
Abstract
Deregulation of HOX transcription factor family has frequently been observed in multiple human cancers; however, their role in nasopharyngeal carcinoma remains largely unclear. In the present study, we found that HOX gene family is consistently upregulated in nasopharyngeal carcinoma and identified HOXA10 as one of the mostly upregulated HOX genes. Importantly, we show that HOXA10 overexpression is associated with transcriptional activation of multiple oncogenes essential for nasopharyngeal carcinoma carcinogenesis, including S-phase kinase-associated protein 2 (SKP2), calcium/calmodulin-dependent protein kinase kinase 2 (CAMKK2), and matrix metalloproteinase 1 (MMP1). Mechanistically, the overexpression of SKP2 induces the degradation of cell cycle inhibitor p27, leading to rapid cell cycle progression and cell proliferation. The overexpression of CAMKK2 is associated with enhanced mTOR signaling activity to meet the increased demand for proteins synthesis in rapid growing nasopharyngeal carcinoma cells. Moreover, MMP1 overexpression facilitates nasopharyngeal carcinoma cell migration and invasion and contributes to cancer metastasis and progression. We thus concluded that HOXA10 overexpression promotes the growth and metastasis of nasopharyngeal carcinoma by transcriptionally activating various oncogenic pathways.
Impact statement
Abnormal expression of HOX transcription factor family has frequently been observed in various human cancers. However, its function remains largely unknown in nasopharyngeal carcinoma (NPC). Here we report a novel finding that HOXA10 is frequently upregulated in NPC, and can promote the rapid growth and metastasis of NPC cells by transcriptionally regulating downstream targets, including SKP2, CAMKK2, and MMP1. These findings expanded our knowledge on the tumorigenic function of HOXA10 in NPC and also provided novel insights into the development of new therapies.
Introduction
Nasopharyngeal carcinoma (NPC) is a type of head and neck cancer originating in the nasopharynx. Although the overall incidence of NPC is very low in most parts of the world (< 1 case per 100,000 individuals per year), it occurs at high frequency in Southern China and Southeast Asia (25–30 cases per 100,000 individuals per year) and is at high risk of metastasis. 1 Advances in imaging techniques and chemoradiotherapy strategies over the past decade have largely improved the diagnosis and treatment of NPC. However, since the majority of the NPC cases (95.5%) present at advanced disease stage (stages III and IV), the overall survival rate of NPC remains as low as 61% and less than 50% of the patients with metastatic NPC stay alive five years after diagnosis. 2 Therefore, there is an urgent need to understand the molecular pathogenesis of NPC and develop novel therapeutic methods to improve patient outcomes.
HOX genes, a subset of homeobox genes, are transcription factors involved in the morphogenesis and cell differentiation during embryonic development. Deregulated HOX gene expression in adult tissues is frequently observed in multiple human cancers, and has been characterized as a causal factor in cancer initiation and progression.3,4 In the present study, by analyzing public transcriptome dataset, we identified an overall upregulation of HOX family in NPC, and we found that HOXA10 is among the top upregulated genes. HOXA10 has been found to participate in various aspects of tumor biology. In gastric cancer, HOXA10 transcriptionally induces the expression of anti-apoptotic BCL2, which subsequently inhibits apoptosis and promotes cancer cell survival. 5 HOXA10 also upregulates MMP3 expression in bladder cancer cells, leading to the rapid cell proliferation and invasion. 6 Moreover, there is evidence suggesting a strong correlation between HOXA10 and histone deacetylase 1 (HDAC1) in hepatocellular carcinoma. Elevated HDAC1 level as a result of HOXA10 overexpression deacetylates p53 and represses p53-mediated cell cycle arrest and apoptosis, implying a possible HOXA10 regulatory mechanism involving protein post-translational modifications. 7 In addition to activating transcription, HOXA10 has also been characterized as an expression inhibitor for a group of genes. HOXA10 suppresses the transcription of fatty acid synthase (FASN) in prostate cancer cell by forming a complex with androgen receptor (AR) and preventing AR binding to FASN gene promoter. 8 HOXA10 also downregulates the expression of Snail in endometrial carcinoma, which causes the aberrant expression of genes involved in epithelial-mesenchymal transition. 9
All these prior findings suggested the association of HOXA10 with cancer development. However, there is still a lack of knowledge regarding HOXA10 in NPC carcinogenesis. In the present study, we thus aim to explore the HOXA10-dependent mechanism in NPC pathogenesis.
Materials and methods
Cell culture, gene overexpression, and gene knockdown
Human NPC cell lines, CNE-1, and HK-1 were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS) and were maintained at 37°C in a humidified atmosphere with 5% CO2. NPC cells with specific gene overexpression or knockdown were constructed using lentivirus expression vector system. To overexpress HOXA10, lentiviral vectors expressing HOXA10 (Lv-HOXA10) and negative control (Lv-NC) were constructed. For specific gene silencing, short hairpin RNA (shRNA) against HOXA10 (sh-HOXA10), CAMKK2 (sh-CAMKK2), MMP1 (sh-MMP1) and scramble control (sh-Ctrl) were cloned into lentiviral vectors. Constructed vectors were transfected into 293 T cells along with packaging vectors. Recombinant lentiviruses carrying target genes or shRNAs were produced from cell culture media and were then used to infect target NPC cells.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from NPC cells using RNeasy Kit (QIAGEN). Samples were then treated with DNase I and were reversely transcribed into cDNA using high-capacity cDNA reverse transcription kit (Thermo). qPCR amplification and analysis were performed using PowerTrack SYBR Green Master Mix (Applied Biosystems) along with gene-specific primers (Table 1) on the 7900HT real-time PCR system (Applied Biosystems). Gene expression was normalized to GAPDH and was quantified using the 2−ΔΔCt method.
