
Abstract
Schisandra chinensis and Evodia rutaecarpa are traditional Chinese herbs that have been used for many years to treat neurodegenerative diseases. In Chinese medicine, multiple herbs are often used in combination to enhance their efficacy, and different combination ratios can produce different therapeutic effects, thus flexibly responding to the needs of various patients. This study aimed to investigate the effects of different ratios of Schisandra and Evodia herbs on learning and memory impairment in rats with Alzheimer’s disease (AD) and their specific mechanisms of action. Morris water maze and hematoxylin and eosin (HE) staining experiments were performed to evaluate the effects of different ratios of Schisandra-Evodia on learning memory in AD model rats. Immunohistochemical experiments were performed to investigate the effects of Schisandra-Evodia on the Aβ1-42 and P-Tau proteins, and protein immunoblotting (WB) was performed to determine the expression of key proteins in two pathways, BDNF/TrkB/CREB and GSK-3β/Tau. Our experimental results show that all Schisandra-Evodia groups showed significant neuroprotective effects, improved learning memory impairment, and reduced levels of Aβ1-42 and P-Tau proteins in AD model rats. Schisandra-Evodia upregulated BDNF, P-TrkB/TrkB, and P-CREB/CREB protein expression and downregulated GSK-3β and P-Tau/Tau protein expression. Among the different Schisandra-Evodia ratio groups, the 2:1 group showed the strongest therapeutic effect on AD. Our research results indicate that Schisandra-Evodia can reduce Aβ1-42 and P-Tau protein content by modulating the activity of two pathways, BDNF/TrkB/CREB and GSK-3β/Tau, thus improving neuronal cell damage and cognitive deficits caused by AD. In addition, we found that a Schisandra-Evodia ratio of 2:1 had the most profound therapeutic effect on AD.
Impact statement
This study fills a gap in the research on the use of combined Schisandra and Evodia for the treatment of Alzheimer’s disease (AD). Our study is the first to report that different ratios of Schisandra-Evodia can modulate both the BDNF/TRKB/CREB and GSK-3β/Tau pathway activities, thereby inhibiting the production and aggregation of Aβ1-42 and P-Tau proteins in the hippocampal region of rats. Notably, this effect seemed to vary with the proportion of the two herbs. Furthermore, through a comprehensive analysis of different Schisandra-Evodia ratio groups for treating AD in model rats, we concluded that a Schisandra-Evodia ratio of 2:1 had the best therapeutic effect on AD. Our study provides a novel approach for an alternative treatment for AD and a reference for clinical drug use. Through this study, we expect to provide additional treatment options for patients with AD, thus improving their quality of life.
Introduction
Alzheimer’s disease (AD) is the most common cause of dementia; 1 the two most prominent pathological features of AD are senile plaques caused by β-amyloid (Aβ) deposition and neurofibrillary tangles due to Tau protein hyperphosphorylation. 2 The behavioral symptoms of AD are closely associated with the accumulation of Aβ and P-Tau proteins, which are a direct result of impaired and disrupted synapses that mediate memory and cognition. As Aβ and P-Tau protein pathology progresses, it gradually causes substantial structural and functional violations of the patient’s brain, mainly manifesting as cognitive deficits and memory loss, 3 which seriously affect the daily life and life health of patients with AD. Therefore, reducing Aβ and P-tau protein levels in the brain of patients is an important research direction for AD treatment. However, the process of research and the failure of clinical trials for single-target Aβ and Tau proteins suggest that when the pathological process of hyperphosphorylated Tau is initiated, Aβ and Tau pathological processes will act in parallel pathways to mutually enhance each other’s toxicity to aggravate AD pathological damage.4,5 This calls for a shift from a single-target treatment for AD to a multifaceted, multi-pathway approach to simultaneously arrest the progression of different pathological factors in AD. 6
BDNF, a neurotrophic factor that promotes neuronal differentiation and regeneration, is involved in neuroplastic mechanisms and is associated with AD. Studies have shown that BDNF protects neurons from the toxicity of the Aβ protein, 7 reverses neuronal atrophy, and improves cognitive deficits. 8 The prerequisite for BDNF to function is its binding to the high-affinity promyosin-related kinase B (TrkB) receptor, which in turn activates the phosphorylation of the cAMP response element-binding protein (CREB), promoting neuronal growth, differentiation, and survival, and enhancing synaptic plasticity. GSK-3β is a protein serine/threonine kinase that plays an essential role in the pathological process of AD. It is a key kinase that promotes Tau protein hyperphosphorylation, 9 which is a fundamental factor that leads to abnormal neuronal function and, ultimately, neuronal death. Therefore, if both BDNF/TrkB/CREB and GSK-3β/Tau pathway activities can be regulated simultaneously, it may be more beneficial to stop the pathological process of Aβ and P-Tau proteins and thus improve cognitive deficits and learning and memory impairment caused by AD.
The Chinese herb Schisandra chinensis, the dried ripe fruit of Schisandra chinensis (Turcz.) Baill. of the Magnolia family, is commonly used in current clinical practice to improve cognitive function; 10 Schisandra has been reported to resist oxidative stress, have anti-inflammatory properties, inhibit apoptosis, and treat neurodegenerative diseases.11,12 Recent studies have shown that Schisandra can produce antidepressant-like effects in mice induced by repetitive corticosterone through the BDNF/TrkB/CREB signaling pathway. 11 In a rat model of scopolamine-induced mild cognitive impairment, Schisandra upregulated the expression of BDNF and its downstream signaling molecules, p-ERK and p-AKT, and enhanced the memory capacity of model rats. 13 However, Schisandra has only been reported to inhibit the production of the Aβ protein, 14 and there are only a few reports on the involvement of Schisandra in Tau protein hyperphosphorylation. In Chinese medicine, holistic therapy of multiple herbal combinations is often used for better therapeutic efficacy, and the medicine pair is the most basic and simple form of the compound formula, which is a centralized representative of herbal combinations. Given their simplicity and the basic characteristics of complex formulas, herbal pairs are greatly significant in the study of herbal combinations. 15 According to the traditional Chinese medicine theory, Schisandra and Evodia synergize with each other to achieve the effects of warming and nourishing Qi, calming the mind, opening the orifices, and waking the brain, which are useful in the treatment of neurodegenerative diseases. The Chinese herb Evodia rutaecarpa is the dried, near-ripe fruit of Evodia rutaecarpa (Juss.) Benth. of the Rutaceae family; modern medical studies have shown that the crude extract of Evodia and its active constituents have a wide range of pharmacological activities, including having antitumor, 16 anti-inflammatory, 17 and antioxidant properties, acting as a modulator of central nervous system homeostasis, 18 and cardiovascular protection. 19 A recent study has shown that Evodia can exert neuroprotective effects by inhibiting Tau phosphorylation through the inhibition of the GSK-3β/CDK5/MAPK pathway. Therefore, we speculated that the combination of Schisandra and Evodia could act on both the BDNF/TrkB/CREB and GSK-3β/Tau pathways to provide a “multi-target, multi-pathway” treatment for AD. In addition, it should be noted that the combination of the two herbal medicines should be guided by medical books and that their therapeutic effects often vary with different ratios of the two herbal medicines. 15 Therefore, the goal of this study was to determine the optimal Schisandra-Evodia ratio for the treatment of AD.
