
Abstract
The myometrium is the smooth muscle layer of the uterus that generates the contractions that drive processes such as menstruation and childbirth. Aberrant contractions of the myometrium can result in preterm birth, insufficient progression of labor, or other difficulties that can lead to maternal or fetal complications or even death. To investigate the underlying mechanisms of these conditions, the most common model systems have conventionally been animal models and human tissue strips, which have limitations mostly related to relevance and scalability, respectively. Myometrial smooth muscle cells have also been isolated from patient biopsies and cultured in vitro as a more controlled experimental system. However, in vitro approaches have focused primarily on measuring the effects of biochemical stimuli and neglected biomechanical stimuli, despite the extensive evidence indicating that remodeling of tissue rigidity or excessive strain is associated with uterine disorders. In this review, we first describe the existing approaches for modeling human myometrium with animal models and human tissue strips and compare their advantages and disadvantages. Next, we introduce existing in vitro techniques and assays for assessing contractility and summarize their applications in elucidating the role of biochemical or biomechanical stimuli on human myometrium. Finally, we conclude by proposing the translation of “organ on chip” approaches to myometrial smooth muscle cells as new paradigms for establishing their fundamental mechanobiology and to serve as next-generation platforms for drug development.
Impact statement
The uterus is essential for reproduction and is susceptible to many untreatable disorders that cause adverse outcomes for women and the children they carry. Uterine contractility is driven by the smooth muscle cells found in the myometrium, the middle layer of the uterus, which have known sensitivity to a variety of hormones during different stages of menstruation, pregnancy, and labor. However, pregnancy and many uterine disorders, such as fibroids, also change the biomechanical load on myometrial smooth muscle cells, which likely impacts uterine contractility by mechanisms that are poorly understood, in part due to a lack of effective model systems. In this review, we describe existing model systems for investigating the mechanobiology of the myometrium, with a focus on in vitro techniques. We also argue that “myometrium on a chip” systems would be especially powerful for establishing fundamental aspects of myometrial mechanobiology and serve as new platforms for drug discovery.
Introduction
Complications with pregnancy and childbirth are relatively common worldwide and can lead to devastating outcomes for the mother and/or fetus. In the United States, approximately 1 in 10 infants are born preterm, defined as delivery prior to 37 weeks gestation. 1 Preterm birth is the leading cause of neonatal death 2 and preterm children are at increased risk for many developmental disorders. 3 Preterm birth also has adverse economic effects due to the increased need for hospitalization and care. 4 The most common cause of preterm birth is preterm labor, when the uterus begins contracting before the fetus reaches term. 5 Currently, medications known as tocolytics are used to treat preterm labor by reducing uterine contractions, many of which were developed for other conditions and are used off-label. 6 Examples include β-2 adrenergic receptor agonists and calcium channel blockers, such as nifedipine. 7 Consequently, these drugs also act on other types of muscle, including the heart, and cause adverse side effects. 8 Oxytocin receptor antagonists have also been developed, which are safer due to their specificity for uterine smooth muscle cells. 9 However, all tocolytics available today are relatively ineffective, as they only delay labor for approximately 48 h, during which time the mother can be transferred to an intensive care unit to prepare for inevitable preterm birth. 6
Another common complication of childbirth is insufficient progression of labor. This often leads to a Caesarean section (C-section), for which a surgical incision is made in the uterus to deliver the fetus. C-sections are the most common surgical procedure in the industrialized world. 10 In 2011, one in three births in the United States were by C-section. 11 One reason for insufficient progression of labor is weak contractility of the uterus.12,13 Low uterine contractility can also delay the initiation of labor, 14 another risky situation for the mother and fetus. 15 Insufficient uterine contractility can also cause postpartum hemorrhage, 16 the leading cause of maternal death. 17
C-sections also cause extensive scarring of the uterine wall, 11 which increases tissue rigidity 10 and can lead to many long-term adverse effects, such as reduced contractility, intermenstrual bleeding, and chronic pain, by mechanisms that are incompletely understood. Increases in tissue rigidity are also observed in other disorders that are independent of pregnancy, such as uterine fibroids, which have been reported in >70% of women. 18 Fibroids are non-cancerous, rigid masses in the myometrium composed of cells and matrix that can grow up to 20 cm. 19 Fibroids can be asymptomatic or cause heavy menstrual bleeding, pelvic pain, and infertility. Another disorder is adenomyosis, in which endometrial tissue grows in the myometrium. This can cause increased uterine contractility, 20 dysmenorrhea, and overexpression of oxytocin receptors, 21 leading to symptoms such as pelvic pain and infertility. 22 Similar to C-section scarring, fibroids and adenomyosis increase tissue stiffness.23,24
A major reason for the limited number of safe and effective therapies for most uterine disorders is that women’s health has been historically underrepresented in biomedical research. 25 Thus, there is an urgent clinical need to improve our understanding of the mechanobiology of the uterus, including the sensitivity of uterine contractions to a variety of biochemical and biomechanical stimuli. In this review, we first describe the structure and function of the myometrium, the smooth muscle layer of the uterus, and then survey conventional in vivo and ex vivo approaches for investigating properties of the myometrium. We then explain how two-dimensional (2-D) and three-dimensional (3-D) in vitro models of human myometrial cells can counter the weaknesses of conventional models because they lend themselves to rigorous and controlled molecular and functional analysis. Finally, we argue that applying “organ on a chip” approaches to model and interrogate human myometrium is a tremendous opportunity to expand our understanding of myometrial mechanobiology and serve as next-generation testbeds for developing safe and effective therapies for the many currently untreatable disorders of the uterus.