Primers used in the study.

HOXA10: Homeobox A10; SKP2: S-phase kinase-associated protein 2; CAMKK2: calcium/calmodulin-dependent protein kinase kinase 2; MMP1: matrix metalloproteinase 1; GAPDH: glyceraldehyde 3-phosphate dehydrogenase; TFBS: transcription factor binding site.
Chromatin immunoprecipitation assay
NPC cells were treated with 1% formaldehyde for cross-linking, followed by glycine treatment to stop the reaction. The cells were then incubated with lysis buffer and nuclei were collected by centrifugation. After that, chromatin was sheared into small fragment by sonication and incubated with antibody against HOXA10 or IgG overnight. Immunocomplexes were harvested by protein G beads, treated by proteinase K, and de-crosslinked by incubation at 65°C overnight. DNA was purified by phenol-chloroform extraction and was used for PCR analysis (Table 1).
Cell proliferation assay
Cell counting kit 8 (CCK-8) (Sigma) was used to assess cell proliferation. Briefly, NPC cells were plated in 96-well cell culture plates and were incubated with CCK-8 solution for 2 h at different time points (0, 24, 48, 72 h) according to manufacturer's instructions. The absorbance was read at 450 nm.
Cell colony formation assay
NPC cells were plated in 12-well cell culture plates and were incubated at 37°C in a humidified atmosphere with 5% CO2 for 14 days to allow colony formation. Cell colonies were fixed with methanol and stained with crystal violet. The colonies were then counted under microscope.
Nascent protein synthesis assay
Click-IT HPG Alexa Fluor Protein Synthesis Assay Kit (Thermo) was used to quantify nascent protein synthesis. In brief, NPC cells were plated in 6-well cell culture plates. After 24-h incubation, cell culture media were replaced with an L-methionine-free DMEM medium supplemented with L-homopropargylglycine (HPG), a methionine analog containing an alkyne moiety. After incubation at 37°C for another 30 min, newly translated polypeptides with incorporated L-HPG signal were detected by flow cytometry.
Wound healing assay
NPC cells were plated in 6-well cell culture plates and were incubated at 37°C in a humidified atmosphere with 5% CO2. At ∼95% confluence, a sterile 100 μL pipette tip was used to make a scratch in cell monolayer, and the detached cells were removed by washing with ice-cold PBS. After incubation at 37°C for another 24 h, wound width was measured under microscope.
Transwell invasion assay
Transwell invasion assay was performed using Transwell chambers coated with Matrigel matrix. NPC cells were suspended in FBS-free DMEM medium were seeded into the upper chamber of the Transwell. The lower chamber was filled with DMEM medium containing 20% FBS as a chemoattractant. After incubation at 37°C for 24 h, cells staying in the upper chamber were gently removed with cotton swabs, and cells on the underside of the upper chamber were fixed with methanol and stained with 0.2% crystal violet.
Immunoblotting assay
NPC cells were lyzed using radioimmunoprecipitation assay buffer (RIPA). Total proteins were extracted and were quantified using BCA assay kit (Bio-Rad); 20 μg of total proteins from each sample was loaded and separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and were then transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were blocked with 5% non-fat milk and were incubated with antibodies against HOXA10 (Santa Cruz, sc-271428), SKP2 (CST, 4358), CAMKK2 (Thermo, PA5-78909), MMP1 (Abcam, ab38929), p27 (Abcam, ab32034), Ki-67 (Abcam, ab16667), phospho-mTOR (CST, 2971), mTOR (CST, 2972), phospho-S6K (CST, 9205), S6k (CST, 9202), and β-actin (Abcam, ab115777). After that, the membranes were incubated with horseradish peroxidase (HRP)-conjugated secondary antibody, and protein signal was detected using enhanced chemiluminescence detection kit (Thermo).
Statistical analysis
All experiments were performed in triplicates. Data were expressed as mean ± SD and was compared between different groups using Student’s t test with P < 0.05 considered as statistically significant. Statistical analysis was performed with SPSS software (IBM).
Results
HOX family is coordinately upregulated in NPC
To determine HOX family gene expression in NPC, we obtained four public datasets comparing the transcriptome of NPC versus normal controls (GSE12452, GSE13597, GSE53819, GSE61218).10–13 Using gene set enrichment analysis (GESA), we found that HOX family members were concordantly activated in NPC (Figure 1(a)). Notably, HOXA10 was among the top of genes and was upregulated in all the four NPC datasets (Figure 1(b) and (c)), indicating a putative role of HOXA10 in NPC pathogenesis.
