release of this swelling bubble to the cell surface that causes cell death”. Namely, cells are subjected to UVC irradiation and/or brief cold shock and then incubated at room temperature or 4°C. Each cell releases an enlarging nuclear gas bubble containing nitric oxide (NO). Brief cold shock was shown to enhance the bubble formation. 12 Many types of cells also release exosome-like particles to the extracellular space during bubbling death. Unlike typical apoptosis, there are no flip-over of phosphatidylserine (PS) onto cell surface, mitochondrial apoptosis, and chromosomal DNA fragmentation in the UV/cold shock-treated cells. 12 In contrast, when cell is exposed to UV/cold shock and incubated at 37°C, bubble formation is significantly retarded and DNA fragmentation occurs. 12 The observation suggests that bubbling death is switched to apoptosis under increasing temperatures. Unlike membrane blebbing, bubbling is irreversible and occurs prior to uptake of propidium iodide by the nucleus. That is, bubbling is essential in leading to ultimate nuclear damage and death. 12
In the absence of WWOX, knockout Wwox−/− MEF fibroblasts resist UV/cold shock-induced bubbling death. Also, in the absence of WWOX or the presence of a dominant-negative p53, bubble formation is suppressed. We have demonstrated that both WWOX and p53 function in a synergistic manner in causing apoptosis under various stress conditions.8–12 Our supporting evidence revealed that NO, nitric oxide synthase 2 (NOS2), p53 and WWOX enhance in the bubbling death, whereas antiapoptotic TRAF2 blocks the cell death.
12
Importantly, nuclear accumulation of p53 and WWOX is needed to activate NOS2 to induce formation of NO. Whether WWOX and p53 bind and counteract the protective function of TRAF2 is unknown and remains to be established (Figure 1).
A schematic model for UV/cold shock-induced signaling event. The graph shows UV/cold shock-induced relocation of the potential complex of p53, TRAF2 and WWOX to the nucleus, wherein p53 and WWOX are activated. NOS2 alone, or together with TRAF2, also relocates to the nucleus. Appearance of NO-containing gas is shown in the nucleus. In most cases, the gas is released as a jet stream tunneling through the nucleus and cytoplasm to pop out a single large bubble per cell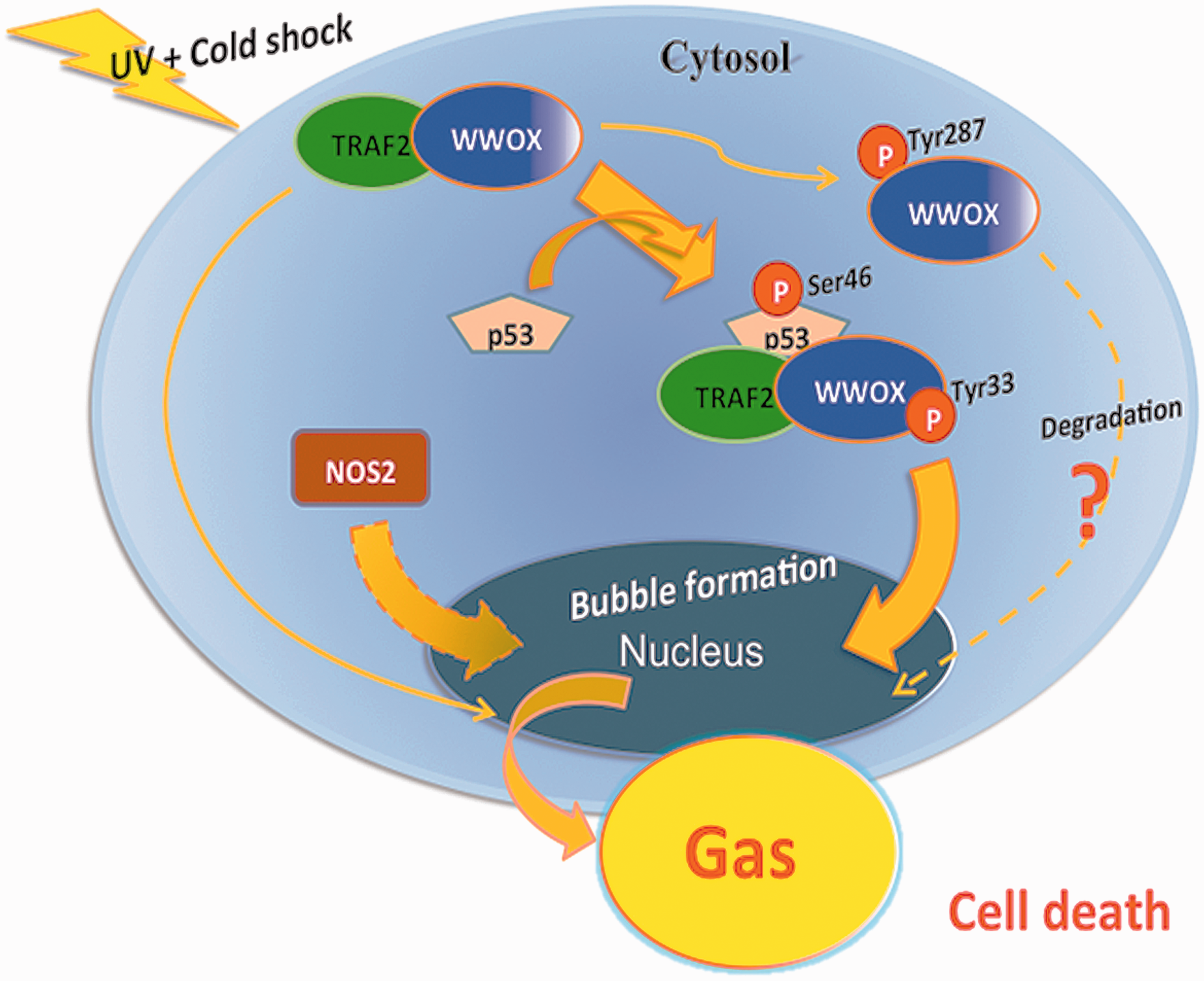
WWOX in neural diseases
Compelling evidence shows that WWOX plays a critical role in the neural development and diseases.5,6,13–21 WWOX levels are upregulated during embryonic brain development. 5 Rapid accumulation of WWOX has been shown in the apoptotic nuclei of damaged neurons when rats are subjected to (1) prolonged exposure of eyes to constant light, 6 (2) exposure to dopaminergic neurotoxin 1-methyl-4-phenyl-pyridinium (MPP+) to develop Parkinson's disease (PD)-like symptoms in the striatum of the brain, 14 or (3) dissection of sciatic nerve. 15 WWOX physically interacts with proteins associated with Alzheimer’s diseases (AD). 13 Proteins of this category include TRAPPC6A, 16 TIAF1,17,18 Tau, 7 GSK-3β, 19 JNK1,7,20 ERK,7,21 and many other unidentified proteins. WWOX is frequently down-regulated in the hippocampi of AD brains.13,16,17 This down-regulation leads to aggregation of TRAPPC6A and TIAF1, followed by caspase activation, APP phosphorylation and degradation and Tau aggregation.13,16,17 Functionally, WWOX inhibits GSK-3β-mediated tau hyperphosphorylation, and thus prevents AD pathogenesis. 19 Also, suppression of GSK-3β by WWOX promotes neuronal differentiation. 19