Structure and physiology of the myometrium
The structure of the uterus has evolved in support of its primary function: to support a fertilized egg as it develops into a fetus. In women aged 12–50 years on average, an egg is released from an ovary approximately every 28 days into a fallopian tube, where it may or may not be fertilized. The egg then travels to the uterus. If fertilized, it will implant into the innermost layer, an epithelial tissue known as the endometrium. Otherwise, the endometrium sloughs off at the end of each menstrual cycle, an event known as the menstrual period. 26 The middle layer of the uterus, the myometrium, consists primarily of smooth muscle cells (Figure 1). During the menstrual period, contraction of the myometrium helps expel the endometrium. 27 Contractions are also the source of dysmenorrhea, commonly known as menstrual cramps. In a pregnancy, the myometrium is highly stretched but remains quiescent during the 40-week gestation. Labor begins when the smooth muscle cells begin rhythmically contracting with increasing intensity to break the fetal membranes and help push the fetus through the birth canal. 26
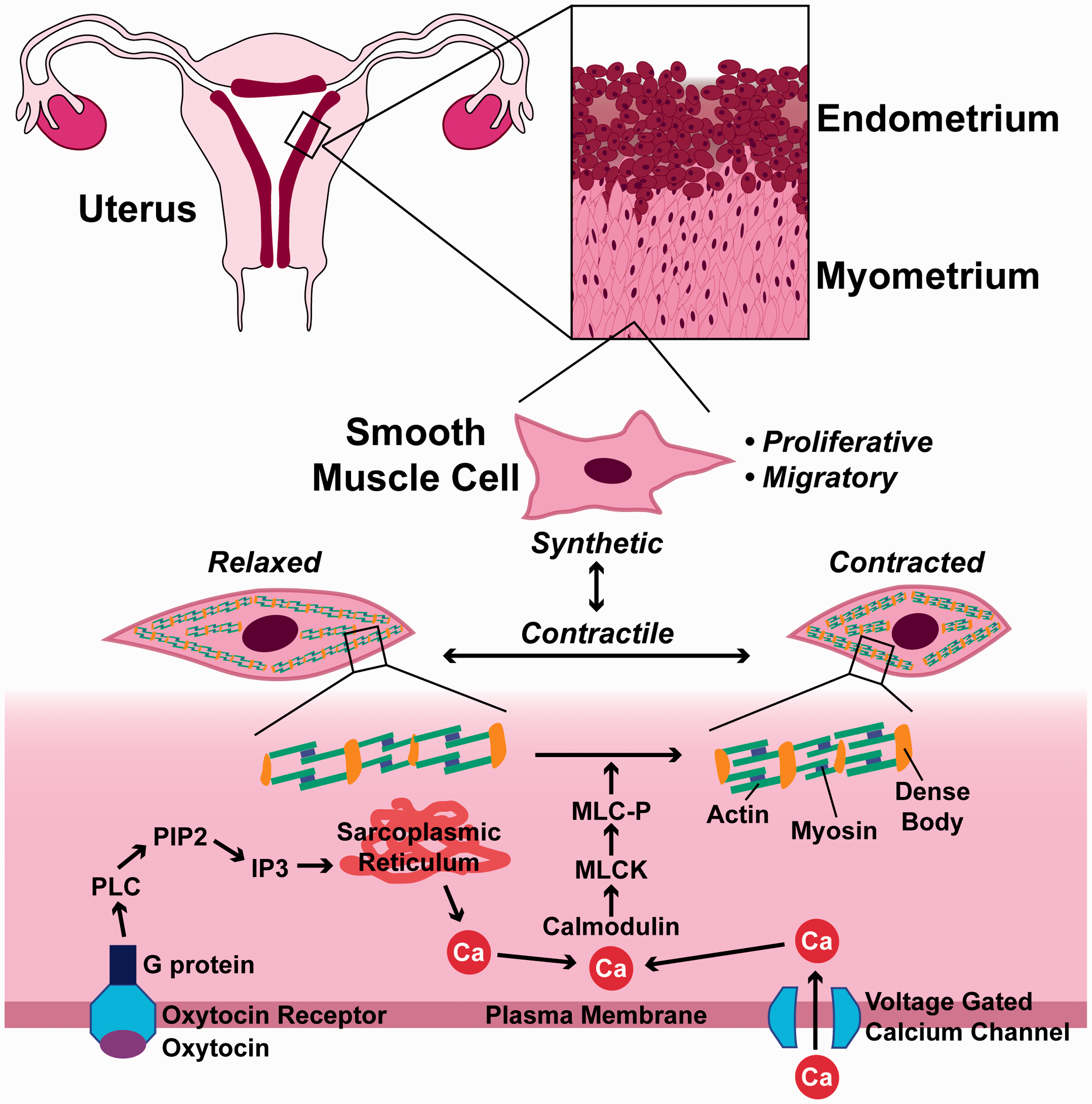
Multi-scale diagram of myometrial smooth muscle cells. Refer to the text for details. (A color version of this figure is available in the online journal.)