This study aimed to investigate the therapeutic effects of different ratios of Schisandra-Evodia on AD and its molecular mechanism of action and to determine the optimal Schisandra-Evodia ratio for AD treatment. Our study provides a novel approach for an alternative treatment for AD and a reference for clinical drug use. Through this study, we expect to provide additional therapeutic options for patients with AD and improve their quality of life.
Materials and methods
Animals and drugs
Eight- to nine-week Sprague-Dawley (SD) male rats, weighing 200 ± 20 g, were purchased from the Experimental Animal Center of our university (animal license number: SCXK (black) 2019-001). Rats were housed in groups of five per cage, allowed casual access to water and food, and maintained at a constant temperature (23 ± 1°C) and humidity (55 ± 5%) under a 12-h light/dark cycle (lights on from 07:00 to 19:00 h). Animal experiments were performed following the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and the experimental protocol was approved by the Animal Ethics Committee of our university (approval number: 20201029001). Guidelines for the care and use of experimental animals were strictly adhered to when conducting the animal studies. The use of animals and their suffering were minimized.
The Aβ1–42 (St. Louis, MO, USA) peptide was dissolved in saline and diluted to a reserve concentration of 1.0 mg/mL. The Aβ1–42 solution was incubated at 37°C for five days to obtain the fibrillated form of Aβ1–42. Schisandra chinensis and Evodia rutaecarpa were purchased from Shenyang Tongrentang Pharmaceutical Co, Ltd. (Shenyang, China), and groups with different ratios of Schisandra-Evodia (4:1, 2:1, 1:1, 1:2, 1:4) were accurately weighed, totaling 150 g each and ground into powder, heated, and condensed to reflux twice for two hours each. The filtrates were then combined and concentrated using a rotary evaporator. The concentrated extracts were set aside.
Treatment and experimental design
Seventy rats were randomly divided into seven groups, including the control, model (Aβ1-42), Schisandra-Evodia4:1 treatment (Aβ1-42 + 2 g/kg Schisandra-Evodia), Schisandra-Evodia2:1 treatment group (Aβ1-42 + 2 g/kg Schisandra-Evodia), Schisandra-Evodia1:1 treatment group (Aβ1-42 + 2 g/kg Schisandra-Evodia), Schisandra-Evodia1:2 treatment group (Aβ1-42 + 2 g/kg Schisandra-Evodia), and Schisandra-Evodia1:4 (Aβ1-42 + 2 g/kg Schisandra-Evodia) treatment groups. All rats were anesthetized with sodium pentobarbital (30 mg/kg), and a guide cannula (25-gauge) was implanted in the right lateral ventricle. The stereotaxic coordinates were 4 mm from the dural surface, 1 mm posterior to the bregma, and 1.5 mm right lateral to the midline. About 1 μl of Aβ1-42 was injected into the lateral ventricle of the rats in the model and treatment groups. An equal volume of physiological saline was administered to the rats in the control group. The needle attached to the Hamilton syringe was left for five minutes after injection of Aβ1-42. The guide cannula was covered with a medical gelatin sponge to prevent the cerebrospinal fluid and injection to drain out. After one week of adaptation, rats in the administration group were administered the drug by daily gavage, and rats in the control and model groups were administered an equal volume of saline by gavage for 30 days (Figure 1).
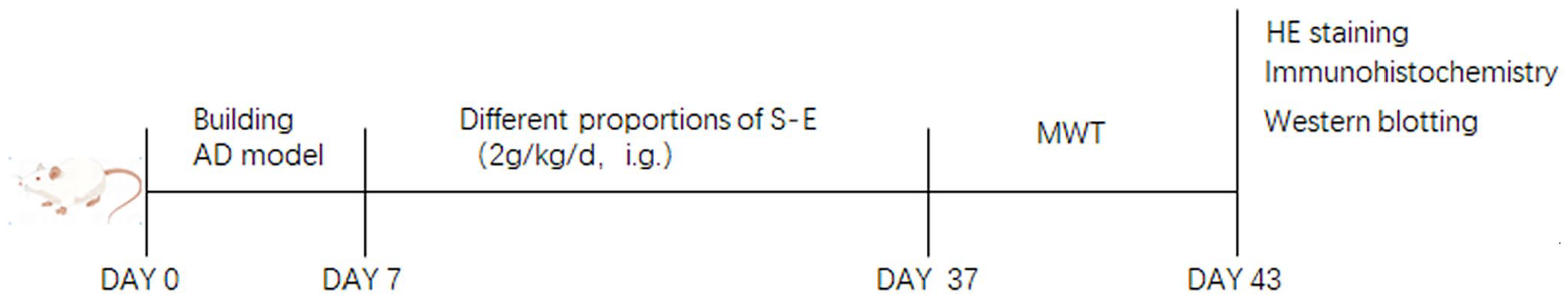
Experimental flow chart. An AD rat model was established by intracerebroventricular injection of Aβ1-42 during days 0 to 7. From day 7 to day 37, model rats were subjected to oral gavage with different proportions of Schisandra-Evodia. On days 37 to 43, the Morris water maze experiment was conducted. Following the behavioral experiments, histological analyses, including HE staining and immunohistochemistry, as well as biochemical assays using Western blotting, were performed.
Morris water maze test
The Morris water maze test was used to evaluate the learning and memory ability of the rats. The rats were positioned to navigate the experiment for five consecutive days, starting from day 37. The experiment was conducted in a circular pool (150 cm in diameter, 50 cm high, and 30 cm deep) divided into four quadrants, with the platform placed 1 cm underwater in the fourth quadrant. The rats were then released into the second quadrant. The time required to swim to the fourth quadrant was also recorded. If the rat did not find the platform within 90 s, it was moved to the platform for 20 s. After five days, a spatial exploration experiment was performed. The platform was removed and the number of times the mouse crossed the original platform position within 90 s was recorded. All experiments were performed using video tracking equipment for data acquisition and analysis with behavioral analysis software (Techman soft, WMT-100, Chengdu, China).
Brain tissue preparation
After behavioral testing, all rats were cervically dislocated and sacrificed, and their brains were immediately removed. The hippocampi of six rats in each group were dissected out and stored at −80°C until biochemical studies were performed. Before the assay, each portion of brain tissue was rapidly homogenized in ice-cold saline and the homogenate was centrifuged at 3500 rpm for 15 min at 4°C. The supernatant was collected for further analysis. The whole brains of the other rats in each group were placed in paraformaldehyde at a mass concentration of 4% for 24 h. The tissue was dehydrated in gradients of 20% and 30% sucrose for 24 h until it sank to the bottom of the bottle, removed, and then freeze-embedded with an optimal cutting temperature compound (OCT) embedding agent for sectioning.
