
Abstract
Circulating cell-free DNA (cfDNA) has attracted attention as a non-invasive biomarker for diagnosing and monitoring various cancers. Given that human papillomavirus (HPV) DNA integration and overexpression of E6/E7 oncogenes are pivotal events for carcinogenesis, we sought to determine if HPV E7 cfDNA could serve as a specific biomarker for cervical cancer detection. We applied droplet digital PCR (ddPCR) to quantify HPV16/18 E7 cfDNA from the serum of patients with cervical cancer, cervical intraepithelial neoplasia, and controls. HPV16/18 E7 cfDNA was highly specific for cervical cancer, displaying 30.77% sensitivity, 100% specificity, and an area under the curve of 0.65. Furthermore, we developed a sensitive isothermal detection of HPV16/18 E7 and the PIK3CA WT reference gene based on recombinase polymerase amplification combined with a lateral flow strip (RPA-LF). The assay took less than 30 min and the detection limit was 5–10 copies. RPA-LF exhibited 100% sensitivity and 88.24% specificity towards HPV16/18 E7 cfDNA in clinical samples. The agreement between RPA-LF and ddPCR was 83.33% (κ = 0.67) for HPV16 E7 and 100% (κ = 1.0) for HPV18 E7, indicating a good correlation between both tests. Therefore, we conclude that HPV E7 cfDNA represents a potential tumor marker with excellent specificity and moderate sensitivity for minimally invasive cervical cancer monitoring. Moreover, the RPA-LF assay provides an affordable, rapid, and ultrasensitive tool for detecting HPV cfDNA in resource-limited settings.
Keywords
Impact statement
Our data show that HPV E7 cfDNA is a potential surrogate blood-based biomarker with high specificity and modest sensitivity for late-stage cervical cancer, and may serve for the monitoring of patients with an already confirmed diagnosis. To our knowledge, this is the first report describing the utilization of the RPA-LF assay for rapid and PCR-free detection of HPV16/18 E7 cfDNA and the PIK3CA WT reference gene. The assay is comparably sensitive and correlates well with ddPCR, thus offering an alternative to currently available molecular detection methods for cervical cancer monitoring, especially in remote areas with limited resources.
Introduction
Cervical cancer is the fourth most common female malignancy worldwide, contributing to 569,847 cases and 311,365 deaths globally in 2018. 1 In spite of advancements in vaccination and screening, the disease ranks second in both incidence and mortality in countries with low/medium human development index. 1 Human papillomavirus (HPV) is a non-enveloped, double-stranded DNA virus associated with over 5% of cancers, including cancer of the head and neck, oropharynx, vagina, penis, anus, and cervix. 2 Among the 15 high-risk oncogenic HPV genotypes, HPV16 and HPV18 account for 70% of cervical cancers. 2 Up to 40% of precancerous lesions, known as cervical intraepithelial neoplasia (CIN), can regress to normal, but lesions harboring HPV genome integration may lead to overexpression of E6 and E7 oncoproteins, preventing lesion regression. 3 Because HPV E6/E7 can target p53 and Rb, and induce all cancer hallmarks, they could predict future malignant transformation. 4
Traditional cervical cancer screening is based on the highly subjective Pap test, characterized by 55.4% sensitivity and 96.8% specificity for CIN2 or CIN3, and HPV DNA detection from cervical specimens, which achieves 94.6% sensitivity and 94.1% specificity. 5 These methods, however, require vaginal examination, limiting participation in Thailand and other developing countries. 6 Minimally invasive liquid biopsy tests could potentially replace conventional techniques, improve women’s admittance to cervical screening programs, and lower the burden of cervical cancer.
Cell-free DNA (cfDNA) contains degraded DNA fragments of 120–220 bp or multiples thereof, and is released into circulation by various modes of cell death and active secretion processes.7,8 CfDNA has been explored as a liquid biopsy biomarker for detecting and monitoring cancer, as well as for identifying resistance and drug targets. 9 Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit α (PIK3CA) mutations E542K or E545K have been found in plasma cfDNA in cervical cancer patients, where they correlated with tumor size and overall survival. 10 Methylated single-minded homolog 1 in plasma cfDNA has also been reported to be highly specific and exhibit moderate sensitivity for diagnosing cervical cancer. 11
Recently, HPV cfDNA has become of major interest as a potential biomarker for the diagnosis and prognosis of cervical cancer.12–14 The unique viral sequence, the abundance of copies upon subsequent amplification, and integrated HPV genomes in cancer cells make HPV cfDNA a favorable marker for detecting cervical cancer. 15 At present, detection of HPV cfDNA remains challenging due to the low sensitivity of traditional quantitative PCR (qPCR), resulting in positive rates in invasive cervical cancer of 6.9% to 50%.12,15–21 Droplet digital PCR (ddPCR) offers better sensitivity and quantification of poorly abundant nucleic acids including HPV cfDNA.14,22 However, the procedure is time-consuming and requires expensive equipment and highly trained staff. To overcome these issues, particularly in under-equipped laboratories or remote areas, alternative simple and rapid HPV cfDNA-based methods with comparable sensitivity to ddPCR are required.
Recombinase polymerase amplification (RPA) has recently emerged as a revolutionary method for nucleic acid detection. 23 RPA utilizes recombinase and single-strand DNA binding protein to replace the heat denaturation step in PCR and replication is performed by DNA polymerase. 24 The reaction is fast and occurs at low constant temperature (25–42°C), eliminating the need for a thermocycler. 24 RPA has been used for rapid detection of various pathogens, such as Escherichia coli O157:H7, 25 Orientia tsutsugamushi,26,27 Rickettsia typhi, 27 Mycobacterium avium, 28 and HIV,29,30 as well as for detection of cancer biomarkers, including TMPRSS2-ERG mRNA in urine for prostate cancer diagnosis, 31 EGFR mutations in lung cancer cells, 32 and HPV16/18 DNA for cervical cancer screening. 33 RPA can be combined with a lateral flow strip (RPA-LF) for rapid visualization by the naked eye while retaining sensitivity and being highly concordant with qPCR.28,34 We hypothesized that HPV E7 cfDNA detection based on RPA-LF could provide rapid, simple, and point-of-care diagnostics of cervical cancer especially in developing countries.
In this study, we sought to (I) test the diagnostic performance of HPV16 and HPV18 E7 cfDNA for cervical cancer detection from the serum of healthy controls, patients with CIN, and cervical cancer patients using a highly sensitive ddPCR; (II) develop the isothermal RPA-LF assay for detection of HPV16/18 E7 and internal control; and (III) compare RPA-LF versus ddPCR for the detection of HPV16/18 E7 cfDNA in clinical specimens. We report that HPV E7 cfDNA is highly specific and could be utilized as a liquid biopsy biomarker for monitoring cervical cancer patients with a confirmed diagnosis. Further, the RPA-LF assay offers a fast, highly sensitive alternative to ddPCR for the detection of HPV E7 cfDNA in resource-limited areas. This minimally invasive liquid biopsy test may potentially augment conventional techniques, improve women’s health, and lower the burden of cervical cancer.
Materials and methods
Serum sample collection
Serum sample collection was approved by the Vajira Institutional Review Board, Faculty of Medicine, Vajira Hospital, Navamindhadhiraj University (COA 091/2019), in compliance with the Helsinki Declaration. A written consent form was obtained from volunteers prior to blood collection. Serum specimens from 39 cervical cancer patients (14 Stage I/II and 25 stage III/IV), 5 patients with CIN (1 CIN1 and 4 CIN3), and 29 controls were collected at the Department of Obstetrics and Gynecology, Vajira Hospital (Table 1). Controls were obtained from healthy volunteers without any evidence of cancer and with a negative Pap test within the past year. Patients with CIN or cervical cancer were diagnosed with colposcopic-directed cervical biopsies. Peripheral blood was collected, allowed to clot for 30 min, centrifuged at 3000g for 15 min at 4°C, and serum was stored at −80°C until use.
Clinical characteristics and data from cfDNA analysis of cervical cancer patients, CIN, and controls used for ddPCR and RPA-LF assays.
