
Abstract
A major characteristic of immunodeficiency associated with life-threatening intracellular infection in adults is the presence of anti-interferon-γ antibodies. Although little is known about the mechanism underlying this syndrome, it is believed that the antibodies inhibit the activity of downstream signaling pathway of interferon-γ. In this study, the characteristics of these antibodies in patients who presented, or have a history of, intracellular infection and were positive to anti-interferon-γ antibodies were investigated. The antibodies exhibited mainly the IgG1 and the IgG4 subtypes and recognized the C-terminal of the interferon-γ linear epitope containing the KRKR motif, which is required for the biological activity of interferon-γ. The antibodies bound to recombinant interferon-γ with significantly lower avidity than antibodies to a recall antigen, tetanus toxoid, suggesting that the antibodies might have not undergone affinity maturation. The data from this study may provide fundamental information to better understand the properties of anti-interferon-γ antibodies, which can be useful for future studies.
Impact statement
An increase in the number of immunodeficient patients related to autoantibodies to interferon (IFN)-γ has been observed particularly in East Asian adults. These patients are often presented with opportunistic infections caused by intracellular pathogens, including non-tuberculous mycobacteria (NTM), Cryptococcus neoformans, Penicillium marneffei (now called Talaromyces marneffei), and non-typhoidal Salmonella spp. The mortality rate for this syndrome is relatively high with 32% patients dying at the median time of 25 months after diagnosis. Characterization of these autoantibodies may promote better understanding of the syndrome, an emerging health problem affecting East Asia populations and impeding their welfare and economic development.
Keywords
Introduction
Adult onset immunodeficiency related to the presence of anti-interferon (IFN)-γ antibodies in patients presenting with non-tuberculous mycobacterial (NTM) and other intracellular infections has been reported mostly in Asian populations.1–6 This syndrome has also been reported in Caucasian and African individuals living in different settings around the world.7–9 The biological mechanism underlying the pathogenesis of these antibodies remains to be elucidated. Several studies have shown that anti-IFN-γ antibodies have inhibitory activity on signal transduction of IFN-γ and the sequentially responsive molecules of IFN-γ.2,3,9
Effect or functions of antibodies are mostly mediated by the heavy chain constant regions of the immunoglobulin (Ig) molecule. In humans, the five major classes or isotypes of antibodies are IgM, IgD, IgG, IgA, and IgE, of which IgG is the most abundant immunoglobulin and it can be divided into four subtypes: IgG1, IgG2, IgG3, and IgG4. 10 Distinct hinge regions and amino acid sequences in the fragment crystallizable (Fc) part determine the different biological and immunological properties of the antibody molecule, such as complement activation and binding to cellular molecules to mediate effector functions. Diverse antibody isotypes and subtypes may, therefore, exhibit different immune functions.
The variable region of an antibody recognizes an antigenic determinant or an epitope on its target. The epitope can be divided into conformational or discontinuous epitope composed of amino acids from different parts of the protein which are brought together by protein folding, and linear or continuous epitope which is a single fragment of continuous amino acids. After binding to a specific antigen, B cells migrate to secondary lymphoid organs where they improve their antigen-binding affinity. 11 This process is called antibody affinity maturation, and it gives rise to a more specific immune response when re-encountering the antigen. An assay on binding affinity could provide some information on whether the antibodies could have undergone affinity maturation.
Although reports on adult-onset immunodeficiency related to anti-IFN-γ antibodies continue to be on the rise, little is known about the antibody characteristics. In the present study, the classes and subtypes of anti-IFN-γ antibodies were determined, the linear epitopes targeted by the antibodies were identified, and the binding affinity of the antibodies was investigated. The information may be useful for understanding the immune response of the disease.
Materials and methods
Study populations
Sixty-three human immunodeficiency virus (HIV) negative cases having records of proven opportunistic infections and anti-IFN-γ antibodies 4 that had been followed up at Chiang Mai University Hospital, Chiang Mai, Thailand, were enrolled in this cross-sectional study. There were 32 male participants aged 39–77 years and 31 female participants aged 41–71 years in this study. These patients experienced at least one episode of NTM, cryptococcosis, histoplasmosis, salmonellosis, penicilliosis, cytomegalovirus infection, or herpes zoster infection, etc. Twenty female and 10 male healthy controls aged 39–61 years without a history of infections in the past month or a history of Mycobacterium tuberculosis infection were enrolled for comparison. No participants were receiving immunomodulating drugs.