All smooth muscle cells are spindle-shaped and packed with elongated actin and myosin filaments that attach to the plasma membrane or to dense bodies in the cytoplasm, which anchor actin and myosin filaments as they slide past each other to generate force (Figure 1). Contraction is activated by calcium that enters the cytoplasm from extracellular fluid through voltage gated channels or that is released from the sarcoplasmic reticulum. 28 Calcium then binds to calmodulin, which activates myosin light chain kinase to phosphorylate myosin regulatory light chain and enable cross bridge formation between actin and myosin. 28 All smooth muscle cells have a basal tone, which is maintained by small, spontaneous calcium transients. In response to contractile agonists, larger, longer calcium transients cause stronger, sustained contraction. 29
Hormones are an essential regulator of contractility in myometrial smooth muscle cells. 30 In pregnancy, high levels of progesterone produced by the placenta 31 maintain uterine smooth muscle cells in a quiescent state by altering signaling pathways 32 and regulating expression of many genes, 33 including ion channels 34 and adrenergic receptors 35 that are directly involved in contractility. In most mammals, declines in progesterone are associated with the initiation of labor. In humans, however, progesterone levels do not decline. Instead, uterine smooth muscle cells experience a “functional withdrawal” of progesterone, guided mostly by changes in progesterone receptor expression and progesterone metabolism. 36 During labor, pulsatile secretions of oxytocin coupled with increased expression of oxytocin receptors by smooth muscle cells are acute activators of contraction. 30 Oxytocin receptors are G protein-coupled receptors that, upon oxytocin binding, activate phospholipase C (PLC) to convert phosphoinositide-bis-phosphate (PIP2) into inositol-tris-phosphate (IP3), which stimulates release of calcium from the sarcoplasmic reticulum 37 (Figure 1). Oxytocin may also increase expression of the gap junction protein connexin 43, which facilitates electrical communication between cells and enables synchronized contractions of the myometrium.37,38
Another important aspect of smooth muscle cell physiology and pathophysiology is their ability to switch phenotypes. Unlike striated muscle cells, which are terminally differentiated, smooth muscle cells transition between contractile and synthetic phenotypes (Figure 1). In the contractile phenotype, cells up-regulate contractile proteins, such as α-smooth muscle actin, and contract in response to relevant stimuli.29,39,40 In the synthetic phenotype, cells down-regulate contractile proteins and are migratory and proliferative.29,39,40 Pregnancy begins with a synthetic phase that is characterized by proliferation of cells, matrix synthesis, and anti-apoptosis. This phase is followed by an intermediate synthetic phase, and ends with a contractile phase. 41 Aberrant phenotype switching is thought to play a role in smooth muscle diseases, such as asthma 39 and vasospasm, 29 and thus may also play a role in uterine disorders.
In vivo and ex vivo models of the myometrium
Historically, animal models have been implemented to investigate healthy myometrial tissue and certain myometrial pathologies. 42 Compared to human tissue, animal tissue is more readily available and can be sampled at various time points, which is particularly important when studying the time-dependent progression of pregnancy to labor. Non-human primates are most similar to humans, as they have similar litter sizes, the placenta is the main source of progesterone, and systemic progesterone withdrawal is not required for labor. 43 Sheep are also used as a model for pregnancy due to their similarities to humans in terms of fetal weight, 43 progesterone source,43,44 and litter size. 44 However, unlike humans, sheep experience reduced levels of progesterone prior to labor. 43 Due to their high cost and ethical concerns, these large animal models are reserved primarily for later stage studies. More commonly used in vivo animal models for pregnancy include rabbits, 45 rats, 46 mice, 47 and guinea pigs. 48 These smaller animal models are less expensive but also generally less relevant to humans. For example, progesterone in rabbits is produced by the corpus luteum for the duration of the pregnancy instead of the placenta. 44 Rats and mice exhibit contractions with higher frequency and shorter duration compared to humans. 37 Rabbits, rats, and mice also experience progesterone withdrawal during later stages of pregnancy and have large litter sizes. 44 Compared to other rodents, guinea pigs are considered the most appropriate animal model because progesterone is produced by the placenta and maintained in late gestation. Guinea pigs also commonly exhibit preterm birth similar to humans and have smaller litters compared to other rodents. 44 However, no animal model completely replicates human uterine physiology.
Human ex vivo samples of the myometrium are a less accessible but generally more clinically-relevant alternative to animal models. Tissue biopsies (1–2 cm3) are typically procured from patients during elective C-sections prior to the onset of labor and then dissected to remove non-myometrial tissue 37,49,50 (Figure 2(a)). The remaining tissue is stored at 4 °C and used within one 51 or two 52 days of collection. Samples are then further sliced into tissue strips on the order of millimeters,37,53 often using an oscillating vibroslicer. 49 The length of the strip ultimately depends on the type of analysis that will be performed, whether functional or molecular.