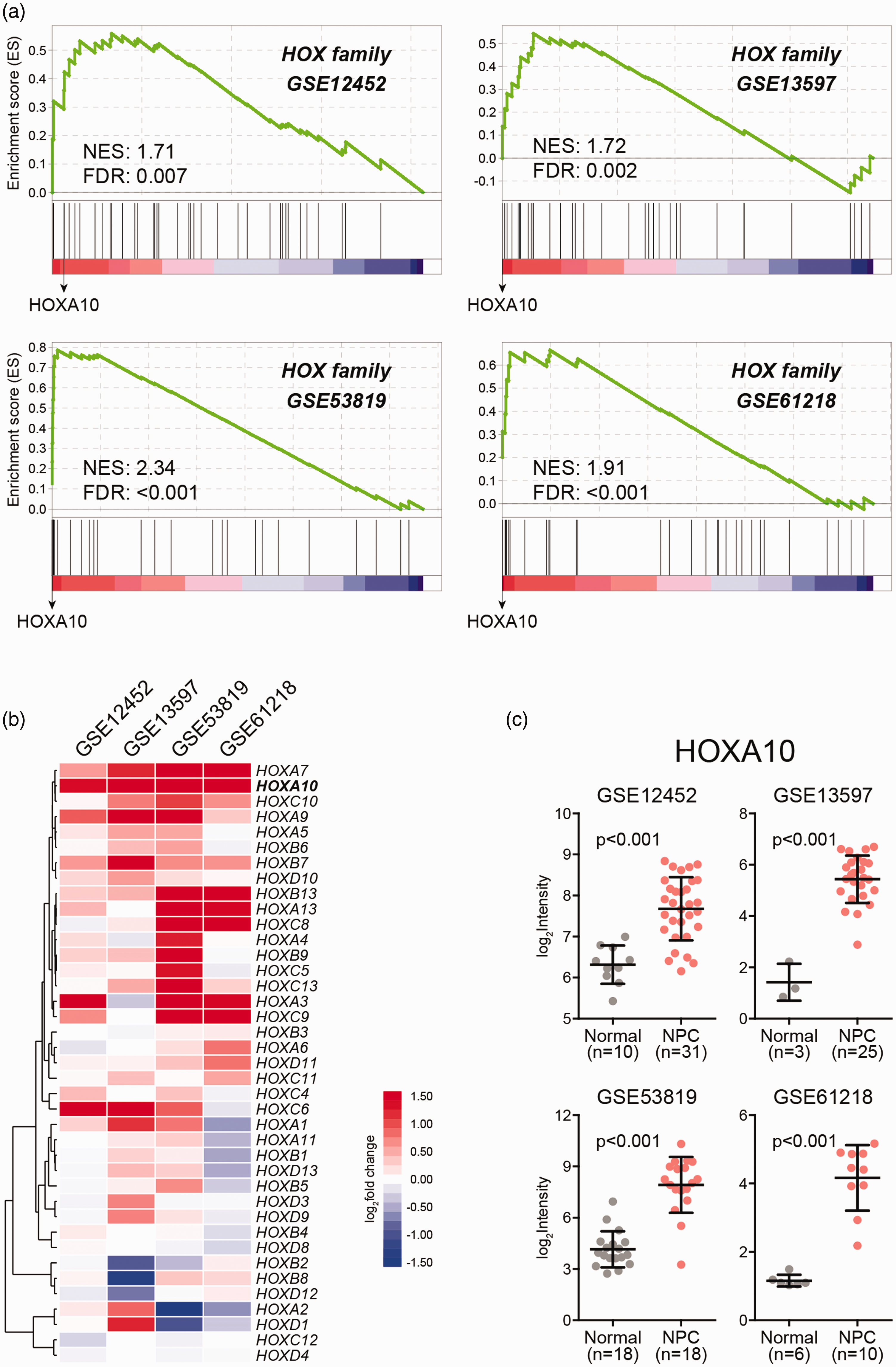
HOX genes are coordinately up-regulated in NPC. (a) Gene set enrichment analysis of HOX gene family expression change in NPC using public transcriptome datasets (GSE12452, GSE13597, GSE53819, GSE61218). (b) Hierarchical clustering of HOX gene fold change. (c) HOXA10 expression in NPC and normal tissues. (A color version of this figure is available in the online journal.)
SKP2, CAMKK2, and MMP1 are possible HOXA10 targets
Given the role of HOXA10 as a transcription factor, we asked what downstream target genes could be affected and contribute to NPC initiation and progression. To this end, we used JASPAR transcription factor database to identify HOXA10 targets from the genes overexpressed in NPC. As shown in Figure 2(a) to (d), 331 genes were upregulated in all the four NPC datasets, of which S-phase kinase-associated protein 2 (SKP2), calcium/calmodulin-dependent protein kinase kinase 2 (CAMKK2), and matrix metalloproteinase 1 (MMP1) were predicated as possible HOXA10 targets with strong HOXA10 binding sites (TFBSs) in their promoter regions (Figure 3(a) and (b)).
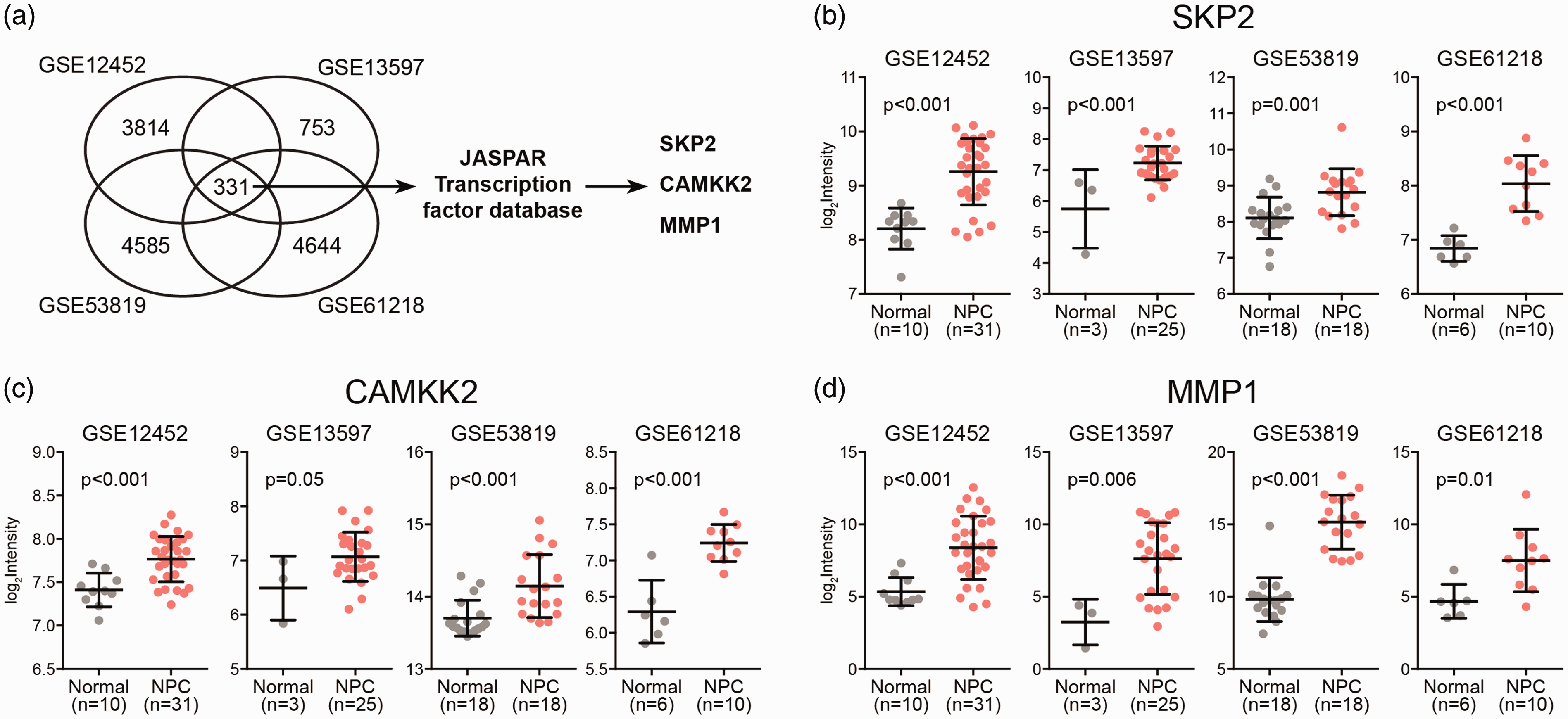