The study was approved by the ethics committees of the Faculty of Medicine (105/2557) and the Research Institute for Health Sciences (13/56), Chiang Mai University. All procedures performed in studies involving human participants were in accordance with the ethical standards and with the 1964 Helsinki declaration. Written informed consent was obtained from all individual participants before enrollment into the study.
Determination of anti-IFN-γ antibody by ELISA
The anti-IFN-γ antibody in the sera was determined by indirect enzyme-linked immunosorbent assay (ELISA), as previously described. 4 Pooled positive sera from patients confirmed positive to anti-IFN-γ antibodies and presented with at least two episodes of culture or histopathologically proven intracellular microorganism infection 4 given an arbitrary unit value for 1000 units/ml were serially diluted on each plate as a standard curve. The antibody levels were then determined by calculation against the standard curve. 12 The negative–positive cutoff value was assigned as the average anti-IFN-γ antibody level in the healthy controls plus three standard deviations. Recombinant IFN-γ used throughout this study was purchased from R&D systems (Minneapolis, MN).
Determination of anti-IFN-γ antibody isotypes and subtypes
Determination of anti-IFN-γ antibody isotypes and subclasses was carried out using the ELISA protocol. After the samples were added to all the wells, and the washing step removed the excess components, HRP-conjugated anti-human IgG1, IgG2, IgG3, IgG4, IgA, and IgM (Invitrogen) were added and the reaction of the enzyme-substrate was read by spectrophotometry. The samples were analyzed in duplicates.
Determination of linear B cell epitopes of IFN-γ
Linear B cell epitopes of IFN-γ were identified by using a library of overlapping synthetic peptides spanning its sequence. 13 The amino acid sequence of IFN-γ was searched on the NCBI website using the basic alignment search tool (BLAST). Biotinylated peptides with 20 amino acids in length, overlapping each other by 10 amino acids (Figure 1) were produced at VCPBIO limited, Shenzhen, China. The purity of the peptides was greater than 95%.

Amino acid sequences of peptides corresponding to IFN-γ. The peptides were 20 amino acids in length and overlapped each other by 10 amino acids.
Immunoplates were coated overnight with 2 μg/ml streptavidin. After washing, the plates were blocked and the biotinylated peptides were added to each well. After incubation with the sera, anti-human IgG conjugated to HRP was added, which was followed by the addition of the OPD substrate. The enzyme reaction was read by spectrophotometry.
Avidity assay
ELISA for the determination of anti-IFN-γ antibodies was carried out as described above. Following the incubation step of the sera, one duplicate set of wells was treated with 4.0 M guanidine hydrochloride (Sigma) dissociating solution in 0.05% Tween-20/PBS for 10 min prior to washing with PBS-T. For tetanus toxoid (TT), plates were coated with 1:500 of TT (Serum Institute of India), which was followed by the protocol described above. The avidity indices were calculated as the ratio of the OD of the guanidine-treated wells to the OD of the untreated wells, multiplied by 100.
Statistical analysis
Mann–Whitney U test was used to analyze the differences in the parameters between groups. All the analyses were performed using the GraphPad Prism Software.
Results
High levels of anti-IFN-γ antibodies in cases
The levels of anti-IFN-γ antibodies in cases detected by ELISA were statistically significantly higher than that of the healthy controls (P < 0.001) (Figure 2(a)). Upon using the cutoff value as the average antibody levels of the controls plus 3SD, all the cases were found positive to anti-IFN-γ antibodies. The median anti-IFN-γ antibody levels in the cases were 105.6 (interquartile range (IQR) 18.7, 677.8) and the same in the healthy controls was 1.7 arbitrary units (IQR 1.0, 2.3).