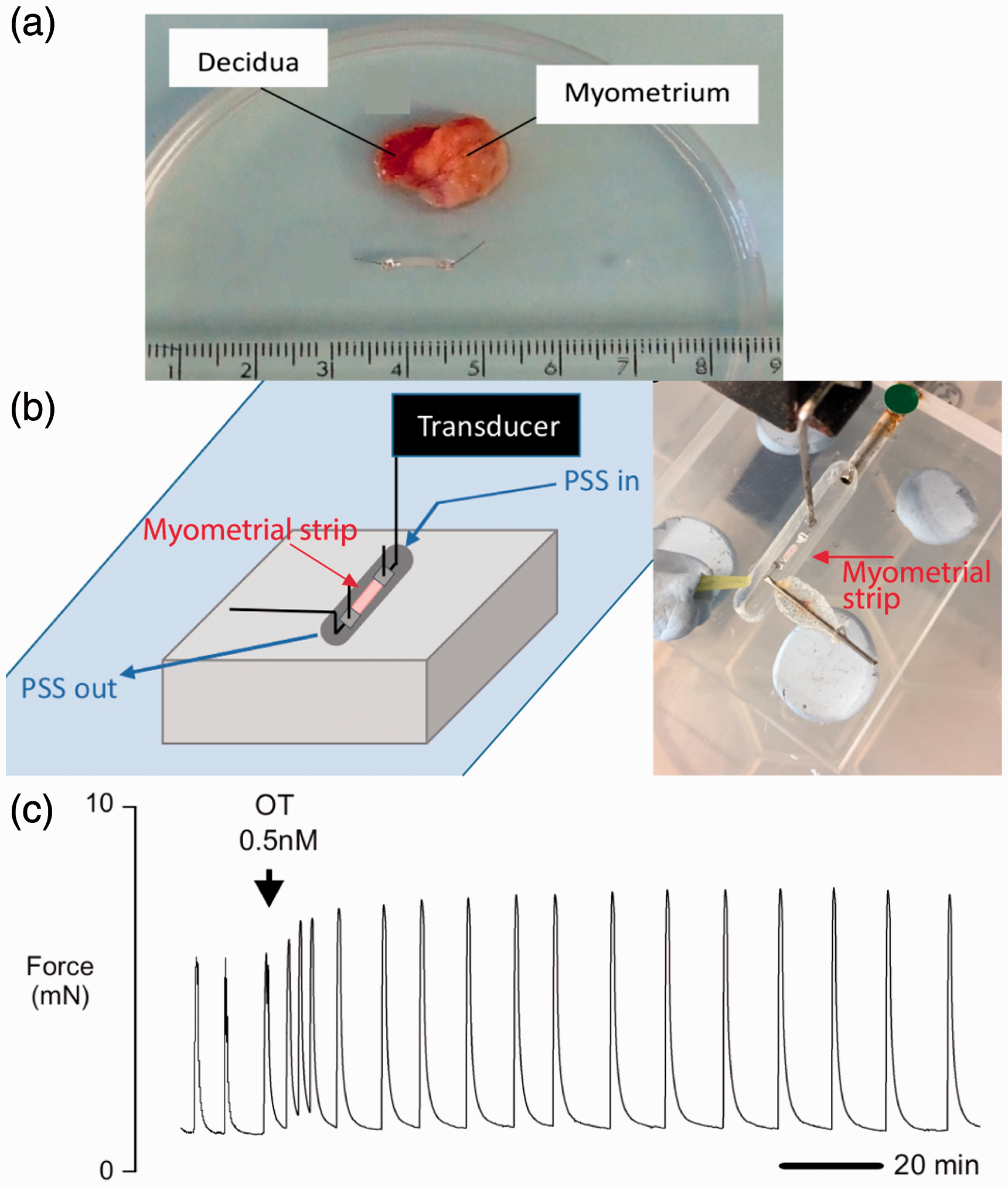
Measuring the contractility of human myometrial tissue strips. (a) Top: Full thickness biopsy of human myometrial tissue and decidua. Bottom: dissected myometrial tissue strip (2 mm wide, 8 mm long, 1 mm thick) attached to aluminum tissue clips. (b) Diagram and photograph of a strip of myometrium mounted horizontally in a heated organ bath chamber. One end is affixed to a hook and the other end is connected to a force transducer. The strip is perfused with physiological saline solution (PSS) using a peristaltic pump. (c) Myometrial tissue strips exhibit spontaneous contractions, which increase in amplitude after the addition of oxytocin (OT). Images reproduced from Arrowsmith et al. 51 (https://creativecommons.org/licenses/by/3.0/us/legalcode). (A color version of this figure is available in the online journal.)
Myometrial tissue strips that do not demonstrate spontaneous contractile activity within about two hours are typically excluded from further functional analysis.49,51 From the remaining tissue strips, contractile forces can be measured by attaching one end of the strip to a force transducer and affixing the other end to an organ bath chamber (Figure 2(b)). After anchoring and applying basal tension, contractile responses to agonists and antagonists can be measured (Figure 2(c)). One study leveraged this method to compare tocolytic responses within the same patient and showed that nifedipine and indomethacin caused concentration-dependent reductions in contractility. 50 When the tocolytics were paired with progesterone, contractions were further inhibited. Another study used tissue strips to expand the basic understanding of drug responses, including targets of interests and pathways of contraction, for the purposes of drug discovery. 51 This study used oxytocin and vasopressin in combination with their receptor antagonists to validate their platform.
Because contractions in smooth muscle cells are initiated by a rise in intracellular calcium, fluorescent calcium indicators are often used as a proxy for contractility that are generally more accessible to researchers compared to the force transducer system. 49 For these assays, tissue strips are loaded with a calcium sensitive dye, such as Fluo-4, and changes in fluorescence intensity are recorded after normalization to a resting state. This method has been leveraged to measure calcium propagation in tissue strips, demonstrating that intracellular calcium signals spread faster along bundles of myocytes than between bundles. 49 Thus, ex vivo tissue strips have proven to be a useful model system for evaluating the contractility of human myometrium. However, tissue scarcity and ethical considerations increase the time and financial cost of acquiring samples. Furthermore, patient-specific differences and the multitude of confounding factors present in ex vivo tissue limit the reproducibility of tissue strips, which hinders their usefulness for drug screening or in-depth mechanistic studies.