HE staining
The OCT-embedded brain tissue was cut into 5-µm sections, rinsed with running water for one minute, immersed in hematoxylin and eosin (HE) staining solution for five minutes, rinsed with running water again for one minute, rinsed with 1% hydrochloric acid-ethanol solution for five seconds, rinsed with running water for 30 s, and saturated with Li2 CO3 solution which returned to blue for five seconds. Afterward, the tissue was rinsed with running water for 30 s, stained with eosin for 10 s, rinsed with running water for one minute, sliced with 80%, 90%, and anhydrous ethanol of gradient ethanol dehydration, two xylene solutions for three minutes each, sealed in neutral resin, dried in an oven at 65°C, and finally observed and photographed using light microscopy.
Immunohistochemistry
OCT-embedded brain tissue was cut into 5-µm sections. Sections were incubated in a peroxidase blocker containing 3% H2O2 for 30 min to block endogenous peroxidase activity and nonspecific antigens. After washing, sections were incubated in normal goat serum for 15 min. Subsequently, sections were incubated overnight in primary antibodies specific for Aβ1-42 (1:400, PhD, Wuhan, China) and P-Tau (1:200, PhD, Wuhan, China). After washing, sections were incubated with biotinylated secondary antibodies at 37°C. Diaminobenzidine development and hematoxylin counterstaining were performed, followed by dehydration, transparency, and sectioning. Positive cells were stained brown and observed at magnification (400×).
Western blot analysis
Total rat brain tissue protein was extracted and measured using a BCA assay kit (Beyotime, Suzhou, China). Equal amounts of protein buffer were separated using 12% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred to 0.22 µm NC membranes. Membranes were blocked with 5% (w/v) skim milk for two hours in phosphate-buffered saline with Tween 20 (PBST). All primary antibodies were incubated overnight at 4°C. The following were used: BDNF (1:1,000), CREB and p-CREB (1:2,000), TrkB and P-TrkB (1:2,000), GSK-3β (1:5,000), Tau and P-Tau (1:2,000), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:10,000). After washing with PBST, the membranes were incubated with secondary rabbit antibodies for one hour at room temperature. After washing with PBST, protein bands were visualized using an ECL kit (Beyotime, Suzhou, China). The bands were scanned using a molecular imager (BIO-RAD, Hercules, CA, USA).
Statistical analysis
All values were expressed as mean ± standard deviation and analyzed with graph pad Prism 8.0. Comparison of data from multiple groups against one group was performed using one-way analysis of variance (ANOVA), followed by Dunnett’s post hoc test or two-way repeated-measures ANOVA with Tukey’s multiple comparison tests. The level of significance was set at P < 0.05.
Results
Schisandra-Evodia improves spatial memory impairment in AD rat models
To investigate the effect of different ratios of Schisandra-Evodia on spatial memory impairment in AD rat models, we conducted a Morris water maze experiment and excluded the effect on rat locomotor ability on the experimental results (P > 0.05, Figure 2(A)). The results of the localization navigation experiment showed that the model group took a long time to reach the platform compared with the control group, and the difference was significant (P < 0.01, Figure 2(B)). Meanwhile, the mean latency of each Schisandra-Evodia treatment group was shortened compared with the model group, and the difference was significant (P < 0.05, Figure 2(B)), and the latency of the Schisandra-Evodia treatment group with a 2:1 ratio was highly significant compared with the model group (P < 0.01). In the spatial exploration experiment, the number of rats crossing the original platform significantly decreased in the model group compared with the control group (P < 0.05, Figure 2(C)), whereas after the Schisandra-Evodia treatment, the number of rats crossing the original platform significantly increased (P < 0.05, Figure 2(C)); the 2:1 group demonstrated no significant difference in the treatment effect compared with the control group. The results of the spatial exploration and localization navigation experiments showed that different ratios of Schisandra-Evodia administration improved spatial memory impairment in AD rat models, with the strongest effect being observed at a ratio of 2:1.
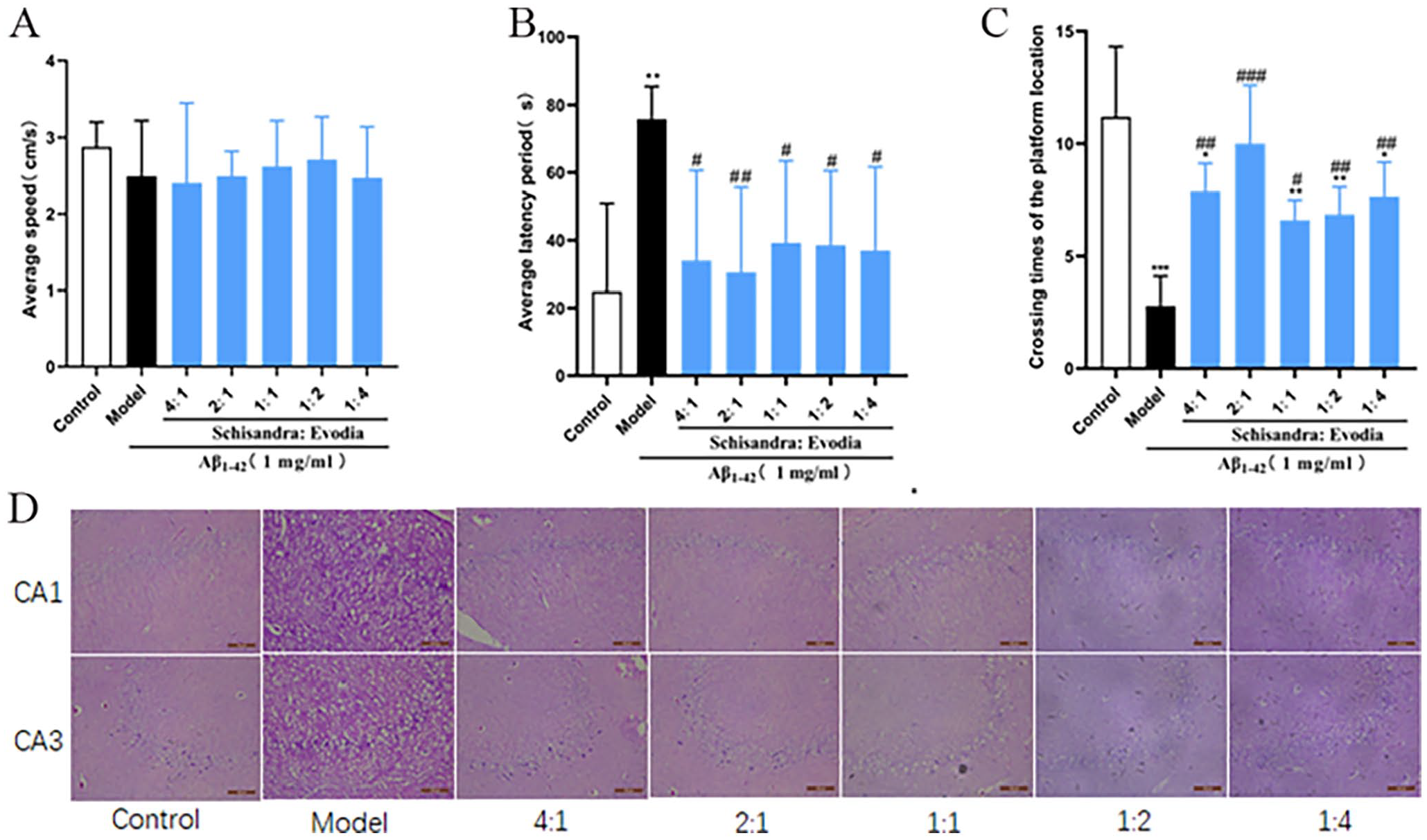
Neuroprotective effect of Schisandra-Evodia on Aβ1-42 induced AD rat model: (A) mean swimming speed of rats in the water maze experiment, (B) latency of rats in the water maze experiment, (C) number of times rats crossed the platform location in the positioning navigation experiment, and (D) HE-stained rat brain tissue sections (data are expressed as mean ± standard deviation (n = 6–8 per group,*P < 0.05, **P < 0.01, ***P < 0.001, compared with the control group; #P < 0.05, ##P < 0.01, ###P < 0.001, compared with the model group).
