
Abstract
Objective
Tobacco smoke exposure is a major risk factor for aortic aneurysm development. However, the initial aortic response to tobacco smoke, preceding aneurysm formation, is not well understood. We sought to create a model to determine the effect of solubilized tobacco smoke (STS) on the thoracic and abdominal aorta of mice as well as on cultured human aortic smooth muscle cells (HASMCs).
Methods
Tobacco smoke was solubilized and delivered to mice via implanted osmotic minipumps. Twenty male C57BL/6 mice received STS or vehicle infusion. The descending thoracic, suprarenal abdominal, and infrarenal abdominal segments of the aorta were assessed for elastic lamellar damage, smooth muscle cell phenotype, and infiltration of inflammatory cells. Cultured HASMCs grown in media containing STS were compared to cells grown in standard media in order to verify our in vivo findings.
Results
Tobacco smoke solution caused significantly more breaks in the elastic lamellae of the thoracic and abdominal aorta compared to control solution (P< .0001) without inciting an inflammatory infiltrate. Elastin breaks occurred more frequently in the abdominal aorta than the thoracic aorta (P < .01). Exposure to STS-induced aortic microdissections and downregulation of α-smooth muscle actin (α-SMA) by vascular smooth muscle cells (VSMCs). Treatment of cultured HASMCs with STS confirmed the decrease in α-SMA expression.
Conclusion
Delivery of STS via osmotic minipumps appears to be a promising model for investigating the early aortic response to tobacco smoke exposure. The initial effect of tobacco smoke exposure on the aorta is elastic lamellar damage and downregulation of (α-SMA) expression by VSMCs. Elastic lamellar damage occurs more frequently in the abdominal aorta than the thoracic aorta and does not seem to be mediated by the presence of macrophages or other inflammatory cells.
Introduction
Exposure to tobacco smoke is a major risk factor for aortic aneurysm formation.1,2 However, the early effects of tobacco smoke exposure on the aorta, prior to aneurysm formation, are not well understood. Identifying the features of early tobacco smoke–induced aortic damage is clinically relevant, as no medical therapy is effective in preventing aortic aneurysm growth after aneurysm formation. We therefore developed a novel model of early tobacco smoke–induced aortic injury, using solubilized tobacco smoke (STS) administered as a continuous infusion from implantable osmotic minipumps, in order to identify the earliest aortic response to tobacco smoke exposure.
The existing animal models of aortic aneurysm consist of angiotensin II infusion, elastase perfusion, and periadventitial elastase or calcium chloride application.3-5 Inhaled tobacco smoke exposure and nicotine infusion have also been combined with existing models of aortic aneurysm.6-8 These models reliably and quickly produce the aneurysm phenotype and allow for investigation of potential therapies. However, the initial aortic injury that occurs prior to aneurysm formation and the contribution of tobacco smoke exposure to aortic aneurysm formation is not easily investigated with existing models.
Genetic models of aortic aneurysm include loss-of-function models for Fbn1, Tgfbr1 or Tgfbr2, which predominantly result in aortic aneurysms confined to the thoracic aorta, or require additional insults, such angiotensin II infusion, to produce abdominal aortic aneurysms. 9 The use of genetically susceptible mice and the use of additional exposures makes it difficult to discern the mechanism by which tobacco smoke exposure contributes to aortic injury and aneurysm formation.
Our goal was to develop a model of tobacco smoke–induced aortic injury using wild-type mice so that the effects of tobacco smoke would be more easily discerned without confounding effects from genetic abnormalities. Our model was designed to characterize early aortic injury that precedes aneurysm formation.
We also sought to compare the susceptibility of the thoracic and abdominal aorta to tobacco smoke–mediated damage. While tobacco smoke exposure is a risk factor for both thoracic and abdominal aortic aneurysms, smoking is more strongly associated with abdominal rather than thoracic aortic aneurysms. The prevalence of abdominal aortic aneurysm has decreased with decreasing smoking patterns, while the prevalence of thoracic aortic aneurysm has increased, underscoring the stronger association of tobacco smoke with abdominal aortic aneurysm.10-13 We hypothesized that regional differences in aortic structure may make the abdominal aorta more susceptible to tobacco smoke–mediated injury than the thoracic aorta. We therefore compared the effect of tobacco smoke exposure on the descending thoracic aorta, suprarenal abdominal aorta, and infrarenal abdominal aorta in our model.