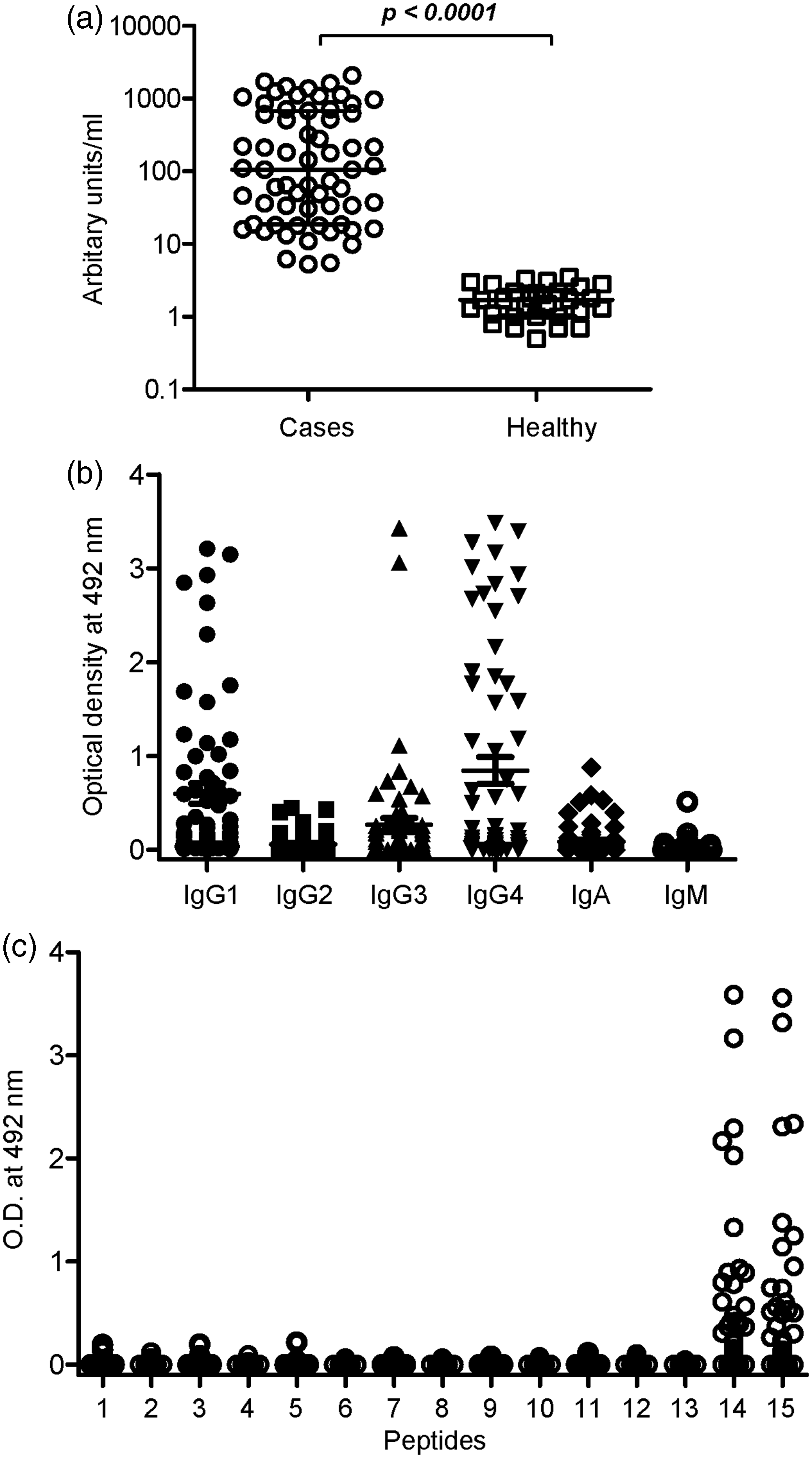
Levels (a), isotypes and subtypes (b), and linear B cell epitope mapping (c) of anti-IFN-γ antibodies. (a) Anti-IFN-γ antibody levels in the cases and the healthy controls were determined by indirect ELISA. (b) Isotypes and subtypes of anti-IFN-γ antibodies in the cases. The data show the average OD value of duplicate wells coated with IFN-γ subtracted by the average OD of antigen-free wells. (c) Identification of B cell epitopes on IFN-γ by ELISA. Each symbol represents one individual.
Anti-IFN-γ antibodies were of IgG1 and IgG4 subtypes
Anti-IFN-γ antibody subtypes in the cases were predominantly IgG1 and IgG4 (Figure 2(b)). The ODs presented in the figure were calculated by subtracting the average OD value of duplicate wells coated with the antigen by the average OD of antigen-free wells. Of the 63 cases, 38 (60.3%), 14 (22.2%), 29 (46.0%), 35 (55.6%), 15 (23.8%), and 3 (4.8%) gave positive results to IgG1, IgG2, IgG3, IgG4, IgA, and IgM, respectively. Different cases showed heterogeneity in the antibody response to IFN-γ, in which some cases had all the antibody subtypes and also IgA and IgM isotypes, whereas some cases had a single isotype (Table 1).
Patterns of anti-IFN-γ antibody isotypes and subtypes in cases (the gray boxes indicate positive response to anti-IFN-γ antibodies).
