
Abstract
The association between the presence of anti-interferon-γ autoantibodies and the onset of immunodeficiency with intracellular infections has been clearly established. No standard regimen to control the production of these pathogenic autoantibodies, apart from antimicrobial therapy to eliminate infections, contributes to the medical burden of this syndrome, which sometimes has a fatal outcome. In this review, we summarize the findings on anti-interferon-γ autoantibodies to facilitate further research and to provide guidance for treatment strategies.
Keywords
Impact statement
The frequency of various opportunistic infections associated with anti-interferon (IFN)-γ autoantibodies (AAbs) has increased in the past decade. Numerous studies including clinical manifestations, diagnostic methods, mechanisms of pathogenesis and treatment have been revealed. This review provides comprehensive information on anti-IFN-γ AAbs associated with opportunistic infections.
Introduction
The term “adult-onset immunodeficiency” was first coined by Browne et al. to describe an immunodeficiency syndrome related to the presence of anti-interferon (IFN)-γ autoantibodies (AAbs) in adults with multiple opportunistic infections similar to that observed in patients with advanced human immunodeficiency virus (HIV) infection. 1 Cumulative reports have clearly confirmed that anti-IFN-γ AAbs contribute to immunodeficiency that is accompanied by intracellular infections, mostly in adults. Several studies, including ours, have shown relatively high rates of fatality in these patients.2–6 Most reports of adult-onset immunodeficiency were from academic or university hospitals regarding patients who had been referred from smaller medical care institutions.1,7,8 Thus, we believe that the actual number of cases of adult-onset immunodeficiency is grossly underestimated as this emerging syndrome may not be commonly recognized. Thus far, most treatments for adult-onset immunodeficiency focus on antimicrobial therapy to eliminate infections. There is no standard treatment guideline for controlling the production of anti-IFN-γ AAbs. This review discusses our current understanding of anti-IFN-γ AAbs in terms of etiology, pathology, its effect on the immune system, detection, characteristics, and treatment.
Etiology of anti-IFN-γ AAbs
It is believed that anti-IFN-γ AAbs is the cause of immunodeficiency rather than the result of opportunistic infections. A time series analysis in a patient with disseminated Mycobacterium avium complex infection showed that neutralizing anti-IFN-γ AAbs gradually develop for some time before the onset of opportunistic infections. 9 However, the factor that triggers anti-IFN-γ AAbs production is not known. It has been shown that natural antibodies against IFN-γ can be found in infection-free healthy individuals ranging from newborns to adults. 10 Nevertheless, titers of anti-IFN-γ AAbs were significantly higher in patients suffering from various viral infections. These titers declined, approaching normal levels as the infections gradually resolved. The use of Anaferon, an IFN-γ-specific therapeutic antibody for children, in patients with varicella infection resulted in a significant decrease of anti-IFN-γ AAbs compared to patients receiving a placebo. 11 These data suggest that the production of anti-IFN-γ AAbs could be the result of natural immune regulation following infections, but how these antibodies become pathogenic remains unclear.
Molecular mimicry, when the immune system responds to foreign antigens sharing a similar sequence or structure to self-antigens, is proposed as another possibility in the production of anti-IFN-γ AAbs. 8 The sequence of amino acids of the IFN-γ molecule that the autoantibodies bind to shares 100% homology to amino acids of the ribosome assembly protein Noc2 of Aspergillus terreus, which is highly conserved across all of the Aspergillus spp. 8 A synthetic Aspergillus Noc2 peptide was able to bind to anti-IFN-γ AAbs in patients and induce antibodies that cross-reacted with human IFN-γ and inhibit the IFN-γ-stimulated Phosphorylated−Signal Transducer and Activator of Transcription (pSTAT)−1 upregulation. The difference in the neutralizing capacity of anti-IFN-γ AAbs found in patients and non-neutralizing antibodies in healthy populations may explain pathogenicity. This assumption is supported by evidence showing that patients with anti-IFN-γ AAbs without neutralizing capacity did not have opportunistic infections,1,12 suggesting that not only the amount of the antibody but also IFN-γ–blocking activity is required for pathogenicity. A longitudinal study is required to identify risk factors for the onset of the anti-IFN-γ AAbs and immunodeficiency.