SKP2, CAMKK2, and MMP1 are possible HOXA10 targets. (a) JASPAR transcription factor analysis of the common genes up-regulated in different NPC transcriptome datasets (GSE12452, GSE13597, GSE53819, GSE61218). The expression level of SKP2 (b), CAMKK2 (c), and MMP1 (d) in NPC and normal tissues. (A color version of this figure is available in the online journal.)
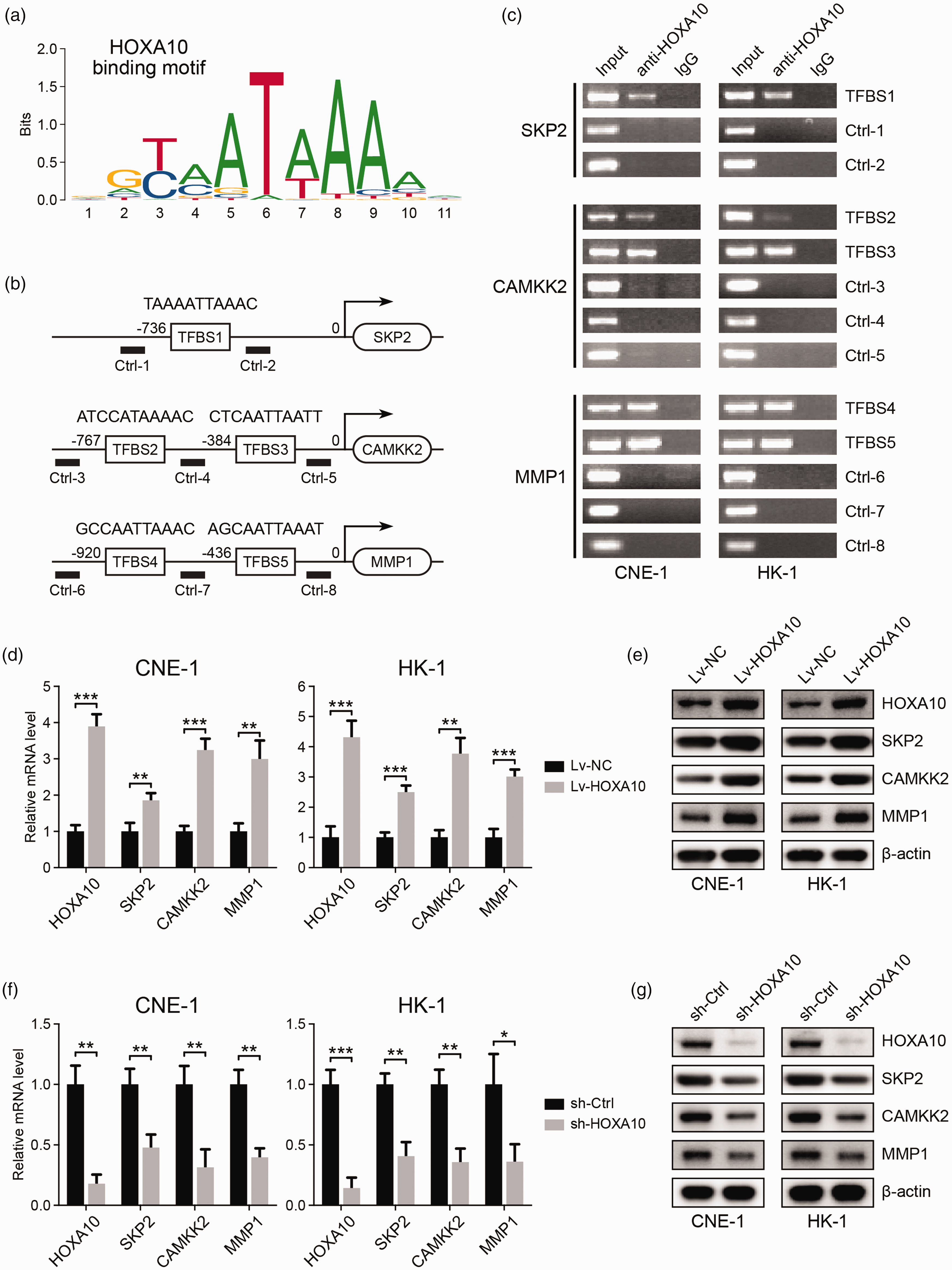
HOXA10 binds to the promoter regions of SKP2, CAMKK2, and MMP1 and regulates their expression. (a) HOXA10 binding motif. (b) The predicted HOXA10 binding sites (TFBSs) in the promoter regions of SKP2, CAMKK2, and MMP1. (c) ChIP-PCR analysis of HOXA10 binding to the promoter regions of SKP2, CAMKK2, and MMP1 in NPC cell lines. (d) mRNA expression of HOXA10, SKP2, CAMKK2, and MMP1 in NPC cells with HOXA10 overexpression. (e) Protein expression of HOXA10, SKP2, CAMKK2, and MMP1 in NPC cells with HOXA10 overexpression. (f) mRNA expression of HOXA10, SKP2, CAMKK2, and MMP1 in NPC cells with HOXA10 knockdown. (g) Protein expression of HOXA10, SKP2, CAMKK2, and MMP1 in NPC cells with HOXA10 knockdown. *P < 0.05, **P < 0.01, ***P < 0.001 by Student’s t-test. (A color version of this figure is available in the online journal.)