In the HE staining experiments, the neuronal cells in the hippocampal area of the control group had regular morphology, were neatly and tightly arranged, and showed no apparent inflammatory infiltration. Compared with the control group, the neuronal cells in the hippocampal area of the model group were irregular in morphology, loosely arranged, and deformed, with large cell gaps, apparent tissue cavities, and evident inflammatory infiltration. The 4:1, 2:1, and 1:1 groups had more regular cell morphology, neatly arranged cells, and a small amount of inflammatory infiltration. The 1:2 and 1:4 groups had slightly irregular cell morphology and arrangement, a small amount of inflammatory infiltration, and a few tissue cavities. This indicated that the protective effect of Schisandra on neuronal cells was stronger than that of Evodia. In conclusion, Schisandra-Evodia ameliorated spatial memory impairment in AD rat models by protecting against neuronal damage, with an optimal therapeutic effect observed at a ratio of 2:1.
Schisandra-Evodia facilitates the clearance of Aβ1-42 protein
To investigate the effects of different ratios of Schisandra-Evodia on the Aβ1-42 protein in the brains of AD model rats, immunohistochemistry experiments were conducted (Figure 3). The results showed (Figure 3(B)) a significant increase in the levels of the Aβ1-42 protein in the model group compared to the control group. In contrast to the model group, all treatment groups exhibited a significant reduction in Aβ1-42 protein levels. Among them, the 2:1 group demonstrated a significantly enhanced clearance effect on the Aβ1-42 protein compared to the other treatment groups, with a significant difference (P < 0.05).
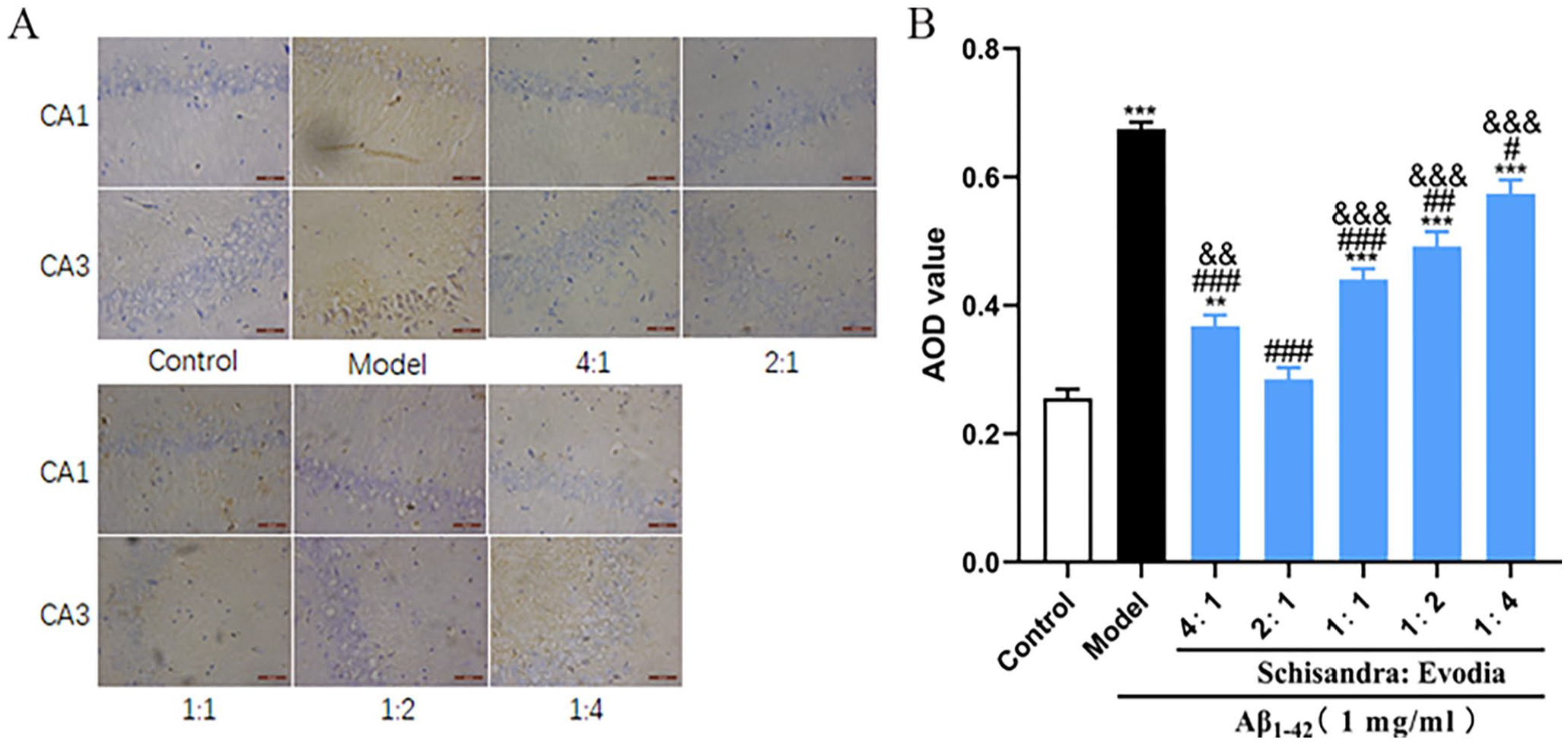
Different ratios of Schisandra-Evodia (S-E) promote the clearance of the Aβ1-42 protein in the hippocampal region of AD rat models: (A) immunohistochemical assay for the Aβ1-42 protein, (B) quantitative analysis of Aβ1-42 protein; (data are expressed as mean ± standard deviation (n = 3 per group, scale: 100 µm, *P < 0.05, **P < 0.01, ***P < 0.001, compared with the control group; #P < 0.05, ##P < 0.01, ###P < 0.001, compared with the model group; &P < 0.05, &&P < 0.01, &&&P < 0.001, compared with the 2:1 group).