Cell sources for in vitro models of the myometrium
In vitro models can overcome several of the limitations described above for in vivo and ex vivo models. In general, primary cells are the closest representation of cells in situ and thus are an attractive option for in vitro studies. To obtain primary smooth muscle cells from human myometrium, tissue samples are usually obtained during an elective C-section, following appropriate ethical procedures. 54 Cells are then enzymatically isolated using a single type of collagenase,55,56 a combination of multiple collagenases,54,57 or a mixture of collagenase and trypsin. Once isolated, cells are usually cultured in Dulbecco’s Modified Eagle’s medium (DMEM) with serum, 57 L-glutamine, and antibiotics55,57 and plated on collagen coated substrates.55,58 Cells can be passaged using standard protocols and are typically used at low passages (between 3 and 5), 59 but can be used for up to 10 passages. 57 However, one study showed that primary myometrial cells transition from a contractile to a synthetic phenotype in vitro, 56 which is problematic because a contractile phenotype must be maintained for these cells to be a relevant model for labor. Another study addressed this concern by measuring RNA levels of several contractile markers in primary myometrial cells, including α-smooth muscle actin, connexin 43, and oxytocin receptors. The expression of these markers was stable for at least 10 passages, with 10–14 days between passages, 57 countering previous data suggesting that these cells cannot maintain a contractile phenotype in vitro. 56 However, although the expression of contractile markers has been measured, there is a great need to rigorously evaluate the functional contractility of primary myometrial smooth muscle cells in vitro, which could be achieved by utilizing some of the assays described in the next section. Furthermore, many strategies developed for promoting a contractile phenotype in vascular smooth muscle cells in vitro 60 can likely be adopted for myometrial smooth muscle cells as well, but have not yet been explored.
Diseased myometrial tissue has also been evaluated in vitro with primary cells. In one study, healthy myometrial tissue and tissue afflicted with leiomyomas, which are benign tumors or fibroids, were obtained from premenopausal women undergoing hysterectomy. 55 Both tissue types were dissected into small pieces, incubated in culture media with DMEM with serum, collagenase, and antibiotics, and cultured by the explant method for three weeks. RNA expression analysis demonstrated that fibrotic tissue exhibited higher levels of α-smooth muscle actin. 55 By using primary cells from diseased and healthy tissues derived from the same patient, this study was able to validate a method to investigate the mechanisms that develop leiomyomas while accounting for patient-specific differences.
Although primary cells obtained from freshly harvested tissue samples closely retain the morphology, signaling pathways, and receptors as cells in situ, primary cells have poor longevity and proliferate slowly in culture. After a few passages, the cells generally cease to divide at a rate to produce an adequate quantity of cells for analysis. 61 Isolating cells from multiple patients helps increase cell quantity, but at the expense of introducing patient-specific variability. To address these challenges, primary cells have been immortalized to generate cell lines that better maintain consistency throughout passages.61–63 One mechanism of cell immortalization involves retroviral transfection with hTERT, a reverse transcriptase that prevents telomere shortening. This approach has been used to immortalize myometrial cells from the lower uterine segment. A comparison between immortalized cells and primary cells demonstrated minimal differences on a molecular level. 61 For example, α-smooth muscle actin and oxytocin receptor levels were maintained, as shown by immunofluorescent imaging and gene expression analysis. Immunoblotting analysis of signaling pathways, such as stimulation of growth, contractile activity, and proinflammatory responses, found few differences between hTERT immortalized cells and primary cells. 61
Another immortalized uterine myometrial cell line, PHM1-41, was derived from a single pregnant patient at 39 weeks of gestation who was not in labor. 62 Briefly, muscle tissue was isolated from endometrial and stromal tissue, enzymatically digested, then immortalized via infection with a human papilloma virus vector. 63 The immortalized cells were compared to unmodified fresh cells, showing minimal differences in α-smooth muscle actin levels and the release of intracellular free calcium in response to oxytocin. The PHM1-41 cell line is commercially available and can be maintained in culture for up to 22 passages, as compared to primary cells that senesce between 12 and 13 passages. 62 Thus, immortalized human cell lines are a viable alternative to human primary cells for investigating certain aspects of myometrial biology or evaluating drug responses due to their higher growth rates and decreased senescence. One major caveat of cell lines is that they likely lose their contractile characteristics more so than primary cells. However, the functional phenotype of immortalized cells lines has not been thoroughly evaluated or compared to primary myometrial cells. In summary, although several sources of myometrial smooth muscle cells currently exist, there is a major need to determine the extent to which both primary cells and immortalized cell lines maintain functional contractile phenotypes in vitro before they can be implemented as reliable models for elucidating mechanisms of uterine mechanobiology and identifying triggers of labor.
Measuring contractility in response to biochemical stimuli in vitro
Assays for measuring contractile forces in vitro have traditionally paralleled ex vivo approaches. For example, the aforementioned method for using fluorescent calcium indicators to measure intracellular calcium dynamics has also been implemented for in vitro models. In one example, mouse myometrial cells were cultured in 384-well plates and loaded with Fluo-4 (Figure 3(a)). Fluorescence intensity was then measured in response to oxytocin and a variety of compounds to identify contractile agonists and antagonists, which were then validated with an ex vivo contractility assay. 64 This approach has much potential as a large-scale, high throughput screening system for identifying novel small molecules that impact uterine contractility.