Materials and Methods
Tobacco Smoke Solution
Tobacco smoke was solubilized in phosphate buffered saline (PBS) solution by drawing the smoke over the PBS at a pressure of 3–5 mmHg to simulate typical tobacco smoke inhalation in humans. The concentration of nicotine within the smoke solution was estimated by comparing the absorbance peak of nicotine in the smoke solution to the absorbance peak of known concentrations of nicotine (figure 1). The samples were reconstituted in 1 mL PBS and analyzed on a Cary 4000 UV/VIS spectrophotometer over a wavelength range of 200–600 nm. Ultraviolet absorbance spectra of solubilized tobacco smoke and nicotine solution demonstrate a peak at 260 nm corresponding to nicotine. The nicotine concentration of the STS solution was determined to be 310 μg/mL. 10 μL of STS solution dried and diluted into 1 mL PBS (red line), yielded an equivalent peak as 3.1 μg/mL of nicotine (blue line). The presence of additional tobacco smoke components in the STS solution is demonstrated by its wider absorbance spectra compared to the absorbance spectra of the nicotine solution.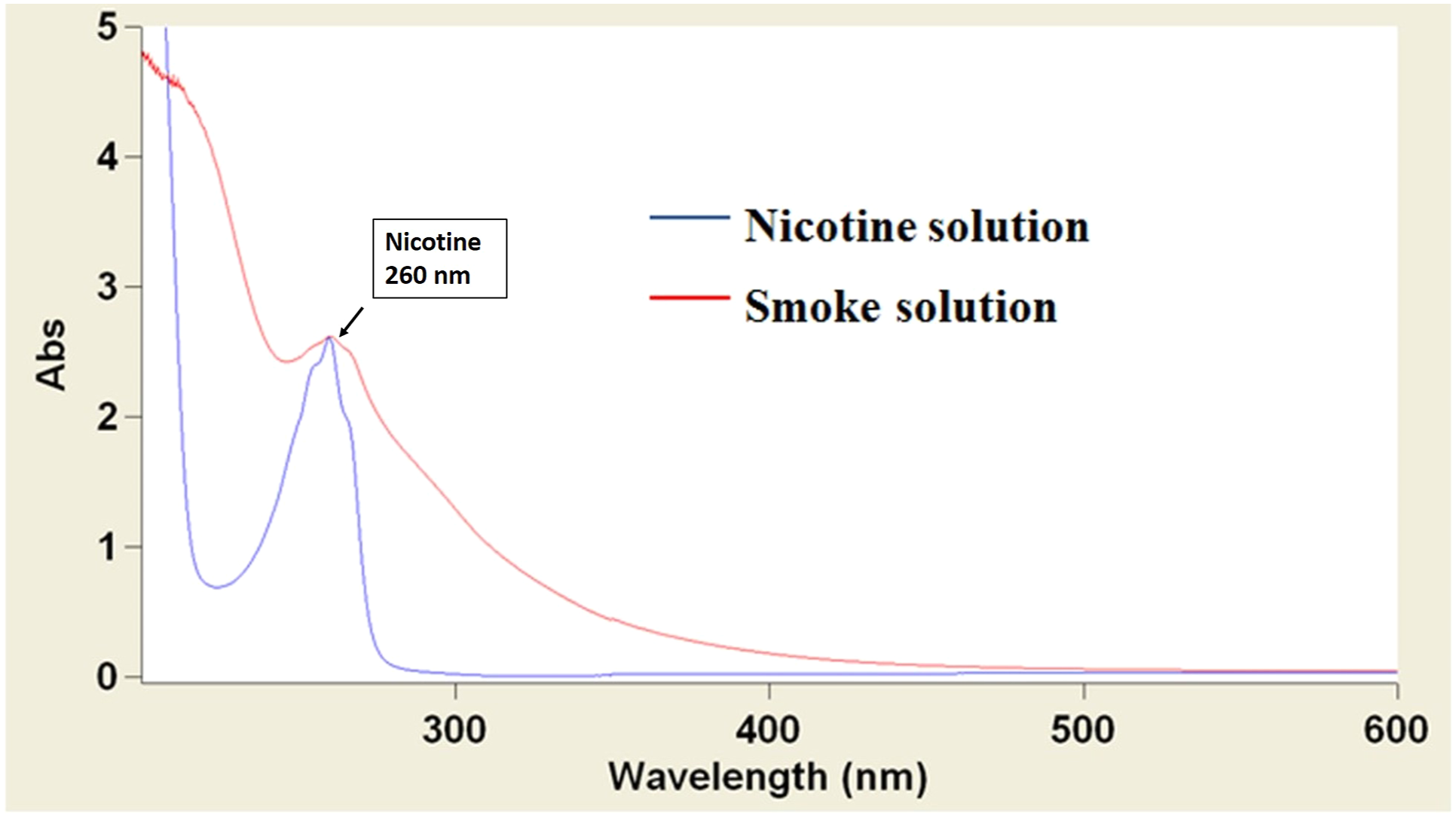
Surgical Protocol
All animals were maintained in accordance with the Guide for the Care and Use of Laboratory Animals and the Oregon Health and Science University approved all IACUC protocols. Male C57BL/6 mice, age 10 or 32 weeks, were obtained (Jackson Laboratories. Bar Harbor, ME). Analysis of young (10 wk) vs older (32 wk) mice was performed to determine age-related differences in the number of aortic elastin breaks. No baseline differences in number of elastin breaks were seen between young and older mice (results not shown). Tobacco smoke solution or vehicle (PBS) was loaded into osmotic minipumps (Alzet 2006, Alzet Cupertino, CA) and surgically implanted into the subcutaneous tissue or inserted into the peritoneal cavity. A total of 200 µl of solution was infused at a rate of .25 μL/hr. The peritoneal route of tobacco smoke solution administration