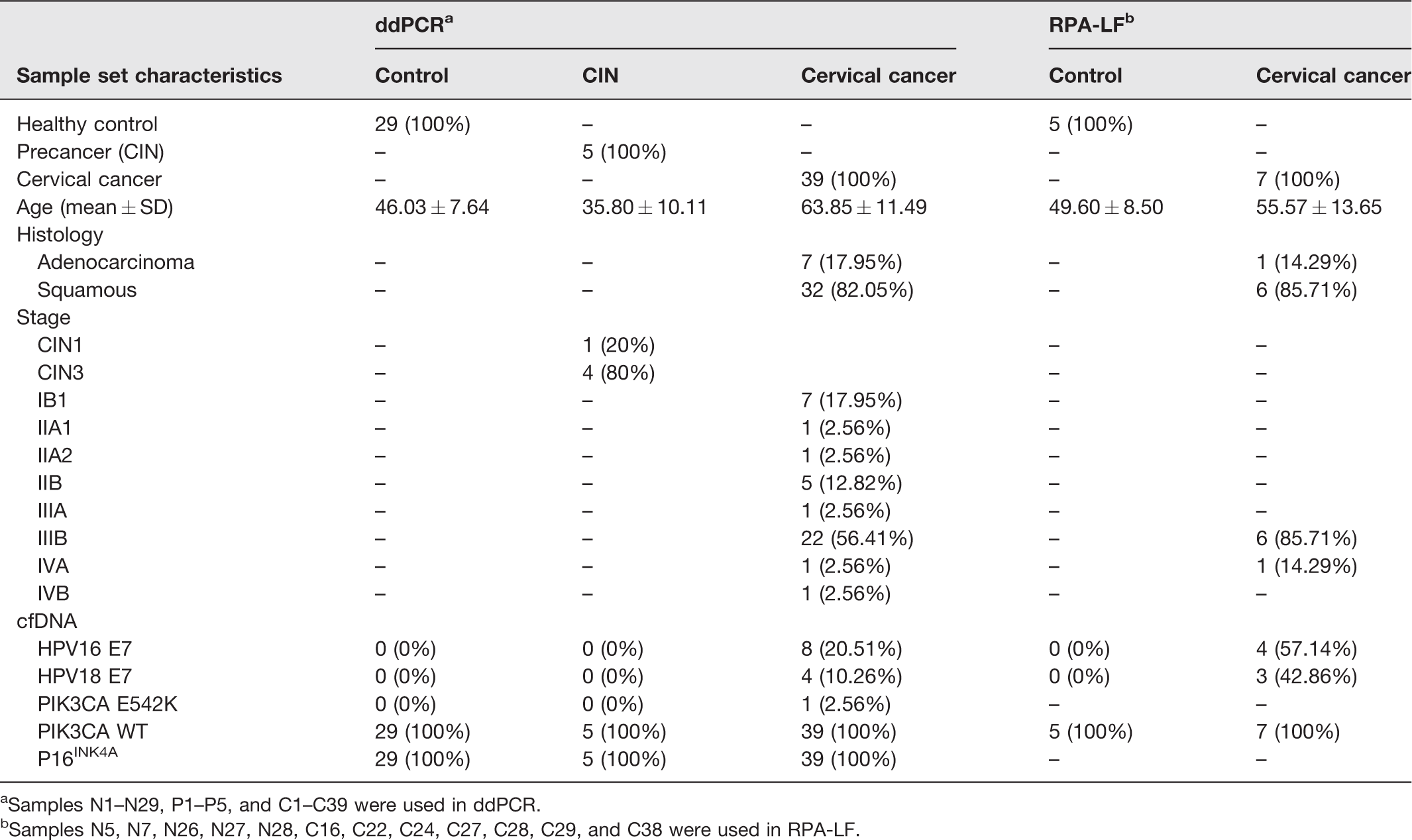
aSamples N1–N29, P1–P5, and C1–C39 were used in ddPCR.
bSamples N5, N7, N26, N27, N28, C16, C22, C24, C27, C28, C29, and C38 were used in RPA-LF.
Cell-free DNA extraction and analysis
Total circulating cfDNA was extracted from approximately 800 µL of serum using the Maxwell RSC® ccfDNA plasma kit (AS1480; Promega, Madison, WI, USA) as per the manufacturer’s instructions. The extracted cfDNA was eluted with 60 µL elution buffer and kept at −80°C. Thirty-five samples of cfDNA, including 17 cervical cancers and 18 controls, were randomly selected to determine fragment size and total cfDNA concentration using the Fragment Analyzer™ Automated CE System and High Sensitivity Small Fragment kit (DNF-477-0500; Agilent, Santa Clara, CA, USA). Data were analyzed by ProSize Data Analysis Software (Agilent).
HPV cfDNA detection and quantification by ddPCR
Primers and probes specific for HPV16 E7 and HPV18 E7 genes, 35 PIK3CA wild-type (WT), PIK3CA E542K, and p16INK4A were indicated in Table S1. The pUC57 plasmid templates containing nucleotide sequences of HPV16 E6-E7, HPV18 E6-E7, and PIK3CA were synthesized by GenScript (Piscataway, NJ, USA) and used for assay validation or positive controls (Table S2). A human mixed genomic DNA (G3041; Promega) served as a negative control for HPV16/18 E7 and PIK3CA E542K. The ddPCR reactions contained 1× ddPCR Supermix Probes (Bio-Rad, Hercules, CA, USA), 900 nmol/L primers, 250 nmol/L FAM- and/or HEX-labeled probes, and 2 µL DNA template. The 96-well plate containing ddPCR reactions was transferred to an Automated Droplet Generator (Bio-Rad). The plate was sealed with a pierceable foil using a PX1™ PCR Plate Sealer (Bio-Rad) at 180°C for 5 s and transferred to a T100™ Thermal Cycler (Bio-Rad). PCR amplification was as follows: 95°C for 10 min, 40 cycles of 94°C for 30 s and 55°C for 1 min, followed by 98°C for 10 min, with a fixed ramp rate of 2°C/s. PCR results were read by a QX200™ Droplet Reader (Bio-Rad) and analyzed by QuantaSoft™ version 1.7.4.0917 (Bio-Rad).
To quantify target DNA in clinical cfDNA specimens, duplex HPV16/18 E7, duplex PIK3CA WT/E542K, and singleplex p16INK4A ddPCR assays were performed in duplicate wells and no template control (NTC) was included in every run. PIK3CA WT served as a reference gene to indicate the quantity of cfDNA.
Recombinase polymerase amplification with lateral flow strip
The primers and probes for the RPA-LF assays were designed based on the manual of the TwistAmp® DNA Amplification Kits (TwistDx, Maidenhead, UK) and synthesized by Integrated DNA Technologies (Coralville, IA, USA). DNA sequences are summarized in Table S4. The RPA reaction included 10 µmol/L each of forward and reverse primers, 10 µmol/L RPA probe, 29.5 µL rehydration buffer, 1 µL DNA template, and nuclease-free water. The plasmid templates harboring sequences of HPV16 E6-E7, HPV18 E6-E7, or PIK3CA served as positive controls; nuclease-free water served as NTC. The reaction was initiated by adding 2.5 µL of 280 mmol/L magnesium acetate, followed by incubation at 37°C for 20 min. Subsequently, RPA products were diluted 50-fold and applied on the sample pad. The lateral flow strip (HybriDetect; Milenia Biotec GmbH, Giessen, Germany) was dipped into 200 µL assay buffer for 7 min prior to visualization. PIK3CA WT was used as an internal control.
Statistical analysis
Boxplots and violin plots were created by BoxPlotR36 and statistical analysis was performed using GraphPad InStat version 3 (GraphPad Software, La Jolla, CA, USA). The two-tailed Mann–Whitney U test was used to compare two groups, whereas the Kruskal–Wallis test with Dunn's post-test was used when comparing three or more groups. The sensitivity is defined as the ability of a biomarker to correctly identify a person with a disease (number of true positives/[number of true positives + number of false negatives) while the specificity refers to the ability of a biomarker to correctly identify people who do not have a disease (number of true negatives/[number of true negatives + number of false positives]). Receiver operating characteristic (ROC) curves and statistical analysis were generated by easy ROC web-tool. Diagnostic test statistics were calculated by MedCalc® (MedCalc Software Ltd, Ostend, Belgium; www.medcalc.org/calc/diagnostic_test.php). Tests were compared by Epitools Epidemiological Calculators (http://epitools.ausvet.com.au). A P value < 0.05 was considered statistically significant.
Results
Analysis of cfDNA from serum samples
To determine whether HPV16 and HPV18 E7 cfDNA could be used as blood-based biomarkers for cervical cancer detection, we first measured HPV16 and HPV18 E7 cfDNA levels from 73 serum samples, including 39 cervical cancer patients, 5 patients with CIN, and 29 healthy volunteers (Table 1). Fragment analysis revealed typical mono-, di-, and tri-nucleosome fragments in cfDNA samples (Figure 1(a)). We randomly selected and analyzed fragment size and total cfDNA concentration from 35 samples, including 17 cervical cancer patients and 18 controls. Fragments of cfDNA from control samples (median 175.25 bp, 95% CI: 175.02–180.76 bp) were significantly longer than those from cervical cancer patients (median 171 bp, 95% CI: 170.28–176.77 bp; P = 0.0089, Mann–Whitney U test; Figure 1(b)). As shown in Table S3 and Figure 1(c), total cfDNA yields from serum were very low and varied substantially across samples, resulting in no significant difference between cervical cancer patients and normal groups (P = 0.573, Mann–Whitney U test). Hence, serum cfDNA concentration failed to discriminate cervical cancer patients from controls in our sample set.