Effect of anti-IFN-γ AAbs on its downstream elements
IFN-γ, a central regulator of the immune system critical for controlling intracellular infections, is secreted by activated T cells, natural killer (NK) cells, and macrophages. The binding of homodimer IFN-γ to its receptor activates the Janus-activated kinase (JAK)-STAT pathway. This JAK-STAT signal harmonizes the transcriptional activation of several genes and mediates various biological responses. 13 High titers of anti-IFN-γ AAbs are able to block the binding of IFN-γ to its receptor which inhibit the early aspects of IFN-γ signal transduction, either STAT-1 phosphorylation or STAT-1 protein expression. These autoantibodies also inhibit the downstream biological consequences of IFN-γ binding which are the up-regulation of tumor necrosis factor (TNF)-α and interleukin (IL)-12 production.14,15 Purified anti-IFN-γ AAbs is able to block the induction of IFN-γ-inducible genes and the upregulation of HLA class II expression on peripheral blood mononuclear cells. 16 Moreover, patients that were anti-IFN-γ AAbs positive could block IFN-γ-mediated antimicrobial immunity in monocytes and macrophages. These include IFN-γ-driven polarization and M1 macrophages activation, the production of cytokines, chemokines and inducible nitric oxide (iNO)/nitric oxide (NO), biosynthesis and reactive oxygen species (ROS) generation, and phagocytosis and degradation efficacy.14,15 These studies clearly demonstrate that anti-IFN-γ AAbs with neutralizing activity can impede the binding of IFN-γ to its receptor, resulting in the absence of downstream signal transduction, directly affecting immune responses against intracellular pathogens.
Characteristics of anti-IFN-γ AAbs
Most anti-IFN-γ AAbs belong to the IgG isotypes, most frequently IgG4 and IgG1 subtypes.1,7,17–21 Several reports showed heterogeneous patterns of isotypes and subtypes among patients. A predominance of IgG3 was also reported in some patients. 18 Our study demonstrated that anti-IFN-γ AAbs mainly exhibited the IgG1 and IgG4 subtypes. However, multiple isotypes and subtypes can be found in some cases. 21 These data suggest that B cells that produce anti-IFN-γ AAbs have undergone antibody class switching.
Several reports found that these autoantibodies recognized an epitope in the C-terminal region of IFN-γ.8,17,21 The data suggest that the major epitope contains the SPAAKTGKRK amino acid sequence, determined by computer modeling. We also found that autoantibodies against IFN-γ in Northern Thai patients recognized the C-terminal linear epitope containing the KRKR motif. 21 This region is critical for IFN-γ receptor (IFNGR) activation and the binding of these autoantibodies impede IFN-γ-mediated activities. 8
Commercially available mouse anti-IFN-γ monoclonal antibodies can bind to distinct epitopes of the IFN-γ molecule with different neutralizing activity. 22 Competitive-binding ELISA using commercial neutralizing mouse anti-IFN-γ monoclonal antibodies showed that anti-IFN-γ AAbs in patients bind to discontinuous epitope of homodimeric IFN-γ. This study also demonstrated the heterogeneity of the auto Abs against IFN-γ in AOID patients and the diverse patterns among individuals. In-depth analysis of binding epitopes in individuals may provide information for designing therapeutic approaches.
Anti-IFN-γ AAbs and pathology
Anti-IFN-γ AAbs has been associated with various opportunistic infections. Disseminated nontuberculous mycobacterial (NTM) infection is the most common infectious disease,1,6 but Talaromyces marneffei, Cryptococcus neoformans, Histoplasma capsulatum, Burkholderia pseudomallei, Salmonella species, and Varicella–zoster virus are also frequently identified as opportunistic pathogens in this patient group.1,2,6,23 Concomitant infections with at least 2 opportunistic pathogens are more accurate parameters to confirm the immunocompromised state in these patients.1,2 Several organ systems are involved, in which lymph nodes, skin, bone and soft tissue appear to be frequently affected.1,2,6 Bone marrow or blood, lung, bladder, liver, and biliary tree are likewise the sites of infection.6,24–26 The clinical manifestations, locations of the infected sites, and causative agents are likely to differ across ethnicities. Rapidly growing mycobacteria (RGM), such as M. abscessus, were the most common NTM species isolated from Thai, Chinese and Filipino patients, whereas M. avium complex (MAC) was predominant among Japanese and non-Asian patients.6,27 Lymph nodes were the most common organ involved in patients with RGM, whereas bone and lung infections were more common in patients with MAC.