Schisandra-Evodia activates the BDNF/TrkB/CREB signaling pathway
To explore the molecular mechanism of Schisandra-Evodia inhibition of the Aβ1-42 protein, we examined the effect of Schisandra-Evodia on the BDNF/TrkB/CREB signaling pathway. Western blot (WB) experimental results showed that each Schisandra-Evodia treatment group upregulated BDNF, P-TrkB/TrkB, and P-CREB/CREB levels compared to the model group (Figure 4). Among them, there was no significant difference in the increase in the expression levels of BDNF, P-TrkB/TrkB, and P-CREB/CREB in the 4:1 group compared to the 2:1 group. Meanwhile, the agonistic effect of other Schisandra-Evodia administration groups on the BDNF/TrkB/CREB pathway decreased with a decrease in the percentage of Schisandra, which may indicate that Schisandra played a greater role in upregulating BDNF/TrkB/CREB pathway activity.
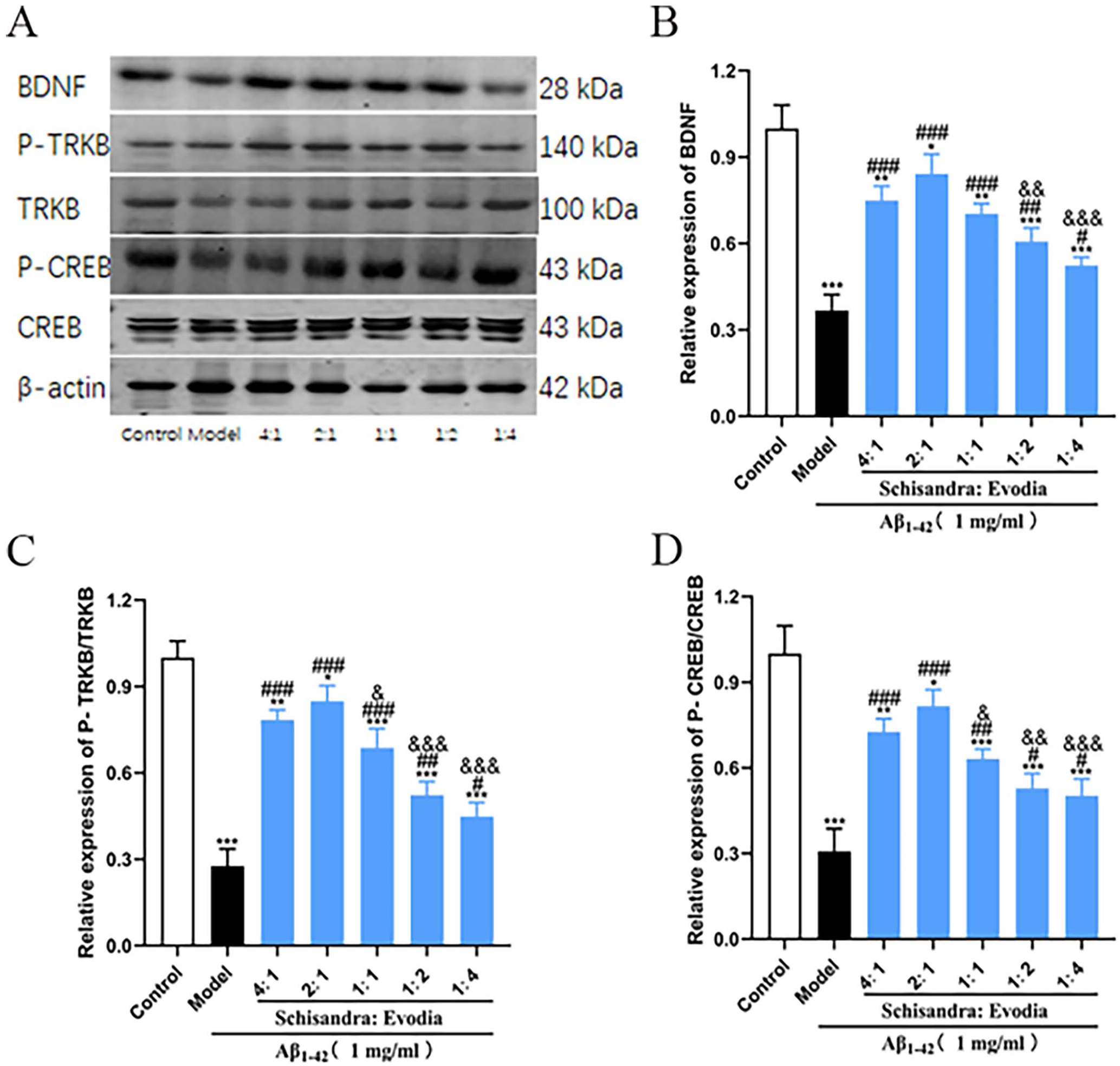
Effect of different ratios of Schisandra-Evodia on the expression of BDNF, TrkB, P-TrkB, CREB, and P-CREB proteins: (A) effect of different ratio groups of Schisandra-Evodia on the protein expression of BDNF, TrkB, P-TrkB, CREB, and P-CREB, (B) relative expression of BDNF, (C) relative expression of P-TrkB/TrkB, (D) relative expression of P-CREB/CREB; data are expressed as mean ± standard deviation (n = 3 per group, scale: 50 µm, *P < 0.05, **P < 0.01, ***P < 0.001, compared with the control group; #P < 0.05,##P < 0.01,###P < 0.001, compared with the model group; &P < 0.05, &&P < 0.01, &&&P < 0.001, compared with the 2:1 group).
Schisandra-Evodia inhibits P-Tau protein production
After studying the Aβ pathology, we investigated the effect of different ratios of Schisandra-Evodia on the P-Tau protein in the brain of the AD rat model and the results of immunohistochemical experiments showed (Figure 5(B)) that P-Tau protein expression was significantly upregulated in the model group compared with the control group. P-Tau protein expression was significantly inhibited in all administered groups compared to the model group. The inhibitory effect on the P-Tau protein was significantly downregulated in the 4:1 and 1:1 groups compared to the 2:1 group (P < 0.05), while the 1:2 and 1:4 groups showed no significant difference compared to the 2:1 group.