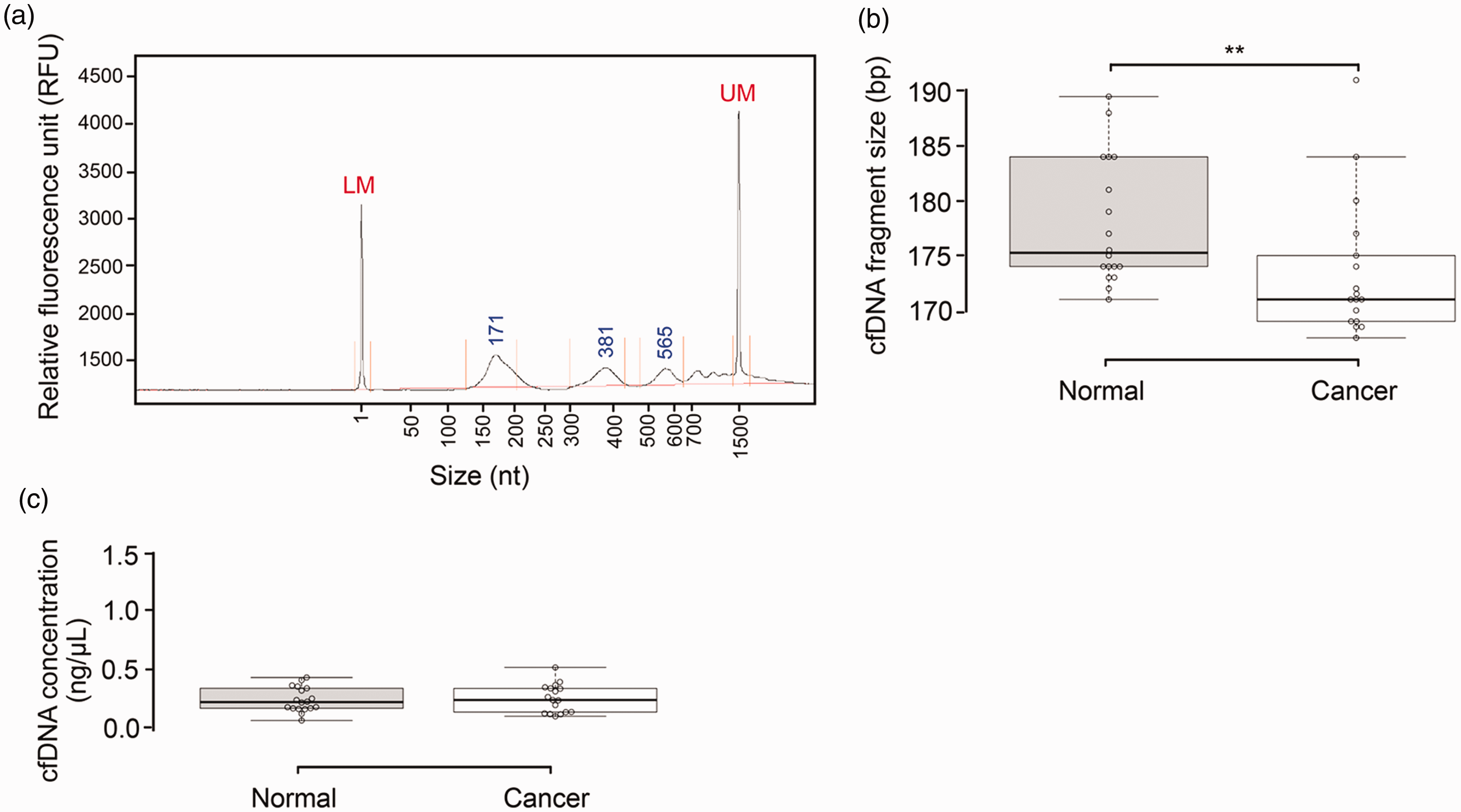
Size distribution and total concentration of cfDNA isolated from the serum of controls and cervical cancer patients. (a) Representative post-separation electropherogram of serum cfDNA from cervical cancer patient C38. LM and UM represent 1-nt and 1500-nt markers, respectively. (b and c) Boxplots showing the size distribution of cfDNA fragments (b) and cfDNA levels (c) in samples from healthy controls (N = 18) and cervical cancer patients (N = 17). Centerlines indicate the medians and box limits denote the 25th and 75th percentiles. Data points are plotted as open circles. Each sample set was compared using the non-parametric Mann–Whitney test (**P < 0.01, two-tailed).
Validation of the ddPCR assay for target DNA detection
We utilized ddPCR for absolute quantification of HPV16 and HPV18 E7 as target biomarkers and PIK3CA WT as a reference gene. To confirm the suitability of the assays, serial dilutions of synthetic plasmids pUC57_HPV16 E6-E7, pUC57_HPV18 E6-E7, and pUC57_PIK3CA were used as templates for singleplex ddPCR detection (Table S2). As shown in Figure S1, we observed good linearity between spectrophotometrically determined log copy number of plasmid DNA and ddPCR (R2 = 0.99) over a dynamic range of 2 to 20,000 copies/reaction. The limit of detection (LOD) with 95% CI of PIK3CA WT was estimated to be 2.45 ± 1.16 copies/reaction (Figure S2(C)).
To develop the duplex ddPCR assay for simultaneous detection of HPV16/18 E7 DNA, we next determined the specificity and sensitivity of the assay. In the presence of HPV16 E6-E7 or HPV18 E6-E7 plasmid DNA, only positive droplets corresponding to the DNA input were detected, indicating no cross-reactivity (Figure S3(A) vs. (B)). In addition, when testing a mixture of HPV16 and HPV18 E6-E7 plasmid DNAs, two droplet populations could be simultaneously observed (Figure S3(C)); whereas only negative droplets were obtained in the absence of plasmid template (NTC; Figure S3(D)). This finding confirmed the specificity of the duplex HPV16/18 E7 ddPCR assay. To evaluate the sensitivity of the duplex assay, the LOD with 95% CI was determined. We could reliably detect as few as 2.13 ± 0.5 copies/reaction (80 copies/mL serum) of HPV16 E7 DNA and 2.30 ± 0.97 copies/reaction (86 copies/mL serum) of HPV18 E7 DNA, with a detection rate of 100% (Figure S2(A) vs. (B)). These results showed that the duplex ddPCR assay was specific, sensitive, and applicable for multiplex detection of HPV16 and HPV18 E7 DNA.
Suitability of PIK3CA WT DNA as a reference gene for cfDNA analysis
To ensure sufficient quantity and quality of the sample, we tested PIK3CA WT as a reference gene for cfDNA analysis. As shown in Tables 1 and S3, as well as Figure 2, we detected PIK3CA WT DNA in 100% of cfDNA samples and human genomic DNA but not in the NTC. In addition, PIK3CA WT copy number correlated linearly with total cfDNA concentration (r = 0.42; Figure S4(A)). Moreover, the level of PIK3CA WT DNA was comparable across normal, CIN, and all stages of cervical cancer (Figure 3(a)). These data demonstrated that PIK3CA WT was a suitable reference gene for cfDNA analysis.