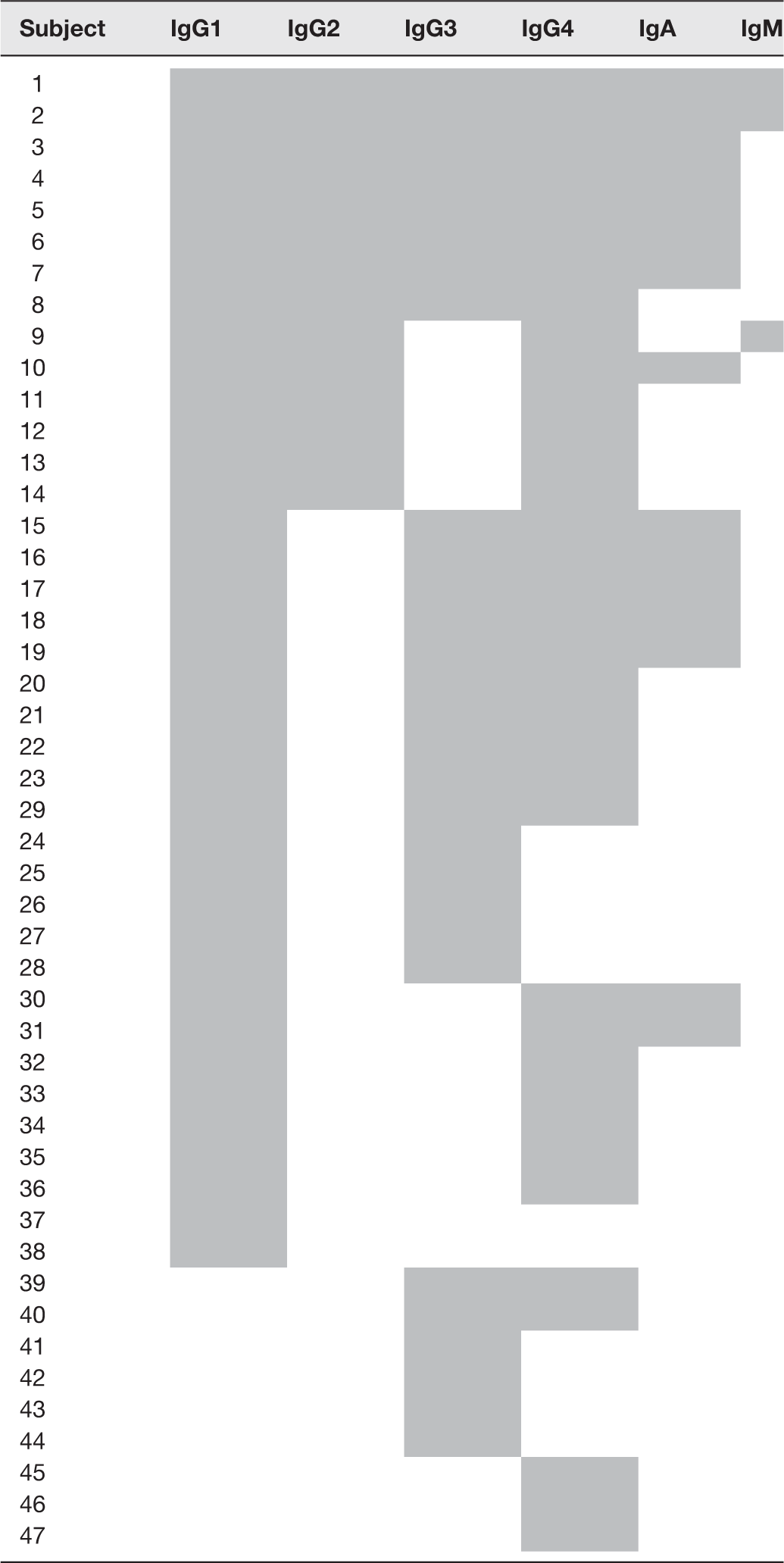
IFN: interferon.
A linear B cell epitope was recognized by sera from cases
To identify the B cell epitope which binds to anti-IFN-γ antibodies, sera from the cases were analyzed against synthetic peptides corresponding to IFN-γ by ELISA, as described in the section “Materials and Methods.” Of the 63 cases, peptides 14 and 15 were recognized by 27 (42.9%) and 25 (39.7%) cases, respectively (Figure 2(c)). Among the 27 cases that gave positive results to peptide 14, 25 of them also reacted to peptide 15 and only two of them reacted to peptide 14 alone.