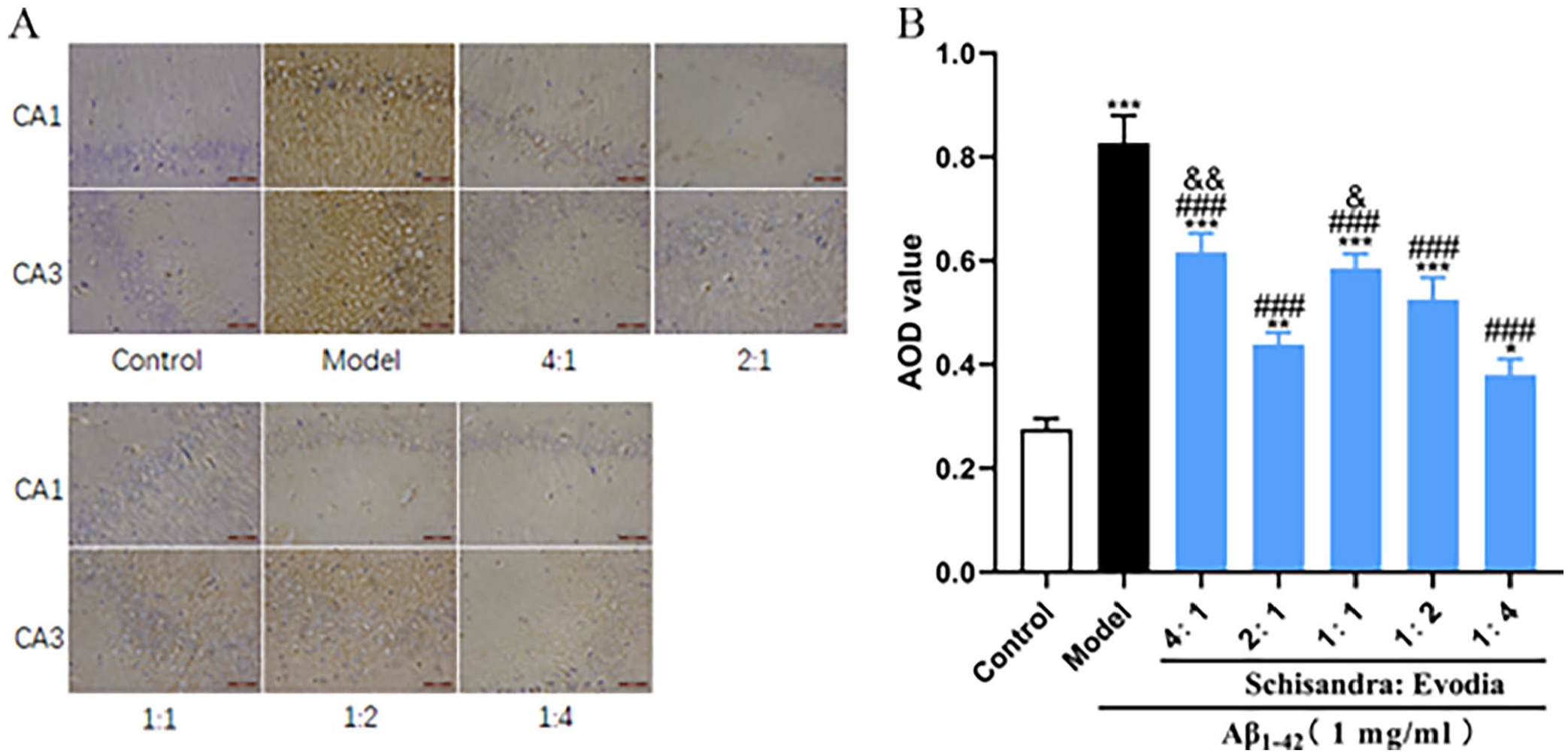
Different ratios of Schisandra-Evodia inhibit the production of the P-Tau protein in the hippocampus of AD rat models: (A) immunohistochemical assay for the P-Tau protein, (B) quantitative analysis of the P-Tau protein; (data are expressed as mean ± standard deviation (n = 3 per group, scale: 50 µm, *P < 0.05, **P < 0.01, ***P < 0.001, compared with control group; #P < 0.05, ##P < 0.01, ###P < 0.001, compared with the model group, &P < 0.05, &&P < 0.01, &&&P < 0.001, compared with the 2:1 group).
Schisandra-Evodia inhibits GSK-3β/Tau pathway activity
To explore the molecular mechanism of Schisandra-Evodia inhibition of the P-Tau protein, we examined the effect of Schisandra-Evodia on the GSK-3β/Tau signaling pathway. The results of the WB experiments showed that each Schisandra-Evodia treatment group inhibited GSK-3β and P-Tau/Tau expression compared to the model group (Figure 6). Notably, the 1:4 group seemed to have a more profound inhibitory effect on the GSK-3β/Tau signaling pathway compared with the 2:1 group (Figure 6).
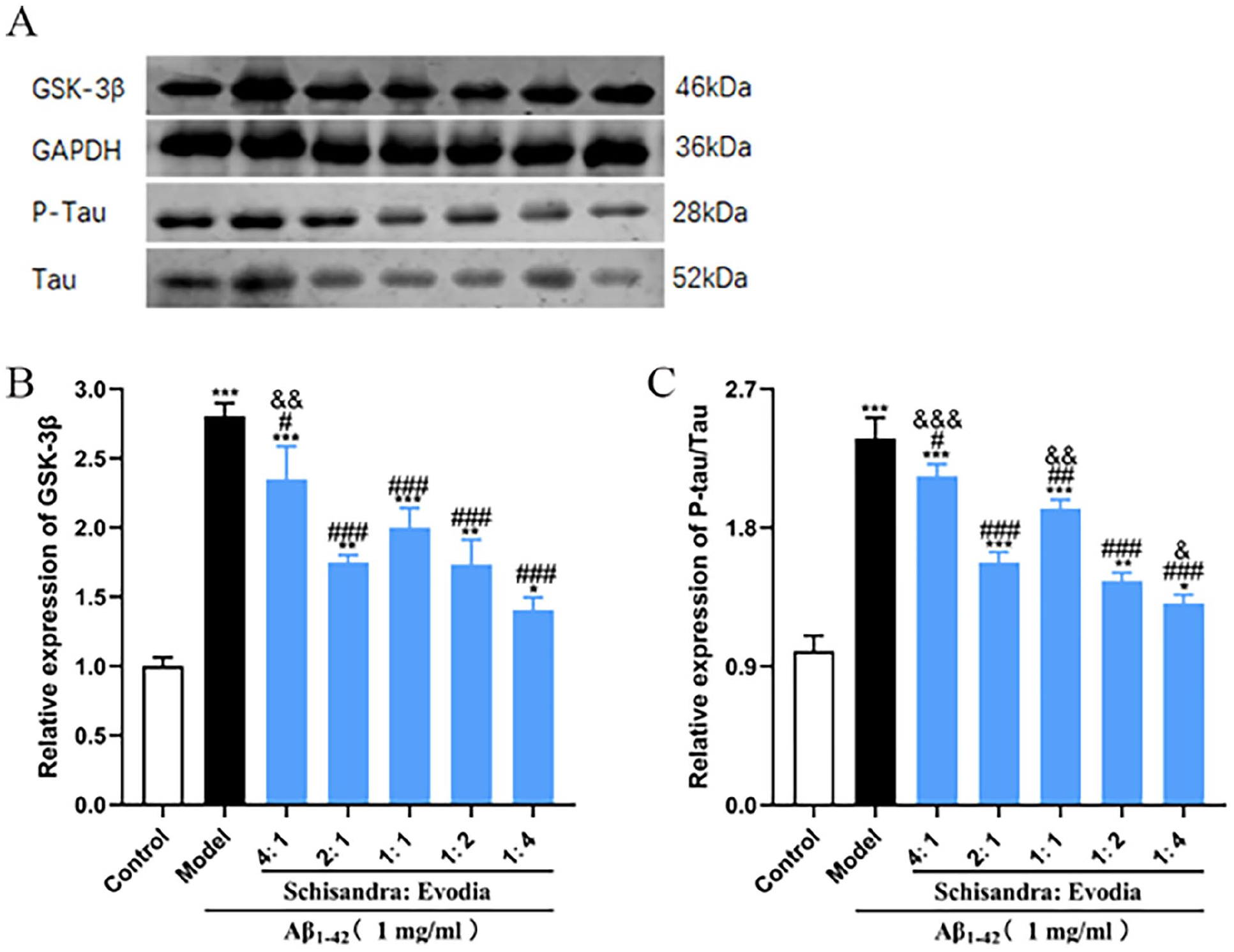
Effect of different ratios of Schisandra-Evodia on GSK-3β and P-Tau/Tau protein expression: (A) effect of different Schisandra-Evodia ratio groups on GSK-3β and P-Tau/Tau protein expression, (B) GSK-3β relative expression, (C) P-Tau/Tau relative expression; data