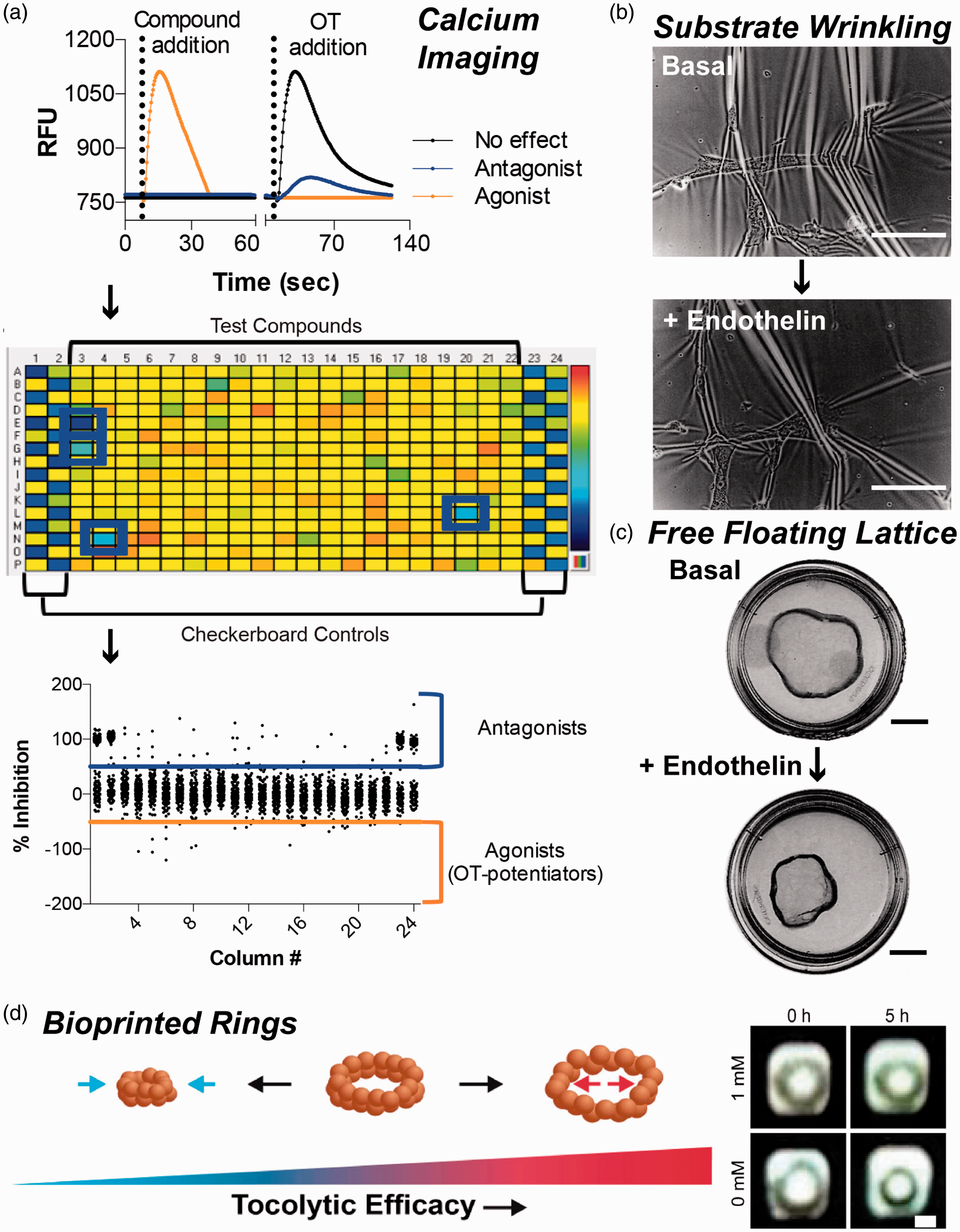
Assays for measuring the contractile response of myometrial smooth muscle cells to biochemical stimuli. (a) Myometrial cells were loaded with the calcium indicator dye Fluo-4 and treated with compounds and/or oxytocin (OT). Agonists and OT induced calcium transients. Antagonists reduced OT-induced calcium transients (top). This assay was implemented for cells in 384-well plates (middle) as a high-throughput screen for identifying agonists and antagonists (bottom). Images reproduced from Herington et al. 64 (https://creativecommons.org/licenses/by/4.0/legalcode). Images of the (b) substrate wrinkling assay (scale bar, 4 mm) and (c) free-floating lattice assay (scale bar, 7 mm) for assessing myometrial smooth muscle cell contractility in response to endothelin. Images reproduced from Dallot et al. 59 by permission of Oxford University Press. (d) Diagram of magnetically bioprinted 3-D myometrium rings (left), which increase in diameter in response to tocolytics. Rings treated with the contractile antagonist indomethacin (1 mM) at t = 0 h are relaxed by t = 5 h (scale bar = 1 mm). Images reproduced from Souza et al. 67 (A color version of this figure is available in the online journal.)
More directly assess the contractility of uterine smooth muscle cells in vitro One approach to is substrate wrinkling. 59 Cells are incubated on a compliant surface, such as silicone, and changes in the wrinkling pattern are used to indicate changes in cell tension (Figure 3(b)). However, this method does not lend itself to identification of agonists or antagonists because the wrinkling response is not standardized. For example, although increased wrinkling is the expected response to a contractile agonist, decreased wrinkling has also been shown. Thus, this method offers mostly qualitative results and is not suitable for rigorous comparisons between samples or chemical stimulants.