Anti-IFN-γ antibodies exhibited lower avidity indices than anti-TT antibodies
To assess the avidity of anti-IFN-γ antibodies, sera from 46 cases that gave positive results to both IFN-γ and TT were tested for avidity indices with regard to both the antigens. TT was chosen as a recall antigen because TT vaccine is routinely given to Thai children and to adults as booster shots. As shown in Figure 3(a), anti-IFN-γ antibodies in cases bound to IFN-γ with statistically significant lower avidity when compared to anti-TT antibodies. The avidity of antibodies to TT in the cases was not different from that in the healthy controls (Figure 3(b)). The avidity of antibodies to IFN-γ between cases that gave positive and negative results to peptide 14 and 15 was not statistically significant (data not shown).
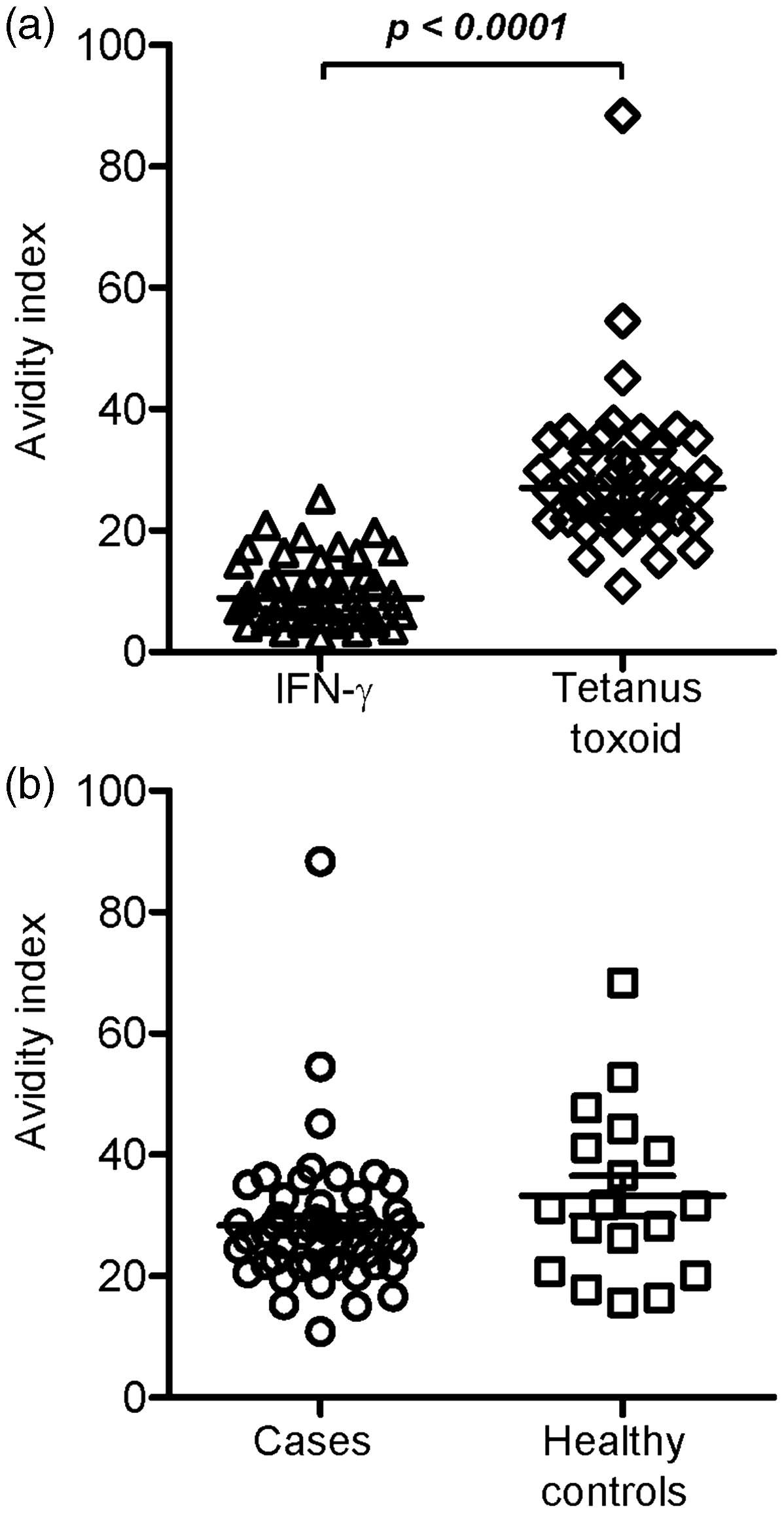
Avidity indices of antibodies. (a) Avidity indices of antibodies specific to IFN-γ and TT in 46 cases that gave positive results to both the antigens. (b) Comparison of avidity indices of antibodies against TT between the cases and the healthy controls.