are expressed as mean ± standard deviation (n = 3 per group, *P < 0.05, **P < 0.01, ***P < 0.001, compared with the control group; #P < 0.05, ##P < 0.01, ###P < 0.001, compared with the model group; &P < 0.05, &&P < 0.01, &&&P < 0.001, compared with the 2:1 group).
Discussion
Recently, Schisandra and Evodia have received considerable attention for their therapeutic potential against AD. 20 However, there is a gap in the research on the therapeutic use of these two drugs in combination with AD. Our study is the first to report the improvement of AD pathological proteins and cognitive functions by Schisandra-Evodia at different ratios, as well as the molecular mechanism of action of AD. In particular, we found that Schisandra-Evodia modulated both the BDNF/TrkB/CREB and GSK-3β/Tau pathway activities, thereby reducing Aβ1-42 and P-Tau protein levels in the hippocampus of rats (Figure 7). Notably, this effect seemed to vary with the proportion of the two herbs. In addition, through a comprehensive analysis of different Schisandra-Evodia ratios for the treatment of AD in model rats, we concluded that a Schisandra-Evodia ratio of 2:1 had a more profound therapeutic effect on AD in general.
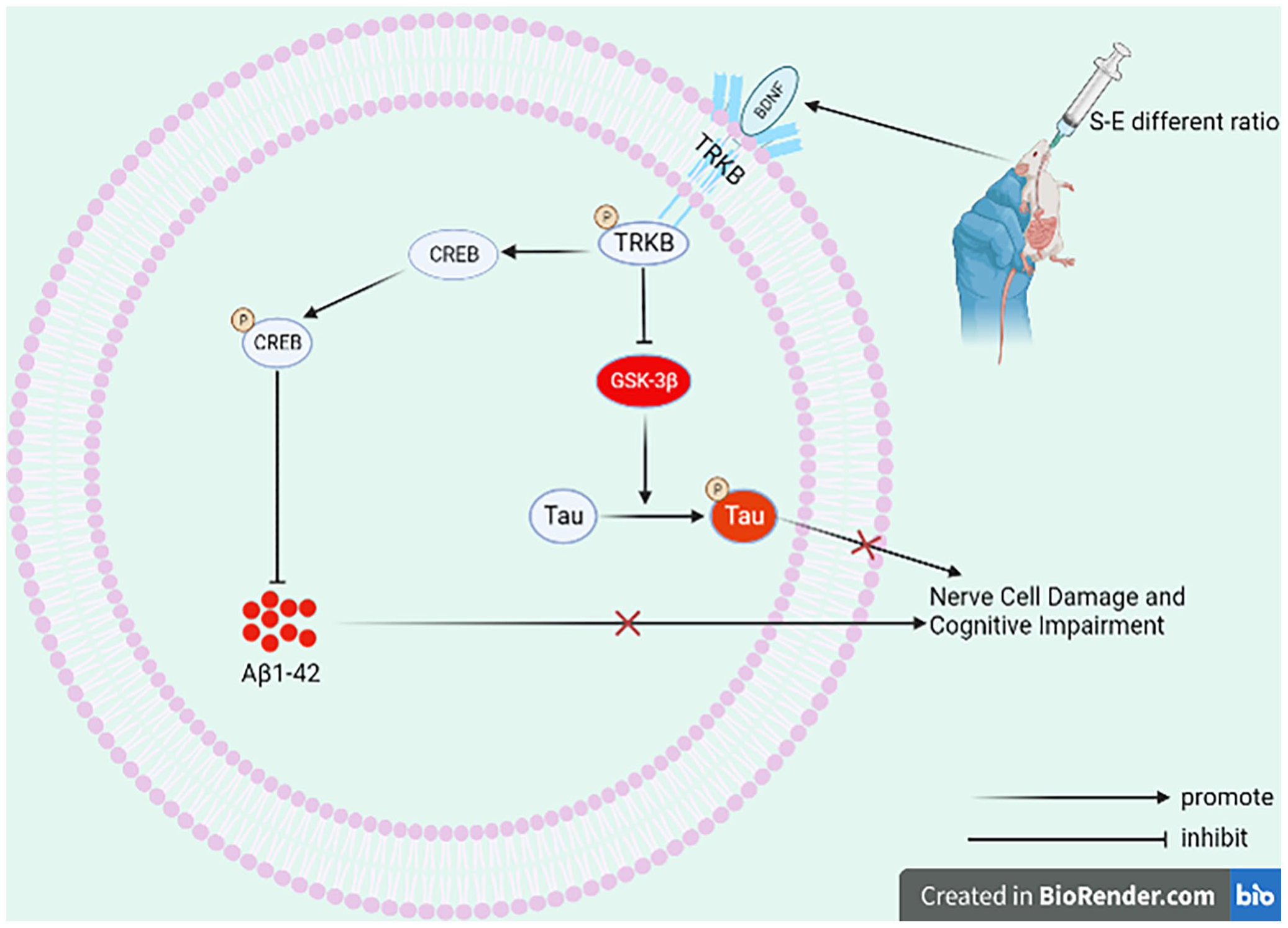
Mechanism of action of Schisandra-Evodia to improve neuronal damage and cognitive impairment in AD (created with BioRender.com).