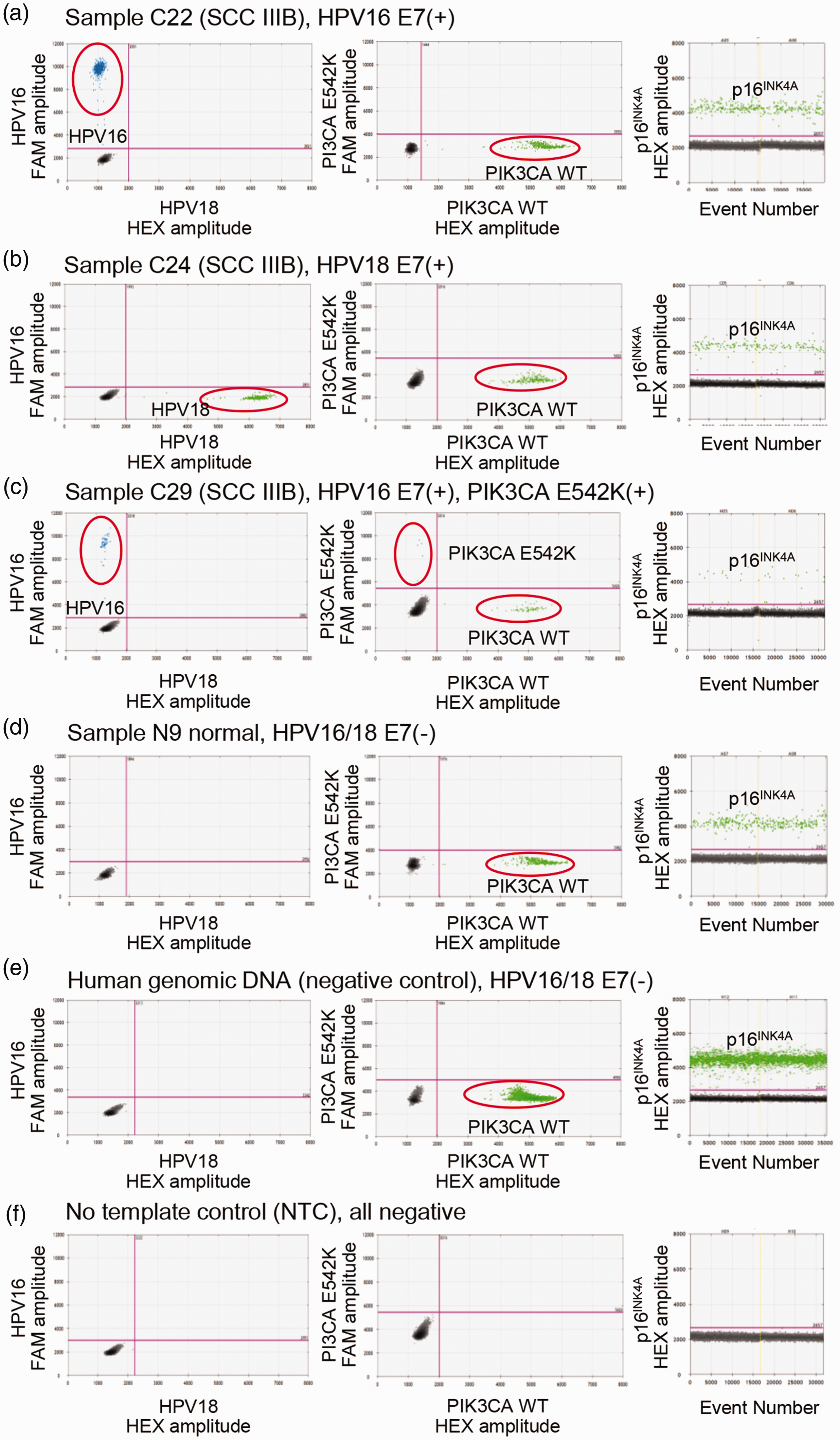
Detection of circulating HPV16 E7, HPV18 E7, PIK3CA WT, PIK3CA E542K, and p16INK4A from serum samples using ddPCR. (a to f) 2D and 1D plots of stage IIIB cervical carcinoma C22 (a), C24 (b), and C29 (c) patients, healthy control N9 patient (d), human genomic DNA (negative control) (e), and no template control (f). Blue dots (FAM) indicate droplets positive for HPV16 and PIK3CA E542K; green dots (HEX) indicate droplets positive for HPV18 and PIK3CA WT by duplex detection. In singleplex detection of p16INK4A, green dots (HEX) show droplets positive for of p16INK4A; black dots indicate negative detection. (A color version of this figure is available in the online journal.)
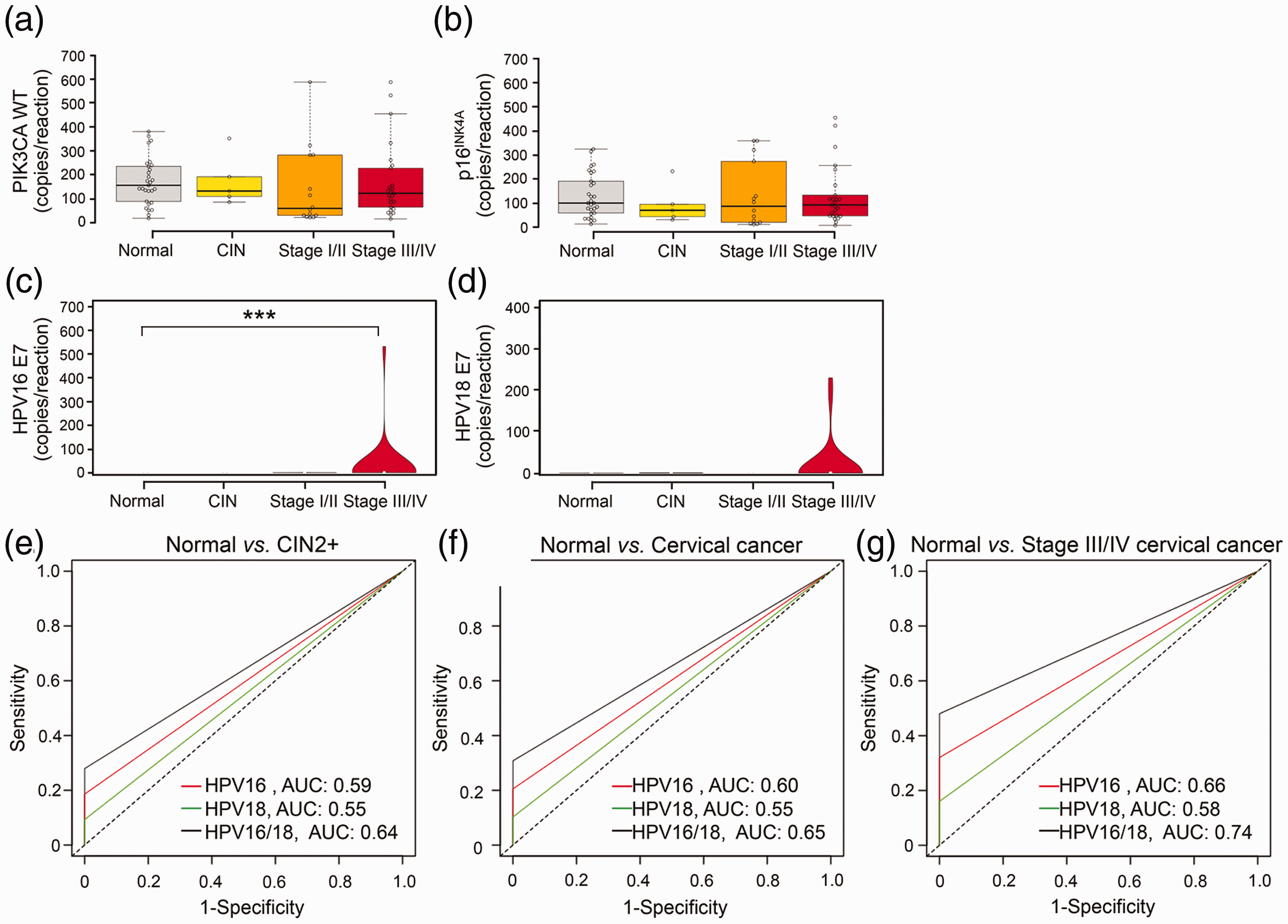
Levels of circulating PIK3CA WT, p16INK4A, HPV16 E7, and HPV18 E7 in clinical samples and analysis of the diagnostic performance of HPV E7 cfDNA by ddPCR. (a and b) Boxplots showing the cfDNA copy number of PIK3CA WT (a) and p16INK4A (b) in normal (N = 29), CIN (N = 5), cervical cancer stages I/II (N = 14), and stages III/IV (N = 25) samples. Centerlines indicate the medians, box limits denote the 25th and 75th percentiles, and whiskers extend 1.5 times the interquartile range from the percentiles. Violin plots showing different levels of HPV16 E7 (c) and HPV18 E7 (d) among the aforementioned four groups. White circles indicate the medians, box limits denote the 25th and 75th percentiles, whiskers extend 1.5 times the interquartile range from the percentiles, and polygons represent density estimates of data and extend to extreme values. Groups were compared using the Kruskal–Wallis test with Dunn’s post-test (***P < 0.001 compared with the normal group). (e to g) ROC curves for HPV16 E7, HPV18 E7, and HPV16/18 E7 cfDNA in normal (N = 29) vs. CIN2+ (N = 43) (e), normal (N = 29) vs. cervical cancer (N = 39) (f), and normal (N = 29) vs. stage III/IV cervical cancer (N = 25) (g). AUC: the area under the curve. (A color version of this figure is available in the online journal.)
We next selected p16INK4A as another candidate reference gene and tested whether it performed similarly to PIK3CA WT. The ddPCR assay revealed that, except for the NTC, p16INK4A-positive droplets were detected in all cfDNA samples including human genomic DNA (Tables 1 and S3, and Figure 2). Moreover, p16INK4A DNA copy number correlated with total cfDNA concentration (r = 0.32; Figure S4(B)) and particularly PIK3CA WT (r = 0.92; Figure S4(C)). Similar to PIK3CA WT, no difference in the p16INK4A cfDNA level was detected across sample groups (Figure 3(b)). Therefore, we concluded that both PIK3CA WT and p16INK4A were suitable reference genes for analyzing serum cfDNA.