3-D in vitro models of myometrial tissue are an attractive option because they offer the simplicity and control of in vitro methods while more closely recapitulating the spatial architecture of in vivo models. 65 One approach is the free-floating lattice method (Figure 3(c)), which also offers a more quantitative method for measuring contractility compared to substrate wrinkling. Cells are cultured within a deformable material, such as a collagen gel, which is then separated from the culture well to form a free-floating lattice. A reduction in area of the floating gel indicates an increase in basal contractile tone, which can be measured over time. This method has been used to demonstrate an increase in contractility due to endothelin-1, 59 TNF-α, 58 and thrombin 58 and a relaxing effect of indomethacin. 58 This technique was also used to identify endothelin-A receptor (ETA) as a mediator of contractility after the addition of an ETA-selective agonist prevented a contractile response. 59 Thus, the free-floating lattice method can be implemented to identify small molecules that induce the contraction and relaxation of 3-D myometrial smooth muscle tissues in vitro.
A 3-D model of the myometrium has also been developed for modeling leiomyomas, which are associated with excessive fibrosis. First, immortalized patient-derived myometrial and leiomyoma cells were embedded in 3-D collagen gels, where they became characteristically spindle-shaped and expressed α-smooth muscle actin. Next, biomarkers were characterized and compared in 2-D cultures, 3-D cultures, and freshly isolated tissue strips. Fibronectin and fibromodulin, both components of the extracellular matrix, were up-regulated in leiomyoma cells in 2-D cultures but not 3-D cultures or freshly isolated tissues strips, highlighting the similarities between 3-D cultures and native tissues compared to 2-D cultures. 2-D and 3-D cultures were also treated with integrin-β1 function-inhibiting antibody, which caused cell detachment in 3-D cultures, as expected, but not 2-D cultures. 66 This study provides evidence that 3-D in vitro models have more physiologically relevant phenotypes related to the extracellular matrix and integrin activity compared to 2-D models, which is especially important for modeling fibroids and other conditions associated with excessive and unorganized extracellular matrix formation.
3-D rings of myometrial tissue have also been magnetically bioprinted for contractility measurements. 67 In this approach, myometrial cells are first magnetized using nanoparticles and then cultured under magnetic levitation to allow cells to produce endogenous extracellular matrix. Cells are then resuspended and transferred to a well-plate atop ring-shaped magnets, which attract cells into a ring-shaped pattern. When the magnet is released, cells contract and the change in area is measured as an indicator of basal contractile activity (Figure 3(d)). Similar to the free-floating lattice method, time course studies were performed to capture the change in ring area in response to tocolytics. 67
Measuring contractility in response to biomechanical stimuli in vitro
In addition to biochemical stimuli, smooth muscle cell function and phenotype is also regulated by diverse forms of mechanical stimuli, such as stretch. Stretch increases cell length and the amount of overlap between actin and myosin filaments. For this reason, the amount of induced stress that a muscle cell can generate is optimal at an intermediate level of stretch, a phenomenon known as the length-tension relationship.68,69 Beyond their immediate response to stretch, smooth muscle cells can also adapt to sustained stretch by remodeling their cytoskeleton.70,71 The adaptation of vascular smooth muscle cells has been characterized in vitro by culturing cells on stretchable membranes and measuring force generation at different timepoints after sustained stretch. After 1 h of stretch, vascular smooth muscle cells have higher basal tone, which approaches un-stretched levels of basal tone after 24 h. 72 Similar results have been observed in airway smooth muscle cells. 73 Like phenotype switching, aberrant mechanoadaptation is thought to contribute to diseases such as asthma 74 and vasospasm. 72 Mechanical stretch has also been shown to immediately increase calcium 75 and alter expression of genes associated with synthetic and contractile phenotypes29,76 in non-uterine smooth muscle cells. Thus, stretch can have many short- and long-term impacts on the contractility of smooth muscle cells via multiple mechanisms, which together have a major role in the physiology and pathophysiology of smooth muscle organs.
The response of uterine smooth muscle cells to stretch is especially relevant because the uterus is highly distended at later stages of pregnancy. Several pieces of evidence suggest that stretch increases uterine contractility and is a trigger for labor. For example, contractility is higher in strips of uterine smooth muscle isolated from women who give birth to twins, who would have a more distended uterus, compared to singletons. 77 This study also found a correlation between contractility and neonatal birth weight, which can be considered a proxy for uterine stretch. 77 Carrying multiple gestations is also a known risk factor for preterm labor, suggesting that high levels of stretch could trigger preterm labor. 78
Compared to other types of smooth muscle, relatively little is known about how myometrial smooth muscle cells respond to stretch. Stretch has been shown to immediately stimulate contraction and increase intracellular calcium levels in rat and human myometrial tissue strips79–82 and human myometrial smooth muscle cells. 83 However, these studies are mostly limited to measuring acute effects of stretch and do not evaluate mechanoadaptation. Most studies to date have also relied on tissue strips, which have confounding factors that cannot be separated from direct effects on the smooth muscle cells. There is also evidence of altered stretch sensitivity as pregnancy progresses, as myometrial tissue strips isolated from pregnant mice have a shifted length-tension relationship as gestation advances. 84 However, mechanisms guiding these changes are unknown.