We then used ChIP-PCR to analyze the interaction of HOXA10 with SKP2, CAMKK2, and MMP1 promoters in two NPC cell lines, CNE-1 and HK-1. As shown in Figure 3(c), we found that HOXA10 can directly bind to the predicted binding sites in these genes. We then overexpressed HOXA10 in NPC cell lines, and the expression of SKP2, CAMKK2, and MMP1 was subsequently upregulated at both mRNA and protein levels (Figure 3(d) and (e)). Consistently, knockdown of HOXA10 expression resulted in the downregulation of these genes (Figure 3(f) and (g)). These results suggested that SKP2, CAMKK2, and MMP1 are the targets of transcription factor HOXA10, which may mediate the oncogenic functions of HOXA10 in NPC.
HOXA10 promotes NPC cell proliferation through upregulating SKP2
SKP2 is an ubiquitin ligase participating in cell cycle control. The primary target of SKP2 is p27, an inhibitor of cell cycle process. 14 As shown in Figure 4(a) and (b), HOXA10 overexpression induced SKP2 upregulation and SKP2-mediated p27 degradation through ubiquitin-mediated mechanism, which may eventually contribute to cell cycle progression and rapid cell proliferation. Consistent with this, the expression of cell proliferation marker, Ki-67, was potently increased in NPC cells overexpressing HOXA10 (Figure 4(b)). Moreover, cell growth rate was largely enhanced and more colonies were formed following HOXA10 overexpression (Figure 4(c) to (f)). In contrast, when we used SMIP004 to inhibit SKP2 in cells overexpressing HOXA10, more p27 was accumulated in the cells (Figure 4(b)), leading to the suppression of cell cycle and cell proliferation. This was evidenced by decreased Ki-67 expression (Figure 4(b)) as well as reduced cell growth rate and colony formation ability (Figure 4(c) to (f)) in SMIP004-treated cells. These findings suggested that HOXA10 can promote NPC cell proliferation by inducing SKP2 expression (Figure 4(a)).
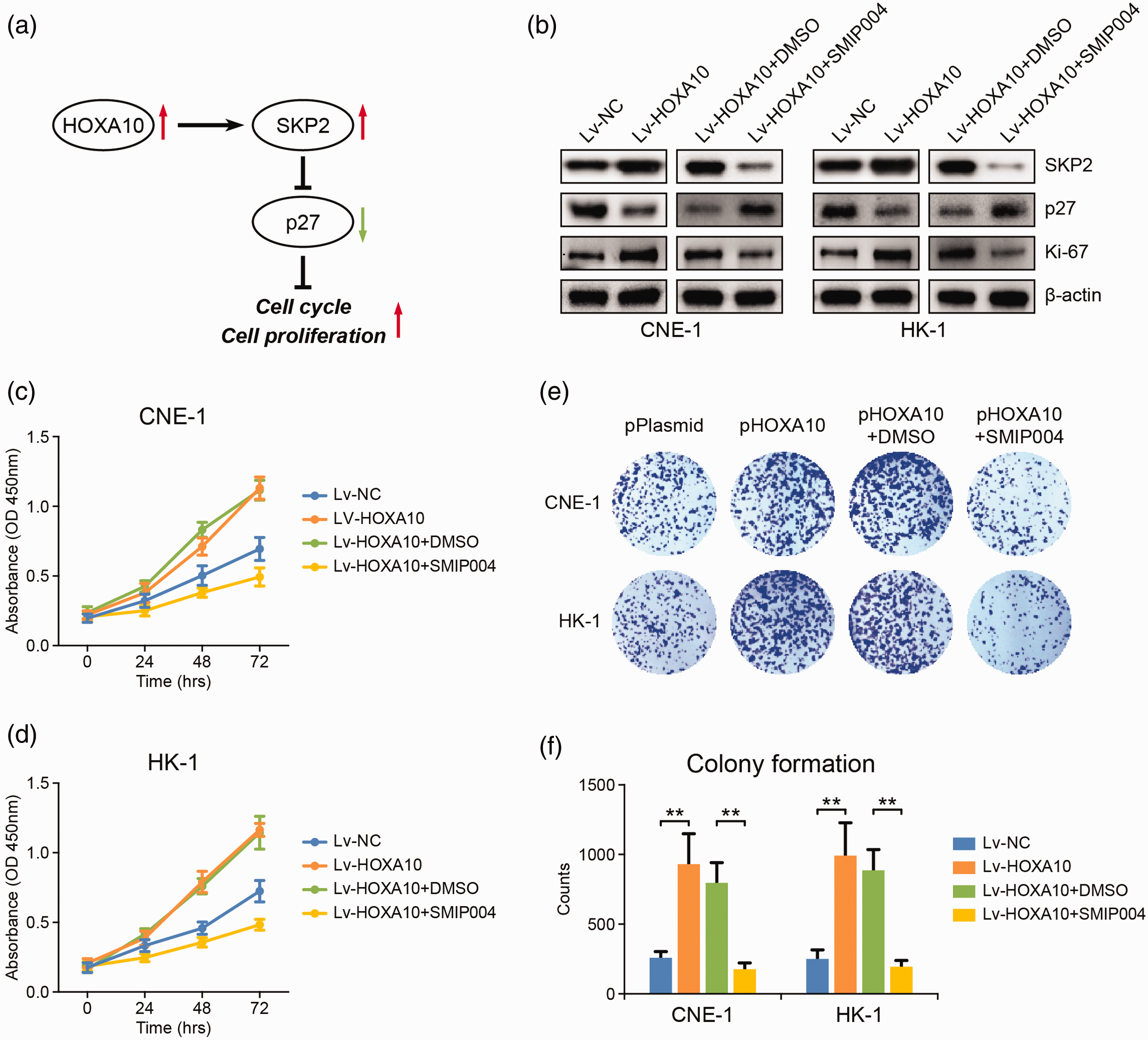