Diagnostic performance of HPV16/18 E7 cfDNA
To evaluate the diagnostic potential of HPV16/18 E7 cfDNA, we utilized duplex ddPCR assays for the detection of HPV16 and HPV18 E7 in clinical samples. Figure 2 shows representative ddPCR 2 D plots of stage IIIB cervical cancer patients C22 with HPV16 E7(+) (Figure 2(a)) and C24 with HPV18 E7 (+) (Figure 2(b)), healthy volunteer N29 with HPV16/18 E7(−) (Figure 2(d)), human genomic DNA with HPV16/18 E7(−) (Figure 2(e)), and NTC (Figure 2(f)). Using the LOD of the assay as a cut-off, we found that eight cervical cancer samples were positive for HPV16 E7 (20.51%) and four for HPV18 E7 (10.26%; Tables 1 and S3). In contrast, none of the controls and patients with precancerous lesions were positive for HPV E7 (Tables 1 and S3), indicating specificity for cervical cancers. Quantitative evaluation of positive cervical cancer samples showed an average of 3234 ± 6833 HPV16 E7 and 4442 ± 3861 HPV18 E7 copies/mL of serum. The high SD values reflected the large variation in HPV cfDNA concentration among patients. The level of HPV16 E7 but not HPV18 E7 DNA was significantly higher in stage III/IV cervical cancer compared with the control (Kruskal–Wallis with Dunn’s post-test, P < 0.001; Figure 3(c) vs. (d)). Based on these findings, HPV16 and HPV18 E7 cfDNAs could potentially serve as circulating biomarkers for cervical cancer.
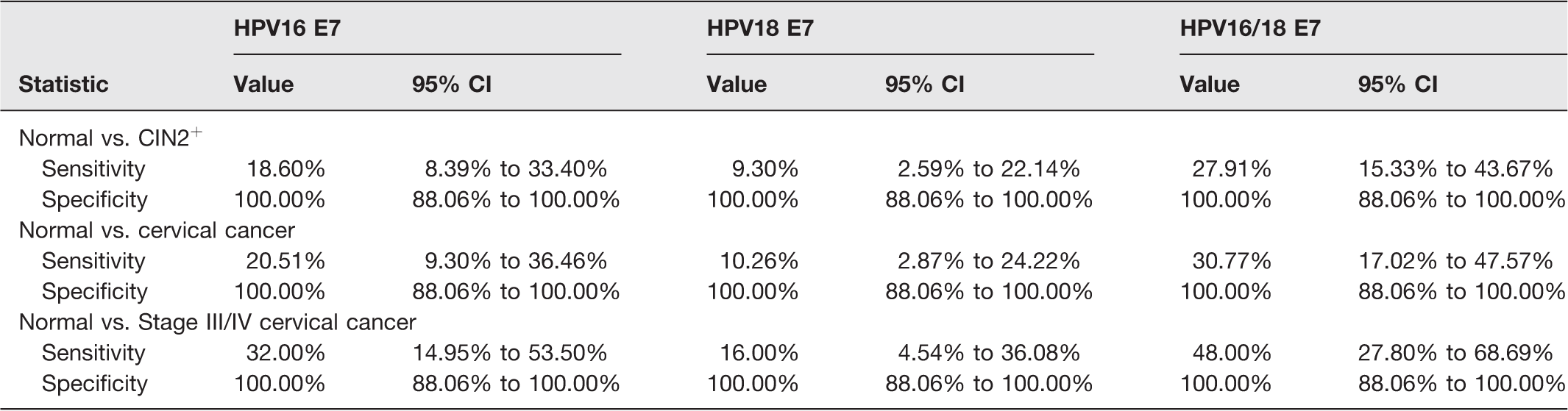
Next, we determined the diagnostic capacity of HPV16 and/or HPV18 E7 cfDNA to discriminate between the disease and control. The sensitivity of HPV16 E7, HPV18 E7, and HPV16/18 E7 was 18.60%, 9.30%, and 27.91%, respectively, in normal (N = 29) vs. CIN2+ (N = 43); 20.51%, 10.26%, and 30.77%, respectively, in normal (N = 29) vs. cervical cancer (N = 39); and 32%, 16%, and 48%, respectively, in normal (N = 29) vs. stage III/IV cervical cancer (N = 25; Table 2). Interestingly, we observed 100% specificity of HPV E7 cfDNA in every group comparison (Table 2).
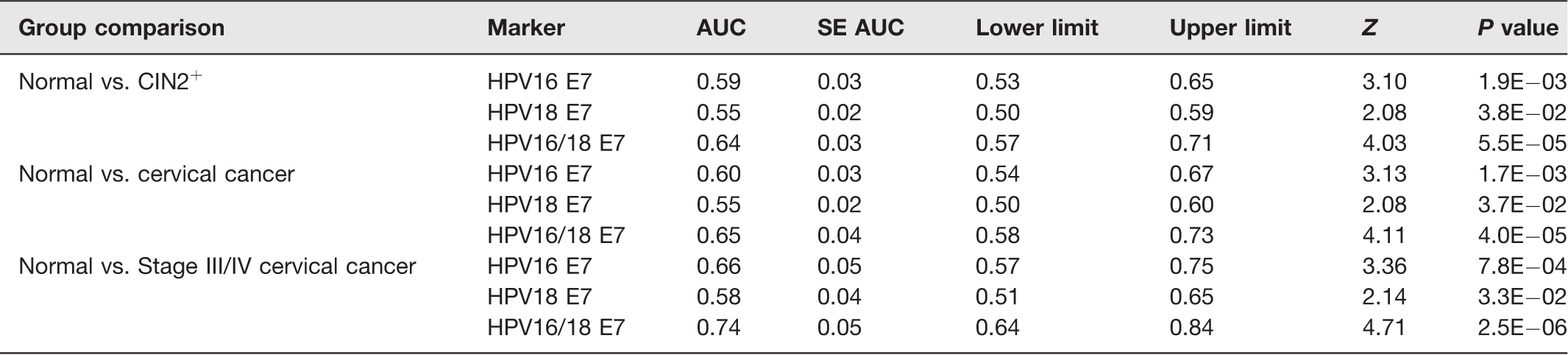
The ROC curve revealed that the area under the curve (AUC) value of HPV16 E7, HPV18 E7, and HPV16/18 E7 was 0.59, 0.55, and 0.64, respectively, in normal vs. CIN2+; 0.60, 0.55, and 0.65, respectively, in normal vs. cervical cancer; and 0.66, 0.58, and 0.74, respectively, in normal vs. stage III/IV cervical cancer (Table 3 and Figure 3(e) to (g)). These results demonstrated that a combination of HPV16 and HPV18 E7 cfDNA represented a highly specific and effective biomarker for separating late-stage cervical cancer patients from controls.
Diagnostic statistics of HPV E7 cfDNA biomarkers in each group comparison.
ROC statistics of HPV E7 cfDNA biomarkers in each group comparison.
Given the link between mutated PIK3CA cfDNA and tumor size, disease-free survival, and overall survival in cervical cancer patients, 10 we tested whether PIK3CA E542K cfDNA could serve as another cervical cancer biomarker and thus improve the diagnostic performance of HPV E7 cfDNA. Using a ddPCR mutation assay, we only detected PIK3CA E542K cfDNA in HPV16 E7(+) stage IIIB cervical cancer patient C29 (2.56%; Tables 1 and S3, and Figure 2(c)). Due to its low positive rate, the PIK3CA E542K cfDNA did not appear suitable as a cervical cancer biomarker in our cohort.
Development of the RPA-LF assay for detection of HPV16/18 E7
The second objective of this study focused on the establishment of a rapid, ultrasensitive, on-site testing method for HPV16/18 E7 cfDNA detection. For rapid visualization, we combined isothermal DNA amplification by RPA with the lateral flow. First, we screened for optimal RPA-LF primer combinations using 5000 copies of plasmid DNA templates (PIK3CA WT, HPV16 E6-E7, HPV18 E6-E7) or nuclease-free water. As shown in Figure S5, F2/R1, F1/R1, and F2/R2 primer combinations produced the highest band intensity (test line) for the detection of PIK3CA WT, HPV16 E7, and HPV18 E7, respectively, without giving any signal in the NTC. Therefore, we selected the aforementioned primer sets for subsequent evaluation. To assess the sensitivity of the RPA-LF assay, we applied serial dilutions of plasmid DNA (5 to 100,000 copies/reaction) as a template. As shown in Figure 4, the LOD of PIK3CA WT, HPV16 E7, and HPV18 E7 was 10, 10, and 5 copies/reaction, respectively. Given that these LOD values were of the same order of magnitude as with ddPCR, the RPA-LF assay was judged highly sensitive.