Many studies showed that disseminated NTM in patients with anti-IFN-γ autoantibodies can mimic malignancy, cancer, and synovitis-acne-pustulosis-hyperostosis-osteitis (SAPHO) syndrome.26,28,29 Therefore, differential diagnosis should be handled delicately. Most reported cases of patients with anti-IFN-γ AAbs are older adults. Nevertheless, adolescents and young adults with disseminated opportunistic pathogenic infections with anti-IFN-γ AAbs have been reported recently.30,31 The majority of patients reported to have anti-IFN-γ AAbs have been narrowed to Asia-born Asians, particularly those from Southeast Asian populations with human leukocyte antigen (HLA)−DRB1 and DQB1 alleles, especially HLA−DRB1*15:01/16:02 and DQB1*05:01/05:02.32–34 However, opportunistic pathogenic infections have been reported in Caucasian patients with anti-IFN-γ AAbs living in the UK, USA, Germany as well as in patients of African ancestry.16,19,20,35 Although correlations between anti-IFN-γ AAbs, types of opportunistic infections, clinical manifestations, HLA alleles, and ethnicity have been demonstrated, other issues, such as environmental factors and genetic diversity must be elucidated for in-depth understanding of this disease. Interestingly, by fitting a linear mixed model adjusted for age, sex, as well as the presence of infection, antibiotic use and cyclophosphamide or rituximab use, anti-IFN-γ AAbs levels decreased overtime regardless of ethnicity, even in the absence of immunomodulatory therapy. 6 Neither age nor sex was a discriminating factor in the antibody levels over time. Therefore, the factors predicting specific outcomes remain to be revealed.
Detection of anti-IFN-γ AAbs
Considering the significance of anti-IFN-γ AAbs-associated pathology, it is important to evaluate antibody detection even though most patients clinically present with disseminated infections. Currently, the approaches used to evaluate anti-IFN-γ AAbs in suspected cases with certain infections vary greatly from simple assays to long tedious bioassays. We have summarized the different wide-ranging methods from qualitative or quantitative IFN-γ-specific binding and biological activity assays based on previous reports (Figure 1). The most common assay used to detect anti-IFN-γ AAbs is the enzyme-linked immunosorbent assay (ELISA), in indirect,2,12,17,18,21,23,36–41 sandwich,14,16,32,42 or inhibitory2,3,8,17,23,32,37,43,44 assay format. For the ELISA-based method, the results could be reported qualitatively (positive or negative) or quantitatively as optical density (OD), 39 titer, 43 or through calculations of arbitrary units 21 or ELISA units (EU).36,39 The easy-to-use, low-cost Dot ELISA strip, which can be read directly on the strip, was developed as a point-of-care screening tool in remote settings.8,45 A particle-based assay with claims as a fast, easy, and relatively inexpensive technique