Discussion
This report describes the characteristic of anti-IFN-γ antibodies. It was found that IgG1 and IgG4 were the most prominent subtypes and that the antibody patterns were heterogeneous among cases. The antibodies recognized the C-terminal of the IFN-γ amino acid sequences. Anti-IFN-γ antibodies were bound to the protein with significantly lower avidity when compared to recall antigen TT.
Isotypes and subtypes of anti-IFN-γ antibodies appear to be heterogeneous. Previous reports have shown different isotypes and subtypes in patients with the same syndrome.7–9 However, Browne et al. 3 have reported a predominance of the IgG4 subtype in studies on larger populations. 3 In this study, it was found that IgG1 and IgG4 were the most frequent subtypes in the population. Moreover, multiple isotypes and subtypes were detected in some cases (Table 1). This implies that anti-IFN-γ antibodies-secreting B cells have undergone antibody class switching. In general, IgG4 comprises approximately 5% of the total IgG and elevated IgG4 levels in the serum, which have been reported in several chronic diseases. 14 Whether the pronounced response of IgG1 and IgG4 against IFN-γ is the contributing factor, as observed in this study, or it is that particular subtypes might be contributing to the progress of individual diseases is under a longitudinal investigation.
Identification of the B cell epitopes that anti-IFN-γ antibodies bind to may provide potential therapeutic or diagnostic targets. A study by Lin et al. 15 using 30-mer overlapping peptides which correspond to the IFN-γ amino acid sequence showed that anti-IFN-γ antibodies recognize a C-terminal region in the IFN-γ protein. Upon using 20-mer peptides, it was also found in this study that anti-IFN-γ antibodies recognize peptide 14 and peptide 15 which contained the conserved KRKR motif, which is required for the biological activity of IFN-γ. 16 Synthetic peptides sharing homology with Aspergillus spp. Noc 2 proteins containing the KRKR motif induced the antibodies capable of neutralizing human IFN-γ in rats. Therefore, molecular mimicry to Aspergillus spp. Noc 2 protein was proposed as the mechanism that induces anti-IFN-γ antibody responses in these patients. 15 It is likely that anti-IFN-γ antibodies in the population of this study did not recognize the SPAAKTG sequence reported previously 15 as reflected by the negative result on peptide 13 which shared the same SPAAKTG sequence with peptide 14. However, substitution of the SPAAKTG motif by the mouse corresponding sequence did not completely abolish the binding capacity of the anti-IFN-γ antibodies 15 ; this suggests that the KRKR motif may be the major target of cross-reactive antibodies.
In contrast to other multiple anti-cytokine antibodies, 17 in this study, it was found that anti-IFN-γ antibodies bound to IFN-γ with significantly lower affinity when compared to anti-TT antibodies. This suggests that anti-IFN-γ antibodies might not have undergone affinity maturation. In general, when antigen-specific B cells re-encounter the antigen, they enter the germinal center in secondary lymphoid organs and receive help from T cells to proliferate and undergo somatic hypermutation of the Ig variable region genes, resulting in the expansion of clones with increased affinity. 18 The binding affinity of anti-TT antibodies was comparable between the cases and the healthy controls, suggesting that the affinity maturation process in the cases was normal. Whether the low binding affinity of the anti-IFN-γ antibodies was due to lack of cognate autoreactive T cell help should be elucidated. In conclusion, the data obtained in this study described the characteristics of anti-IFN-γ antibodies, which might provide useful information for understanding the immune response in the context of the disease or for developing a therapeutic approach.
Footnotes
Authors’ contributions
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript; KC and KR performed the laboratory assay and data analysis, RC, JP and KS provided patient samples, JW designed the study and wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We are grateful to all the participants, to Narumon Tachawong for her technical assistance, to Hi-Tech Outsourcing Services for the editorial assistance, and to the staff of the Department of Internal Medicine of the Chiang Mai University Hospital for recruiting the participants.
Declaration Of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study was funded by Chiang Mai University, Chiang Mai, Thailand.