To study the effects of Schisandra-Evodia on AD, a reliable animal model needed to be developed. After reviewing the literature, we found that a lateral ventricular injection of Aβ1-42 treatment resulted in a rat model of AD that exhibited a significant decrease in spatial memory, increased Aβ deposition, and increased Tau levels, 21 which is consistent with our study. As measured by Morris water maze and HE experiments, we found higher spatial memory capacity and neuronal cell damage in the model group rats, and immunohistochemical experiments showed a significant increase in Aβ1-42 and P-Tau protein levels in the model group rats; these results suggest that our modeling was a success.
Unlike other traditional herbal therapies around the world, traditional Chinese medicine is unique in that a variety of herbal therapies are developed based on unique theories and healing modalities regarding it. 15 Among them, herbal pairs, one of the most basic and common forms of herbal therapies, not only have the basic functions and main treatments of multiple herbal therapies but also facilitate the in-depth study of their mechanisms of action. For example, Qi et al. inhibited neuroinflammation and apoptosis in AD mice using a medicine synthesized from the combination of Alpinia Oxyphylla and Schisandra chinensis. 22 By reviewing traditional Chinese medicine texts and collecting clinical compound prescriptions, we determined that Schisandra, in combination with Evodia, has therapeutic effects on AD. However, in clinical use, traditional Chinese medicine practitioners are always flexible in changing the ratio of herbs to respond to the multiple needs of patients. Therefore, we investigated the effects of different ratio groups of Schisandra-Evodia on Aβ1-42-induced AD model rats. Through the Morris water maze and HE experiments, we found that Schisandra-Evodia was effective in improving spatial memory impairment and neuronal cell damage in AD rat models. Among the different Schisandra-Evodia ratios, we found that a 2:1 ratio had a more profound therapeutic effect on AD.
The Aβ protein is one of the most essential causative factors in the etiology of AD, and studies of clinical imaging and biomarkers in AD patients have conclusively confirmed the central role of Aβ in the development and progression of AD and in triggering the spread of neurodegeneration. 23 Aβ1-42 is the most misfolded and aggregated form of the Aβ protein monomer and is the major form of seed starch aggregates and oligomers. 24 In our study, each Schisandra-Evodia treatment group significantly reduced Aβ1-42 protein levels in AD rat models, and, notably, the inhibitory effect of Schisandra-Evodia on the Aβ1-42 protein was downregulated with the decrease in the percentage of Schisandra, which may be caused by the inhibitory effect of Schisandra-rich lignans on the Aβ protein. 25 Schisandra has been reported to produce antidepressant-like effects via the BDNF/TrkB/CREB pathway in mice, which are induced by repetitive corticosterone. 11 The lignan-like component Macranthol is able to exert antidepressant-like effects through the BDNF-TrkB-PI3K/Akt signaling pathway 26 ; therefore, we examined the expression of proteins related to the BDNF/TrkB/CREB pathway, and the results of WB experiments showed that the expression of BDNF, P-TrkB/TrkB, and P-CREB/CREB increased significantly under Schisandra-Evodia treatment. Furthermore, the stimulatory effect of Schisandra-Evodia on the BDNF/TrkB/CREB pathway was enhanced with the increase in the percentage of Schisandra, which may imply that Schisandra plays a more profound role in stimulating the BDNF/TrkB/CREB pathway. Our experimental results demonstrate that S-E reduces Aβ1-42 protein levels by stimulating the BDNF/TRKB/CREB pathway, thereby mitigating neuronal damage and cognitive impairment caused by AD. In addition, among the different ratios of Schisandra-Evodia, the 2:1 group exhibited the most profound agonistic effect on the BDNF/TrkB/CREB pathway and the strongest scavenging effect on the Aβ1-42 protein.
The susceptibility risk factors for AD pathogenesis are diverse. Therefore, targeting multiple interconnected pathological networks may be an effective strategy for treating AD. Aβ has been reported to be located upstream of Tau in AD pathogenesis, either directly or indirectly, and triggers the transition of Tau from the normal to the toxic state of the P-Tau protein. 27 However, there is also evidence that P-Tau can enhance Aβ toxicity through a feedback pathway 28 ; we, therefore, examined the effect of Schisandra-Evodia on the P-Tau protein and its molecular mechanism. WB and immunohistochemical results showed that Schisandra-Evodia significantly downregulated GSK-3β as well as P-Tau/Tau protein expression and reduced P-Tau protein levels in the hippocampus, suggesting that Schisandra-Evodia reduces Tau protein phosphorylation by inhibiting GSK-3β/Tau pathway activity and reducing P-Tau protein production. However, the inhibitory effect on the P-Tau protein was upregulated with an increase in the content of Evodia in the different Schisandra-Evodia ratio groups. In terms of reducing P-Tau/Tau expression, the 1:4 group, rather than the 2:1 group, demonstrated the most profound inhibitory effect. Furthermore, in previous studies, evodiamine, rutaecarpine, and dehydroevodiamine contained in Evodia exhibited inhibitory effects on Tau protein hyperphosphorylation;20,29 these results suggest that alkaloid components in Evodia play a major role in reducing Tau protein phosphorylation.
In summary, our study demonstrates that Schisandra-Evodia can reduce the levels of Aβ1-42 and P-Tau proteins by modulating the activity of the BDNF/TrkB/CREB and GSK-3β/Tau pathways, thereby ameliorating neuronal cell damage and cognitive deficits caused by AD. The therapeutic effect on AD varies with the herbal dosage. In addition, we showed that, in general, the Schisandra-Evodia2:1 group has a better therapeutic effect on AD, which will provide an experimental basis and guidance for clinical use. However, there are still some limitations in our study; for example, we did not conduct in vitro experiments to further investigate the specific molecular mechanism of the BDNF/TrkB/CREB pathway to inhibit the Aβ protein, nor did we conduct in vitro experiments to exclude the effect of pharmacokinetics on Schisandra-Evodia dosing, which will be our next research direction.
Footnotes
Acknowledgements
The authors would like to express their gratitude to Xu Yuan and Yuanjin Chen for their suggestions regarding statistical analysis. They also thank Kemeng Liu, Ruijiao Wang, and Chengguo Pang for their assistance and guidance during the experiments.
Authors’ Contributions
All authors participated in the design and interpretation of the studies, data analysis, and manuscript review. CQ and JL conducted the experiments. HH and PD supplied all reagents and animals. CQ wrote the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Heilongjiang Provincial Natural Science Foundation Joint Guidance Project (grant no: LH2019H053) and the Science Foundation Project of Heilongjiang University of Traditional Chinese Medicine (grant no: 2017XY01).