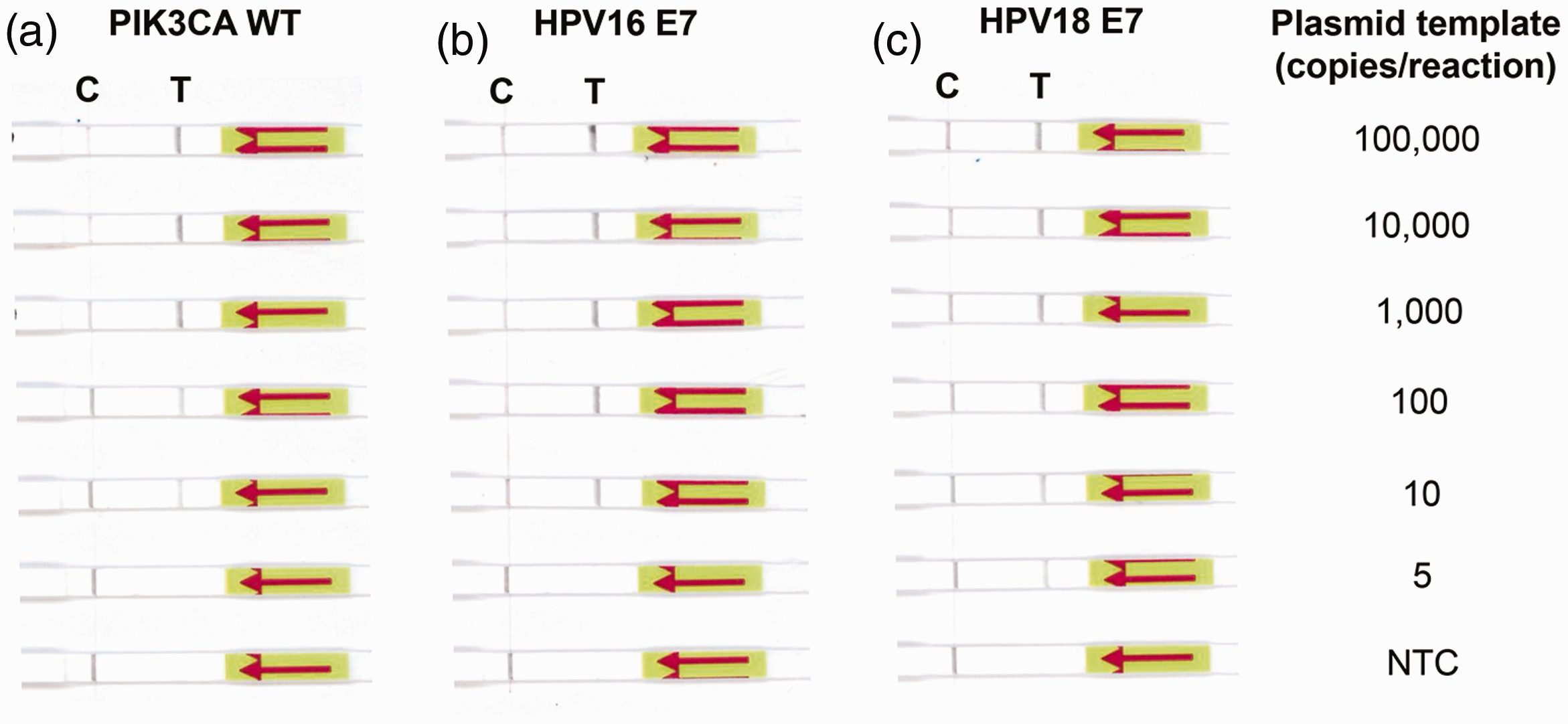
Determination of the RPA-LF assay sensitivity. The LOD of RPA-LF was evaluated for PIK3CA WT (a), HPV16 E7 (b), and HPV18 E7 (c). Serial dilutions of 5–100,000 copies of plasmid DNA were used as the template. Primer sets F2/R1, F1/R1, and F2/R2 were used to detect PIK3CA WT, HPV16 E7, and HPV18 E7, respectively. Nuclease-free water was used as no template control (NTC). C: control line; T: test line. (A color version of this figure is available in the online journal.)
Comparing the diagnostic performance of RPA-LF with ddPCR
Next, we attempted to validate the performance of the RPA-LF assay using clinical samples from seven cervical cancer patients and five controls (Table 1) and compared the result to ddPCR. All cfDNA samples were positive for the PIK3CA reference gene and negative for the NTC control (Figure 5(a)). Samples C16, C22, C27, and C29, previously shown to be positive for HPV16 E7 DNA by ddPCR, were positive also by RPA-LF (Figure 5(b)). However, false-positives were detected in samples N7 and C24 (Figure 5(b)). We correctly identified all HPV18 E7(+) cervical cancer samples and controls without any false-positives (Figure 5(c)). Moreover, as exemplified in samples C16, C22, C27, C28, C29, and C38, we did not observe any cross-reactivity between HPV16 and HPV18 E7 (Figure 5(b) vs. (c)).
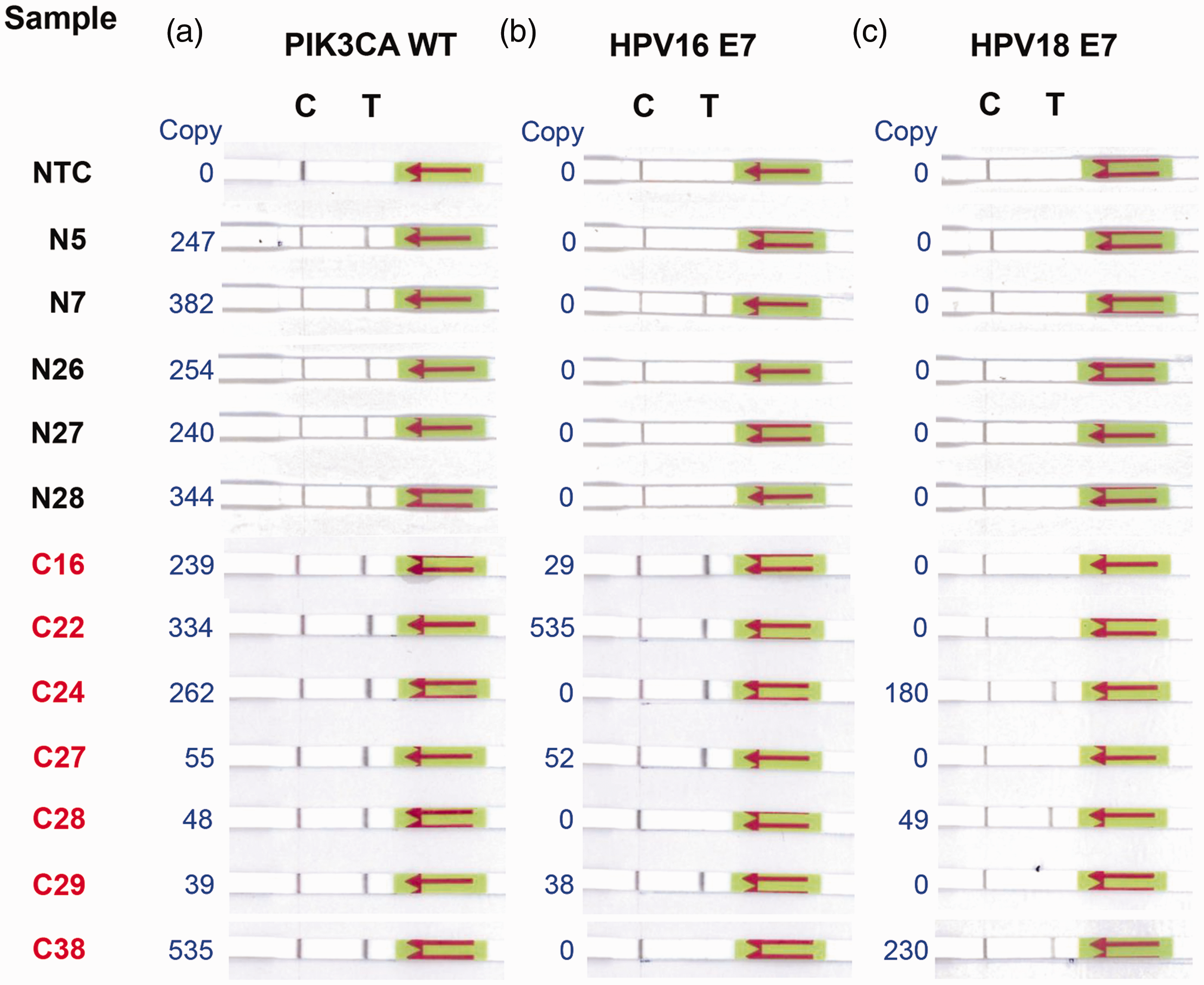
Detection of circulating HPV16 E7, HPV18 E7, and PIK3CA cfDNA from clinical samples by RPA-LF. Twelve cfDNA samples isolated from the serum of seven cervical cancer patients (C16, C22, C24, C27, C28, C29, and C38) and five controls (N5, N7, N26, N27, and N28) were randomly selected to compare the diagnostic performance of RPA-LF and ddPCR. (a to c) RPA-LF results for PIK3CA WT (a), HPV16 E7 (b), and HPV18 E7 (c). One microliter of cfDNA was used as a template and primer sets F2/R1, F1/R1, and F2/R2 were used to detect PIK3CA WT, HPV16 E7, and HPV18 E7, respectively. Nuclease-free water was used as no template control (NTC). Copy number of target DNA quantified by ddPCR is shown in blue text. C: control line; T: test line. (A color version of this figure is available in the online journal.)