has also been used to detect multiple anti-cytokine autoantibodies simultaneously in human plasma.6,18,46 QuantiFERON-TB Gold In-tube (QFT-GIT), a commercialized IFN-γ release assay commonly used for detection of latent tuberculosis in many hospitals has also been modified for the screening of neutralizing anti-IFN-γ AAbs.38,47 Luciferase immunoprecipitation system (LIPS) is a high throughput quantitative method useful for detecting multiple anti-cytokine autoantibodies at the same time.1,48 Immunoblot of IFN-γ have also been used to verify the exact nature of the binding activity.8,14,40,44
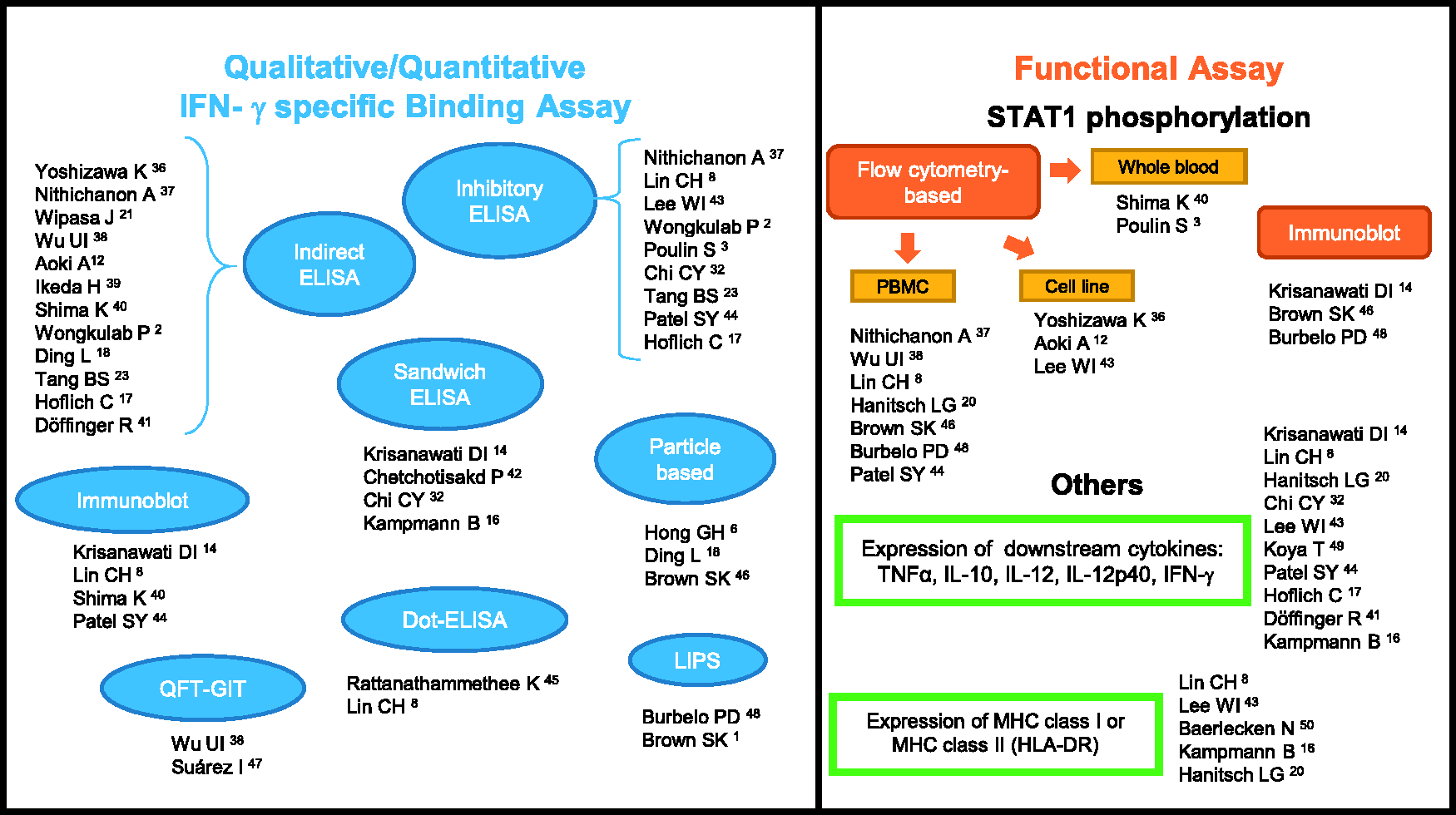
Summarized methods of detection, the qualitative/quantitative IFN-γ specific binding, and functional assays of anti-IFN-γ AAbs. IFN-γ: interferon gamma; AAbs: autoantibodies; ELISA: enzyme-linked immunosorbent assay; QFT-GIT: QuantiFERON-TB gold in-tube; LIPS: luciferase immunoprecipitation system; STAT1: signal transducer and activator of transcription 1; PBMC: peripheral blood mononuclear cell; TNF: tumor necrosis factor; IL: interleukin; MHC: major histocompatibility complex. (A color version of this figure is available in the online journal.)
Assessment of pSTAT1 is an important approach often used to confirm the anti-IFN-γ AAbs inhibitory function, commonly performed by flow cytometry3,8,12,20,36–38,40,43,44,46,48 or immunoblot.14,46,48 Shima et al. demonstrated that all disseminated NTM participants with inhibited STAT1 phosphorylation have high titers of anti-IFN-γ AAbs. 40 However, non-concomitant results in three participants with high-titer anti-IFN-γ AAbs but without opportunistic infections showed no IFN-γ–blocking activity. 1 A report by Lin et al. illustrated that among healthy individuals who possessed autoantibodies against IFN-γ, no neutralizing activity nor effect to IL-12 production was found. 8 These findings suggest that detection of the existence and the level of anti-IFN-γ AAbs may not be enough for evaluation of clinical management. Other biological activity assays include evaluation of IFN-γ downstream effector molecules.8,14,16,17,20,32,41,43,44,49,50
To date, a standard method has not yet been endorsed. Therefore, sensitivity, specificity, positive predictive value (PPV), and negative predictive value (NPV) of assays should be taken into consideration when selecting a screening method. Nithichanon et al. demonstrated that inhibitory ELISA is more specific and has greater PPV and NPV than indirect ELISA with comparable sensitivity. 37 Some methodological limitations are of concern. For example, pSTAT1 is diminished in a day dependent manner when kept at 4°C, therefore it is important to take into account the timing of the assay following phlebotomy when using whole blood for pSTAT1 detection. 40 Several methods may be required to confirm the presence and functional activities of anti-IFN-γ AAbs.