HOXA10 promotes cell cycle progression and cell proliferation through up-regulating SKP2. (a) The possible pathway through which HOXA10 promotes cell cycle progression and NPC cell proliferation. (b) The influence of HOXA10 overexpression and SKP2 inhibition (40 μM SMIP004, 24 h) on the expression of SKP2, p27, and Ki-67 in NPC cells. (c, d) The influence of HOXA10 overexpression and SKP2 inhibition on the proliferation of NPC cells. (e, f) The influence of HOXA10 overexpression and SKP2 inhibition on the colony formation ability of NPC cells. **P < 0.01 by Student’s t-test. (A color version of this figure is available in the online journal.)
HOXA10 promotes protein synthesis through upregulating CAMKK2
Our analysis above also identified CAMKK2 as a HOXA10 target in NPC cells (Figure 3). CAMKK2 is an oncogene upregulated in various types of cancers. Its tumorigenic function involves activating the mammalian target of rapamycin (mTOR) signaling pathway. 15 In NPC cells, we found that HOXA10 overexpression was associated with CAMKK2 upregulation and mTOR activation, while knockdown of CAMKK2 expression resulted in dephosphorylation and deactivation of mTOR (Figure 5(a) to (c)), suggesting that HOXA10 can activate mTOR through upregulating CAMKK2.
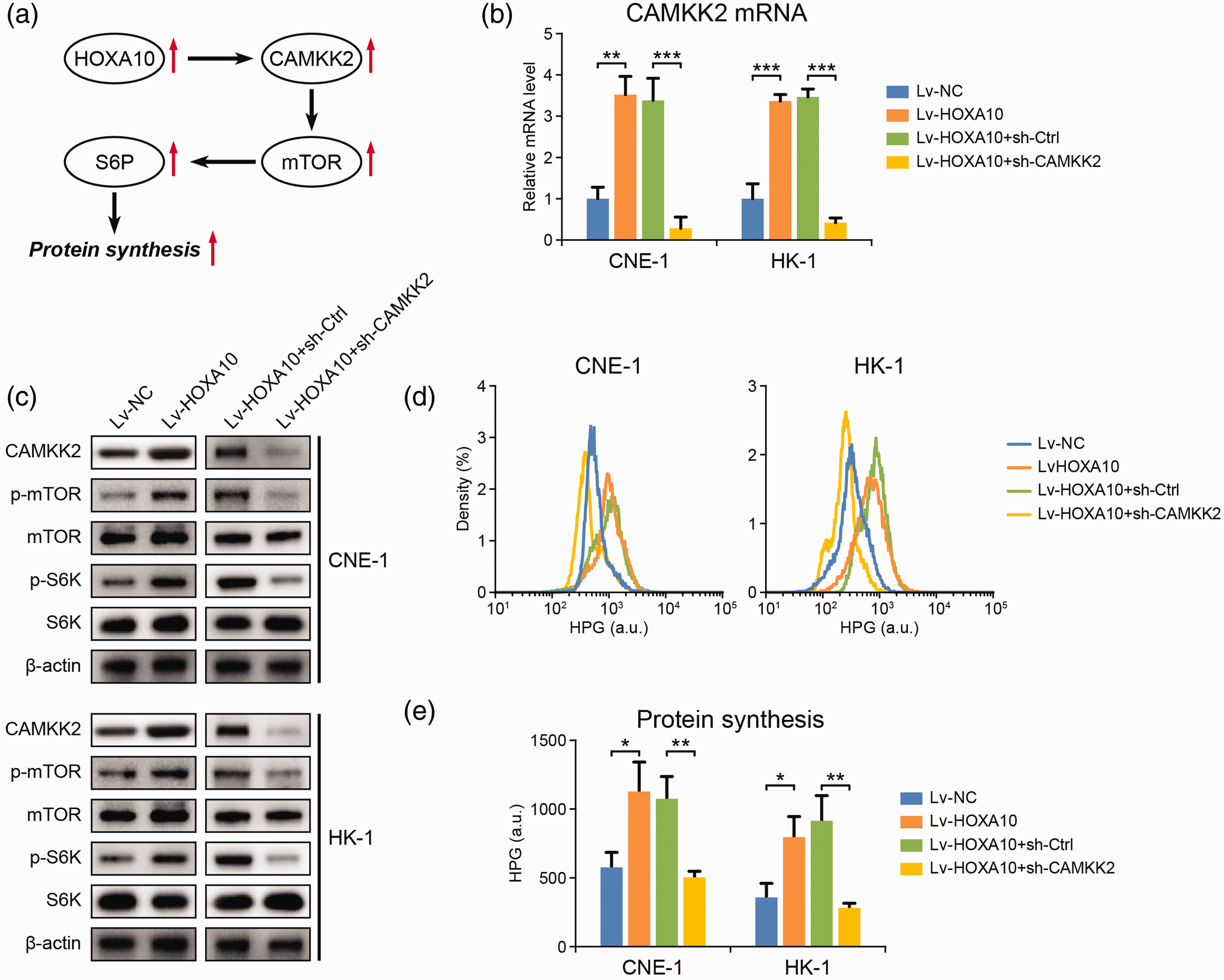
HOXA10 promotes protein synthesis through up-regulating CAMKK2. (a) The possible pathway through which HOXA10 promotes protein synthesis in NPC cells. (b) mRNA level of CAMKK2 in NPC cells with HOXA10 overexpression and with CAMKK2 knockdown. (c) The influence of HOXA10 overexpression and CAMKK2 knockdown on the expression of CAMMK2 and the activation of mTOR and S6K in NPC cells. (d, e) The influence of HOXA10 overexpression and CAMKK2 knockdown on protein synthesis rate in NPC cells. *P < 0.05, **P < 0.01, ***P < 0.001 by Student’s t-test. (A color version of this figure is available in the online journal.)