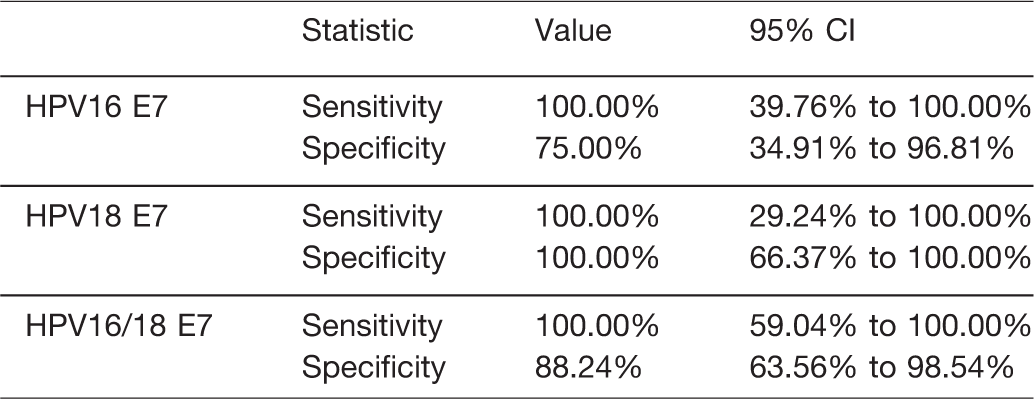
Finally, we conducted diagnostic statistics of the RPA-LF assay based on the data obtained by ddPCR as the reference. As shown in Table 4, HPV18 E7 RPA-LF gave the sensitivity and specificity of 100% while the HPV16 E7 assay was less specific, exhibiting 100% sensitivity and 75% specificity (Table 4). The combination of HPV16/18 E7 revealed 100% sensitivity and 88.24% specificity (Table 4). The agreement between RPA-LF and ddPCR assays was 83.33% (κ = 0.67) for HPV16 E7, 100% (κ = 1.0) for HPV18 E7, and 91.67% (κ = 0.81) for HPV16/18 E7, indicating good correlation between the two tests. 37 Therefore, we concluded that the RPA-LF assay was highly sensitive and could serve as a rapid HPV E7 cfDNA test for monitoring cervical cancer patients, especially in resource-limited settings.
Comparison of diagnostic statistics of HPV E7 cfDNA detection by RPA-LF as compared with the ddPCR.
Discussion
The present study evaluated the possibility of using HPV E7 cfDNA as a liquid biopsy biomarker for cervical cancer detection, as well as its ultrasensitive and rapid detection by RPA-LF. First, we employed ddPCR for dual detection of HPV16/18 E7 and the internal control PIK3CA WT. The assay was highly sensitive, with LOD of 2–2.5 copies/reaction (Figures 2, S1, and S2). The use of the reference gene enabled the rapid quantitative and qualitative assessment of cfDNA, overcoming the need for measuring total cfDNA concentration (Figure S4). HPV16/18 E7 cfDNA is tumor-specific and serves as an excellent biomarker for separating late-stage cervical cancer patients from controls (Tables 1 to 3 and Figure 3). Next, we developed rapid isothermal detection of HPV16/18 E7 and PIK3CA WT using RPA-LF. The reaction was completed within 30 min and its sensitivity was comparable to ddPCR (Figure 4). HPV16/18 E7 analysis of clinical samples by RPA-LF revealed 100% sensitivity, 88.24% specificity, and good correlation with ddPCR (κ = 0.81; Table 4 and Figure 5). Thus, we conclude that HPV E7 cfDNA is a potential surrogate blood-based biomarker for cervical cancer monitoring and its detection by RPA-LF represents a promising alternative to ddPCR for point-of-care testing in low-resource settings.
HPV is a human oncogenic virus associated with the development of numerous malignancies, including cervical, anogenital, and head and neck cancers.38,39 High-risk HPV strains have also been found in several other tumors, including lung, colon, breast, and esophageal cancer.40–43 Of the 15 high-risk genotypes, HPV16 and HPV18 account for 70% of cervical cancers worldwide. 44 Because HPV infection is a crucial step in tumorigenesis, HPV DNA is detected in 99.7% of cervical cancer tissues. 45 In over 80% of HPV-positive cervical cancers, the extrachromosomal HPV genome often integrates into cellular DNA, leading to dysregulated E6 and E7 expression. 46 Increased expression of these oncoproteins can stimulate cell proliferation, abrogate cell cycle checkpoints, and induce genetic instability, thereby promoting carcinogenesis. 47 Integration of the HPV genome usually results in the partial deletion of viral genes such as E1, E2, L1, and L2; whereas the presence of E6 and E7 is essential for all stages of tumor progression. 48 Therefore, we hypothesize that the E6/E7 genes are more tumor-specific than the rest of the viral genome.
To date, several types of circulating nucleic acids have been identified, including segments of genomic, mitochondrial, or viral DNA, RNA, and microRNA. 49 Circulating nucleic acids can be released from apoptotic and necrotic cells, or actively released by some living cells. 50 In healthy persons, a majority of cfDNA originates from apoptosis of blood nucleated cells and its level might be increased in some inflammatory conditions. 51 In cancer patients, cfDNA levels can be elevated, especially in advanced stages.52,53 Here, we observed no difference in total cfDNA levels between samples (Figure 1(c)), confirming similar findings on cfRNA. 54 While a larger cohort is required to confirm our results, the obtained data argue against the use of total cfDNA yields as a cervical cancer biomarker. Alternatively, it is possible that this discrepancy could be attributed to the leakage of genomic DNA from white blood cells since we did not use the cfDNA collection tubes minimizing genomic DNA contamination. Importantly, cfDNA fragments originating from tumor cells are reportedly shorter than those from non-malignant cells.53,55 Indeed, cfDNA in cervical cancer patients was consistently shorter than normal cfDNA (Figure 1(b)). These data are in agreement with a previous report on pancreatic cancer (locally advanced cancer, median 170 bp, range 167–173 bp vs. normal, median 176.5 bp, range 168–185 bp). 53 Although a mechanism for the increased fragmentation of tumor cfDNA is not well understood, profiling of cfDNA fragment size might serve as an additional cancer biomarker, especially for monitoring cancer patients.
Circulating tumor DNA may harbor cancer-associated molecular characteristics, such as mutations, methylation changes, and cancer-derived sequences, 56 making it a potential tool for non-invasive diagnosis and monitoring of cancer. 57 These cfDNA alterations may serve as specific tumor markers; however, an ultrasensitive test is essential as mutant DNA fragments only account for a small fraction of total cfDNA. 58 Common PIK3CA mutations in cfDNA, such as E542K and E545K, which were found in 22.2% of plasma cfDNA from cervical cancer patients, 10 appear to be very promising biomarkers.59,60 In our cohort, we detected PIK3CA mutation solely in cfDNA derived from a stage IIIB cervical cancer patient, resulting in 100% specificity but only 2.56% sensitivity (Tables 1 and S3, and Figure 2(c)). Therefore, PIK3CA E542K does not appear to be a suitable cervical cancer biomarker; however, a larger cohort should be explored to evaluate a correlation in Thai cervical cancer patients.
Besides cfDNA of cellular origin, HPV DNA can be detected also in peripheral blood.15,35,58,61 As the HPV DNA sequence is ubiquitous within tumor cells and is distinct from the human genome, circulating HPV DNA could help detect cancer and monitor the disease. 58 Nevertheless, it can be detected in only 12–25.5% of blood samples of HPV-positive cervical cancer patients based on conventional PCR.12,62 As an alternative, ddPCR offers enhanced sensitivity for detecting HPV cfDNA in patients with HPV-associated carcinomas, exhibiting a pooled sensitivity of 0.83 (95% CI, 0.71–0.91) compared with 0.21 (95% CI, 0.18–0.25) by qPCR. 14 Here, we utilized ddPCR to detect HPV16/18 E7 cfDNA from serum samples of patients with cervical cancer, CIN, and controls. As shown in Figures S1 and S2, the duplex ddPCR assays had LOD of 2.13 and 2.30 for HPV16 and HPV18 E7 (80–92 copies/mL serum), respectively, with 100% detection rate when plasmid DNAs were used as templates. The LOD can be improved by applying a larger volume of serum and concentrated cfDNA. In addition, no cross-reactivity between HPV16 and HPV18 (Figure S3) as well as no HPV DNA in human genomic DNA (Figure 2(e)) were detected. Thus, the duplex ddPCR assay was suitable for detecting poorly abundant HPV cfDNA.
To assess the quality and quantity of samples, we included the PIK3CA and p16INK4A genes as internal controls. The average concentration of extracted cfDNA is very low, usually less than 10 ng/mL plasma in healthy individuals. 63 Moreover, liquid biopsy samples including blood contain various PCR inhibitors, such as heparin, immunoglobulin G, or hemoglobin, potentially lowering amplification efficiency in ddPCR.64,65 As shown in Tables 1 and S3, we were able to detect PIK3CA (16–591 copies/reaction) and p16INK4A (8.6–455 copies/reaction) in all samples, indicating that the extracted cfDNA was amplifiable and in sufficient amount for ddPCR. Furthermore, their copy numbers were not dependent on the disease stage and were proportional to the total cfDNA concentration as quantified by the Fragment Analyzer (Figures 3(a) and (b), S4A and B). PIK3CA WT levels correlated linearly with p16INK4A (Figure S4(C)), confirming their suitability as reference genes for cfDNA analysis. Several previous studies on circulating HPV DNA did not include either reference gene detection or total cfDNA measurement. Thus, it is possible that false-negatives were detected due to insufficient cfDNA or contamination with PCR inhibitors. To ensure data accuracy, we suggest the incorporation of reference genes in the assay.