Finally, WWOX modulates high-density lipoprotein (HDL) and lipid metabolism, including cholesterol homeostasis, ApoA-I/ABCA1 pathway, and fatty acid biosynthesis/triglyceride metabolism. 13 Alteration of lipid metabolism contributes to the pathogenesis of neurological diseases. Large consanguineous family studies have determined that homozygous WWOX point mutations (e.g. P47T and G372R) lead to a new form of childhood onset autosomal recessive cerebellar ataxia and epilepsy. 22 Homozygous nonsense mutation of WWOX results in protein loss, and the patient suffers from severe anomalies, including short and growth retardation, microcephaly with seizure, retinal degeneration, and early death at 16 months of age. 23 In Ide/Ide rat, the Wwox gene has been shown to possess a 13-bp deletion in exon 9 that causes the rats to suffer dwarfism, postnatal lethality, male hypogonadism, and a high incidence of epilepsy and many vacuoles in the hippocampus and amygdala. 24 Under the loss of WWOX protein, no spontaneous tumor formation was shown. In stark contrast, functional deficiency of WWOX in Drosophila does not generate the disease phenotypes in the neural system. 25
Conclusion and perspectives
Taken together, accumulative evidence supports the notion that WWOX is a protein of multiple functions. Under a physiological setting, WWOX probably interacts with many proteins in different pathways at different locations in the cell membrane, cytoplasm, and nucleus. Outstanding questions still to be resolved are: (1) WWOX is considered as a bona fide tumor suppressor. 26 However, a homozygous nonsense mutation of WWOX causes neural disorders and early death but without spontaneous tumor formation in humans. 23 Similar observation is shown in the Ide/Ide rat. 24 The underlying mechanisms remain to be established. (2) WWOX binds viral proteins and is likely to control viral proliferation. NF-κB appears to provide a mechanism to suppress WWOX. NF-κB suppresses the expression of WWOX.27,28 However, transiently overexpressed WW domain induces activation of NF-κB. 15 WWOX modulates the function of transcription factors such as CREB, CRE, c-Jun, Elk-1 and AP-1. 15 Viruses could have utilized the cellular machinery to enhance survival. Whether WWOX regulates the immune cell functions is unknown. (3) The C-terminal SDR domain of WWOX is frequently altered. This domain is responsive to stimulation by sex steroid hormones such as androgen and estrogen. The underlying signaling and physiological relevance of hormone-regulated WWOX signaling need further investigation.
Footnotes
ACKNOWLEDGEMENTS
I would like to thank Dr. Steven Goodman, Editor-in-Chief, and the Editorial Board of Experimental Biology and Medicine for approving the 2015 Thematic Issue for WWOX. I gratefully appreciate the outstanding efforts of Dr. Rami Aqeilan of the Hebrew University, Israel, and Dr. Chun-I Sze of the National Cheng Kung University, Taiwan, in making the thematic issue possible. Finally, I would express my sincere thanks to all the contributing authors. Your dedication to your article is absolutely outstanding. Without doubt, the whole collection of articles would dramatically enhance our understanding of the biology of WWOX and provide insights to bring up a bigger picture of WWOX in the near future.
This research was supported, in part, by the Department of Defense USA (W81XWH-08-1-0682), the Ministry of Science and Technology, Taiwan, ROC (NSC98-2628-B-006-041-MY3, 99-2320-B-006-012-MY3, 102-2320-B-006-018-, 102-2320-B-006-030-, and 102-3011-P-006-005-), the National Health Research Institute, Taiwan, ROC (NHRI-EX101-10102BI, and NHRI-EX102-10102BI), the National Cheng Kung University Landmark Projects (C0167) and the Department of Health, Taiwan, ROC (DOH101-TD-PB-111-TM010) (to NS Chang).