mTOR is a key regulator of protein synthesis, which is essential to support normal cell growth and function. 16 Here, we found that ribosomal protein S6 kinase (S6K) was phosphorylated and activated following HOXA10/CAMKK2-induced mTOR activation (Figure 5(c)) and more newly translated proteins were detected in NPC cells with HOXA10 overexpression (Figure 5(d) and (e)), consistent with enhanced protein synthesis. In contrast, deactivating mTOR through CAMKK2 knockdown resulted in S6K deactivation (Figure 5(c)) and proteins synthesis inhibition (Figure 5(d) and (e)). We thus concluded that, in NPC cells, HOXA10 could promote protein synthesis to fuel the rapid cancer cell proliferation through the CAMKK2/mTOR axis (Figure 5(a)).
HOXA10 promotes NPC metastasis through upregulating MMP1
In addition to SKP2 and CAMKK2, MMP1 was also identified as a potential HOXA10 target in NPC (Figure 3). MMP1 is an enzyme responsible for extracellular matrix degradation, which is crucial for cancer cell metastasis. In NPC cells, HOXA10 overexpression largely induced MMP1 expression (Figure 6(a) to (c)). As revealed by wound healing and transwell assays, cell migration and invasion abilities were also dramatically increased by HOXA10 (Figure 6(d) and (e)), indicating enhanced cancer metastasis. All these alterations were reversed by MMP1 knockdown (Figure 6(d) and (e)), suggesting that HOXA10 could promote NPC cell metastasis by upregulating MMP1 expression.
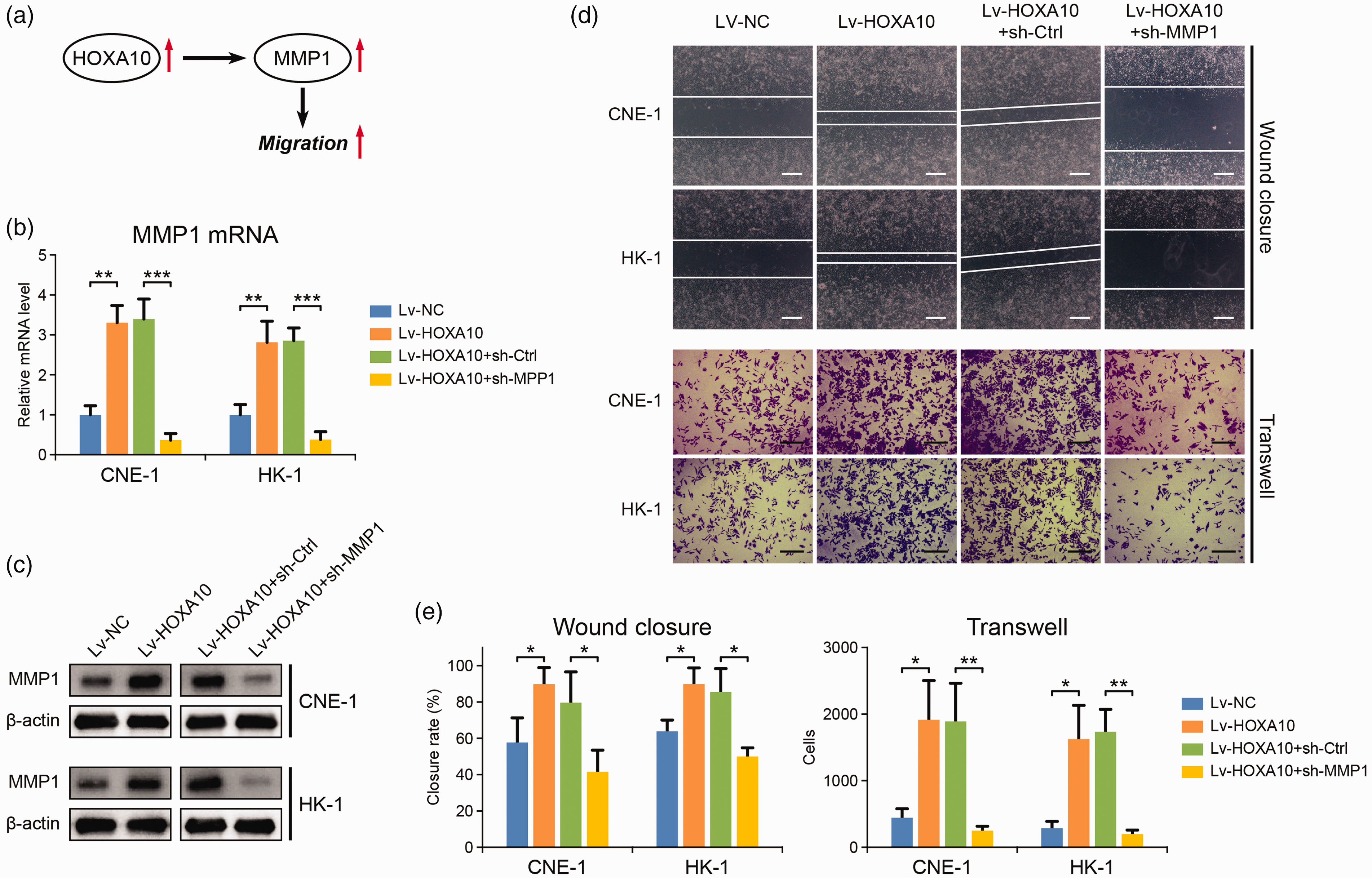
HOXA10 promotes NPC metastasis through up-regulating MMP1. (a) The possible pathway through which HOXA10 promotes NPC metastasis. (b) mRNA level of MMP1 in NPC cells with HOXA10 overexpression and with MMP1 knockdown. (c) Protein expression of MMP1 in NPC cells with HOXA10 overexpression and with MMP1 knockdown. (d, e) Wound closure and transwell assays of the influence of HOXA10 overexpression and MMP1 knockdown on the migration and invasion ability of NPC cells. The scale bar is 400 μm for wound closure assay and is 100 μm for transwell assay. *P < 0.05, **P < 0.01, ***P < 0.001 by Student’s t-test. (A color version of this figure is available in the online journal.)
Discussion
HOXA10 is a member of HOX transcription factor family initially recognized to regulate embryonic development. Recently, aberrant HOXA10 expression has been frequently reported in various human cancers and is associated with disease stages and prognosis. HOXA10 overexpression is observed in oral squamous cell carcinoma, gastric cancer, bladder cancer, and hepatocellular carcinoma, which is considered to facilitate cancer progression by activating cell proliferation, inhibiting apoptosis and promoting cell metastasis.5–7,17 In addition, HOXA10 also shows anti-tumor properties with downregulated expression detected in endometrial cancer, testicular cancer, and prostate cancer,9,18–20 implying the complexity of HOXA10 function in cancer biology. In the present study, through systematic transcriptome analysis, we identified HOXA10 as an oncogene overexpressed in NPC tissues. We also provided evidence that HOXA10 promotes NPC growth and metastasis through transcriptionally regulating downstream targets, including SKP2, CAMKK2, and