Based on ddPCR analysis, HPV E7 cfDNA was detected only in stage III/IV cervical cancer patients (Figure 3(c) and (d)), with HPV16 being the most commonly identified genotype (Table 1). This confirms earlier reports of HPV16 as the more prevalent strain in both cervical and blood samples.16,18 Here, sensitivity was 20.51%, 10.26%, and 30.77% when using HPV16 E7, HPV18 E7, or HPV16/18 E7, respectively (Table 2). A combination of HPV16 and HPV18 E7 yielded AUC values of 0.65 and 0.74 when separating healthy controls from cervical cancer and stage III/IV cervical cancer, respectively (Table 3, and Figure 3(f) and (g)). It should be noted that, unlike other studies,15,35,58 we did not pre-select patients with HPV16- or HPV18-associated carcinomas. Thus, our data would more likely reflect the probability of using HPV16/18 E7 cfDNA in identifying cervical cancer patients from a general population, rather than sensitivity towards HPV16/18 E7 cfDNA detection from HPV16/18-positive cancer patients. Nevertheless, we found that the diagnostic performance of HPV cfDNA from our study was in good accordance with previous meta-analysis studies, showing pooled sensitivity and specificity of 0.27 and 0.94, respectively. 14
One obvious way to improve the sensitivity of HPV E7 as a biomarker is to include more high-risk HPV strains in the assay. Kang et al. demonstrated that including 12 other HPV genotypes in cfDNA analysis improved sensitivity in cervical cancer detection without a loss of specificity. 35 Moreover, analysis of frequent HPV oncogenic types in Thai women revealed that HPV16 and 18 accounted for only 43.6% of cervical cancer cases. Addition of eight high-risk genotypes (58, 52, 45, 33, 35, 51, 39, and 31) could improve coverage to 95%. 66 Another means to enhance the sensitivity of HPV E7 cfDNA is to include another sequence as a surrogate biomarker. Cheung et al. increased sensitivity from 55.8% to 61.6% by combining ddPCR testing for both E7 and L1. 58 Therefore, we suggest that the limited sensitivity of HPV16/18 E7 cfDNA in identifying cervical cancer patients can be overcome by adding multiple high-risk HPV genotypes as well as another HPV target gene to increase coverage.
Despite the potential of HPV E7 cfDNA as a cervical cancer biomarker, there are still practical limitations in detecting patients with low disease burden. Our study shows that HPV E6/E7 cfDNA is 100% specific for cervical cancer as none of the patients with precancerous lesions and controls was positive (Tables 1 and 2). This is in agreement with previous reports showing that HPV E6/E7 cfDNA was not detectable in healthy blood donors or patients diagnosed with CIN.12,15,18,35 It is possible that no HPV E6/E7 DNA is released at a detectable level in the blood of non-cancerous individuals. Due to its moderate sensitivity and strong association with advanced-stage cancer, we suggest that HPV E6/E7 cfDNA is a more suitable biomarker for monitoring cervical cancer patients with an already confirmed diagnosis, rather than being used for screening and early detection of cervical cancer.
Paradoxically, Cocuzza et al. reported detecting HPV E1/E2 cfDNA (types 16, 18, 31, 33, 45, 51, and 52) in 33.3% (8/24) of atypical squamous cells of unknown significance, 42.3% (22/52) low-grade squamous intraepithelial lesions, 66.7% (12/18) high-grade squamous intraepithelial lesions, and 0% in the control group. 61 The discrepancy may arise from the disruption of the E2 gene during the integration of the HPV genome. 67 Thus, detection of E2 DNA might correspond predominantly to the positive episomal HPV form; 35 whereas E6/E7 genes can be found in either episomal or integrated form. We, therefore, suggest the development of blood-based cervical cancer detection tests that incorporate E1/E2 and E6/E7 as targets to increase the likelihood of detecting both low-grade/precancerous dysplasia and cervical cancer.
Although HPV cfDNA serves as a promising biomarker for diagnosing and monitoring cervical cancer, detection of circulating nucleic acids remains a challenge and requires the use of qPCR or, preferably, ddPCR. 14 These methods are time-consuming and require both expensive instruments and experienced operators, which renders them unsuitable for facilities with limited resources, especially in developing countries. In this study, we developed a fast, highly sensitive, PCR-free method based on RPA-LF. To our knowledge, this is the first report utilizing RPA-LF for circulating HPV DNA detection. The RPA reaction was carried out at a constant temperature and was completed in only 20 min. RPA amplicons could be detected by the naked eye using the lateral flow assay by dipping the strip in the product–buffer mixture for another 7 min. RPA-LF is thus much faster than qPCR and LAMP assays (1 h), Clinichip HPV™ (2.5 h), and ddPCR (>2 h).33,68,69 Another unique advantage of RPA is its high sensitivity, capable of amplifying as few as 1–10 copies of target DNA.70,71 Indeed, the LOD was 5–10 copies/reaction (Figure 4), similar to the HPV16/18 RPA assay described by Ma et al. 33 The sensitivity is also comparable to qPCR and ddPCR.72,73 As with ddPCR, we utilized PIK3CA as a reference gene to ensure that cfDNA was sufficient and amplifiable by the RPA reaction (Figure 4). Finally, we strongly argue in favor of incorporating an internal control in RPA-based analysis due to the limited quantity and potential contamination of tumor-derived cfDNA with PCR inhibitors. 74
Analysis of clinical specimens demonstrated that the RPA-LF assay was sensitive enough to detect cfDNA samples including the PIK3CA WT reference gene and there was no cross-reactivity between HPV subtypes (Table 1 and Figure 5). In addition, the HPV18 E7 RPA-LF assay produced identical results as ddPCR (100% sensitivity, 100% specificity). RPA-LF correctly identified all HPV16 E7-positive samples; however, we observed some false-positives (100% sensitivity, 75% specificity; Table 4 and Figure 5). False-positivity might be caused by unexpected amplification or primer dimers. 75 Further optimization of an appropriate primer/probe combination is likely to improve assay specificity. 76 Regardless, our data revealed a satisfactory correlation (83.33–100%) between RPA-LF and ddPCR for HPV16 E7, HPV18 E7, and HPV16/18 E7. Hence, RPA-LF provides an alternative method to ddPCR for rapid and simple detection of HPV E7 cfDNA for cervical cancer monitoring. The RPA-LF assay could assist in clinical decision making by qualified health care professionals in accordance with professional guidelines.
In summary, HPV E7 cfDNA serves as a surrogate blood-based cervical cancer biomarker with excellent specificity and moderate sensitivity. It may serve for monitoring patients with a confirmed diagnosis, but not well suited for screening and early detection of cervical cancer. Moreover, HPV E7 cfDNA can be detected using a rapid, easy-to-visualize, highly sensitive, and thermocycler-free RPA-LF assay, which exhibits a good correlation with ddPCR. Further studies are required to test the diagnostic performance of HPV E7 cfDNA from multiple high-risk genotypes in a larger cohort. This should be accompanied by the development of an RPA-LF-based assay applicable for highly sensitive multiplex detection of HPV for point-of-care testing in resource-limited areas.
Supplemental Material
sj-pdf-1-ebm-10.1177_1535370220978899 - Supplemental material for Rapid and ultrasensitive detection of circulating human papillomavirus E7 cell-free DNA as a cervical cancer biomarker
Supplemental material, sj-pdf-1-ebm-10.1177_1535370220978899 for Rapid and ultrasensitive detection of circulating human papillomavirus E7 cell-free DNA as a cervical cancer biomarker by Phetploy Rungkamoltip, Sasithon Temisak, Kitiya Piboonprai, Deanpen Japrung, Pattanapong Thangsunan, Saranya Chanpanitkitchot, Woraphot Chaowawanit, Nutthaporn Chandeying, Siriwan Tangjitgamol and Tawin Iempridee in Experimental Biology and Medicine
Footnotes
AUTHORS’ CONTRIBUTIONS
TI supervised the study, designed experiments, analyzed and interpreted data, and wrote the manuscript. PR, ST, and KP performed experiments, analyzed data, and drafted the methodology and results. DJ provided suggestions and helped coordinate the study. PT, SC, WC, NC, and ST collected samples and analyzed clinical data. All authors approved the content of this manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
ETHICAL APPROVAL
Written informed consent was obtained from all volunteers and the study was approved by the Vajira Institutional Review Board, Faculty of Medicine, Vajira Hospital, Navamindhadhiraj University (COA 091/2019).
FUNDING
This work was supported by the National Nanotechnology Center, Thailand [grant numbers P1750034, P1851699, and P1750162].
SupplementAL MATERIAL
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.