In addition to stretch, matrix stiffness has also been shown to regulate smooth muscle cell phenotype and contractility. For example, evidence suggests that vascular85–87 and airway 88 smooth muscle cells transition from a contractile to a synthetic phenotype on stiffer surfaces, which could contribute to smooth muscle dysfunction in diseases like atherosclerosis. However, the response of uterine smooth muscle cells to matrix stiffness has not been thoroughly characterized. One study reported an increase in uterine smooth muscle cell size with increasing elastic modulus, 89 consistent with a synthetic phenotype, but the effects of matrix stiffness on cell function are unknown.
Future opportunities for modeling human myometrium
As described in the previous section, many fundamental aspects of myometrial mechanobiology are still unknown. One major unanswered question is: What are the mechanosensors in myometrial smooth muscle cells? In vascular smooth muscle cells, stretch has been shown to activate platelet-derived growth factor receptors, focal adhesion kinase (FAK), and stretch-activated channels. 90 FAK and stretch-activated channels have also been implicated as sensors of matrix stiffness in vascular smooth muscle cells. 91 Myometrial smooth muscle cells are known to express mechanosensitive ion channels, such as transient receptor potential canonical channels,83,92–94 which allow calcium entry, and TWIK-related potassium channels, which suppress contractility in response to stretch.95–97 Thus, myometrial smooth muscle cells express multiple types of mechanosensitive ion channels that can stimulate or suppress contractility. Their balance is likely important for maintaining quiescence during gestation and activating labor at appropriate times. However, the identities of active mechanosensors and mechanisms of mechanosensing in myometrial smooth muscle cells are still unestablished, which is critical information needed to understand labor and uterine disorders and ultimately identify therapeutic targets for intervention.
To answer these and other questions related to myometrial contractility, new approaches for modeling human myometrium in vitro are critically needed. Inspiration can be taken from the several sophisticated approaches for engineering and interrogating striated and smooth muscle tissues “on a chip” that have been developed using a variety of microfabrication techniques. For example, 2-D muscle tissues have been engineered by culturing cells on surfaces microcontact printed with extracellular matrix proteins98,99 or embedded with topographical features.100,101 The contractility of these tissues has been directly measured using traction force microscopy98,102,103 or the cantilever-based muscular thin film assay.99,104 3-D muscle tissues have been generated by casting cell-hydrogel mixtures into custom molds105–107 or by leveraging new advances in bioprinting.108–110 The contractility of these 3-D tissues has been measured by attaching tissues to a force transducer 105 or by monitoring the deflection of tissue anchor points.111,112 Although these approaches for patterning tissues and quantifying contractility have yet to be translated to myometrial smooth muscle cells, they are likely compatible and could be implemented as new paradigms for engineering in vitro models of human myometrium that recapitulate key features of native tissue structure, such as alignment and matrix rigidity, while also facilitating rigorous functional measurements at scale. These technologies would be powerful for investigating basic mechanobiology and screening the functional effects of drugs, such as tocolytics, 9 and other types of biochemical stimuli, such as adipokines, which are elevated in obesity and thought to affect uterine contractility. 113 The effects of environmental stimuli, such as cigarette smoke or nanoparticles, could also be tested. Furthermore, a “myometrium on a chip” model could be coupled to existing “organ on chip” models of the placenta, 114 endometrium,115,116 and cervix. 115 Together, these would be powerful systems for understanding the complex interconnectivity between different cell and tissue types in a more complete model of the female reproductive system.
Another important future direction is the development of patient-specific models of human myometrium, which is especially important for understanding and addressing certain disparities in uterine disorders. For example, non-Hispanic Black women are at twice the risk for preterm birth compared to non-Hispanic white women for unknown reasons. 117 Investigating this and other patient-specific differences from a molecular perspective requires the use of patient-derived cells. However, as described above, cell sources for in vitro models of human myometrium are limited to primary cells, which have limited scalability and reproducibility, and cell lines, which have limited physiological relevance and cannot be implemented to correlate patient genotype to phenotype or identify patient-specific risk factors. Cells derived from human induced pluripotent stem cells (hiPSCs) mitigate these limitations because they can be easily expanded in culture while also preserving patient-specific genotypes. However, although methods for differentiating hiPSCs into vascular smooth muscle cells 118 have been established, there are currently no techniques for differentiating hiPSCs into myometrial smooth muscle cells. This is an important topic for future work, as these cells would enable comparison of myometrial function between patients with disease-relevant genetic mutations or from different racial or ethnic backgrounds.
In summary, although uterine function is highly dependent on the contractility of myometrial smooth muscle cells, we still have a relatively poor understanding of the response of these cells to a variety of biochemical and biomechanical stimuli, especially compared to other types of muscle that have historically been more represented in biomedical research. These issues motivate the development of more sophisticated in vitro models of human myometrium, especially those that integrate patient-derived cells into microfabricated “organ on chip” devices that enable recapitulation of tissue architecture and rigorous functional measurements at scale. These types of approaches can then be implemented to establish fundamental aspects of uterine mechanobiology and serve as platforms for developing and screening safe and effective therapies for a variety of uterine disorders, leading to better health outcomes for mothers and newborns.
Footnotes
AUTHORS’ CONTRIBUTIONS
All authors participated in the development and writing of this manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by the University of Southern California Annenberg and Graduate School Fellowships (APM) and NSF CAREER Award CMMI 1944734 (MLM).