Clinical management of anti-IFN-γ AAbs
Sensitive antimicrobial therapy is the most important strategy for eliminating infections, including in patients carrying anti-IFN-γ AAbs. The most common drugs used for the treatment of NTM are a regimen combining rifampin, ethambutol, clarithromycin, linezolid, and amikacin.4,19,26,39,51–53 Anti-fungal medicines that include amphotericin B, itraconazole, and fluconazole have been used for the treatment of Talaromyces marneffei and Cryptococcus spp.54,55 Nevertheless, the disease can be refractory and fatal, despite appropriate and aggressive antimicrobial treatment.3,46,56
In addition to antimicrobial therapy for clinical disease management, there is no standard treatment guideline for anti-IFN-γ AAbs. Treatment with various immunomodulatory agents has been evaluated in order to modulate humoral immunity. Rituximab, a chimeric monoclonal antibody specific to CD20 expressed by B cells, is the most studied immunomodulatory agent in patients carrying anti-IFN-γ AAbs. Treatment with rituximab results in depletion of circulating B cells, reduction of anti-IFN-γ AAbs titers, restoration of IFN-γ signaling, and improvement of clinical conditions and inflammatory markers.46,57,58 A combination of methylprednisolone and rituximab treatment produced favorable outcomes.58,59 Nevertheless, clinical relapse has been observed in some patients and the time to clinical improvement, reduction of anti-IFN-γ AAbs titers, and improved IFN-γ signaling differed among patients. 46 Therefore, optimization of the immunomodulatory agents is needed to constitute appropriate duration and dosage of the therapies. One patient did not respond to multiple doses of anti-CD20 monoclonal antibody, but an anti-CD38 monoclonal antibody targeting plasma cells (daratumumab) successfully suppressed the antibody levels with clinical improvement. 60 Although immunomodulatory agents could successfully offer effective treatment for autoantibodies, these regimens may not be required for patients who respond to antimicrobial therapy alone. 46 Therapies directed at autoantibodies may be important adjunct treatment for patients with high-titer anti-IFN-γ AAbs who continue to have persistent and progressive infection despite long-term antimicrobial therapy.6,46
In settings where rituximab is less accessible, cyclophosphamide, an alkylating agent capable of inhibiting protein synthesis through DNA and RNA crosslinking, may be an alternative option. Treatment of patients carrying anti-IFN-γ AAbs with cyclophosphamide results in the decrease of the antibody titer with clinical improvement, faster complete remission, a longer duration of remission, and a lower incidence of relapsed infection compared with patients receiving rituximab.42,61 This may be due to broader immune suppression on various immune cells.
In addition to immunomodulatory agents, it has been demonstrated that a variant of IFN-γ, in which the anti-IFN-γ AAbs binding epitope was replaced with a corresponding sequence from mouse, is capable of restoring IFN-γ signaling in vitro despite the presence of patient sera. 8 Restoration of IFN-γ responses by using a variant of IFN-γ or an epitope-deprived peptide offers a potential treatment regardless of depletion of the anti-IFN-γ AAbs. Nevertheless, more investigations in vivo are needed to apply this approach for clinical purposes.
Conclusions
Cumulative reports on anti-IFN-γ AAbs reveal increasing impact of anti-IFN-γ AAbs-associated opportunistic infections. This not only affects health, but also impedes the welfare and economic status of the patients and caregivers, as well as nations. This review provides insight into the context of the etiology, pathology, function, diagnostic methodologies, characteristics, and clinical management of anti-IFN-γ AAbs. This information may promote better understanding of adult onset immunodeficiency associated with anti-IFN-γ AAbs.
Footnotes
Authors’ contributions
All authors wrote and approved the manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
This report was partly funded by Chiang Mai University, Chiang Mai, Thailand and the Thailand Research Fund (RSA6280065 to JW).