MMP1. Specifically, HOXA10 induces SKP2-mediated p27 degradation to accelerate cell cycle progression and cell proliferation. HOXA10 activates protein synthesis via CAMKK2/mTOR axis to support rapid NPC cell growth. HOXA10 also facilitates NPC metastasis by upregulating MMP1 expression. All these findings expanded our knowledge on HOXA10 function in NPC pathogenesis and provide HOXA10 as a promising therapeutic target for NPC treatment. It has been reported that microRNA135a and microRNA135b regulate HOXA10 expression. 21 Future studies are thus needed to evaluate the efficacy of these microRNAs in inhibiting HOXA10 and HOXA10-mediated tumorigenic pathway in NPC. Moreover, developing novel HOXA10 inhibitors is also required to target HOXA10-overexpressed NPC.
In addition to HOXA10, our study revealed a consistent upregulation of the HOX gene family in NPC. For example, HOXA7 was upregulated in three out of the four NPC transcriptome datasets (Figure 1). HOXA7 is associated with müllerian-like differentiation of epithelial ovarian tumors and also plays a critical role in liver cancer metastasis by activating Snail expression.22,23 It is thus possible that the aberrant HOXA7 expression in NPC may also contribute to NPC progression, which deserves further investigation. Other than that, many other HOX genes are also known to regulate various cancer developments. In acute leukemia, HOXA9 and HOXC6 are frequently upregulated and drive the leukemic transformation, resulting in poor prognosis.24,25 In breast cancer, HOXB13 is responsible for the generation of drug resistance in estrogen receptor-positive tumors,26,27 while the up-regulation of HOXB7 confers epithelial-mesenchymal transition, induces angiogenesis, and is associated with bone metastasis. 28 In addition to the oncogenic function, some HOX genes have been characterized as tumor suppressors in some types of cancers. HOXB13 inhibits prostate cancer growth by deactivating androgen receptor-mediated tumorigenic pathways. 29 HOXA5 was reported to transcriptionally induces p53 expression and activate p53-dependent apoptosis in breast cancer cells.30,31 Moreover, in colon cancer cells, HOXA5 drives the loss of stem cell phenotype and induces cancer stem cell to differentiate.32,33 These pieces of evidence not only indicate a complex function network associated with HOX gene family, but also highlight the necessity of a systematic exploration of HOX family in cancer development.
In summary, our study revealed that transcription factor HOXA10 is overexpressed in NPC and promotes the progression of NPC through transcriptionally activating the expression of SKP2, CAMKK2, and MMP1. Future studies are warranted to explore the clinical value of HOXA10 as a therapeutic target in NPC treatment and obtain a comprehensive view of HOX gene family in NPC development.
Footnotes
AUTHORS’ CONTRIBUTIONS
DG, HZ, LZ, RH, and JH conducted the experiment; DG and HZ analyzed the data; DG and JD conceived the original idea; DG wrote the initial version of the manuscript; JD supervised the project and revised the manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) received no financial support for the research, authorship, and/or publication of this article.