was used to determine whether direct administration of the smoke solution adjacent to the abdominal aorta via the posterior peritoneum would have an increased effect on the aorta compared to the subcutaneous infusion. Seven mice were treated with control solution pumps, 8 mice were treated with tobacco smoke solution pumps in the peritoneal position, and 5 mice were treated with tobacco smoke solution in the subcutaneous position.
Animals received a total infusion of 200 μL over a 4-week period. Animals were anesthetized using sevoflurane at the time of pump insertion and at 4 weeks when the animals were sacrificed. Postoperative analgesia was managed with buprenorphine using a pain assessment protocol. The aorta was flushed with 4% paraformaldehyde via the left ventricle. The heart and aorta were removed en bloc to the aortic bifurcation, the left kidney and renal artery were left attached for orientation. The aorta was preserved in 4% paraformaldehyde for 2 hours followed by a 10% sucrose solution until tissue sectioning.
Tissue Preparation
Perfusion-fixed aortic tissue was divided into descending thoracic, suprarenal abdominal, and infrarenal aortic sections. Histologic analysis was performed on fifteen sections, taken from 3 to 5 distinct regions of each aortic segment.
Histochemical and Immunohistochemical Staining
Trichrome and Verhoeff-Van Gieson (VVG) stains were used to visualize collagen and elastin fibers respectively. Monoclonal antibodies recognizing myeloperoxidase, CD-3, and CD-20 were used to assess for neutrophils, T and B lymphocytes, respectively.
Immunofluorescence microscopy with Mac-2 antibodies (Cedarlane, Burlington NC, CAT# CL8942AP) and antibodies to α-smooth muscle actin (α-SMA) (Sigma, Santa Fe NM, CAT# A2547-.2 ML) were used to evaluate macrophage infiltration and smooth muscle cells respectively. Autofluorescence of elastin was also evaluated.
Micrographs were taken using a Zeiss Axiophot (light microscopy) or Axiovert microscope (fluorescence microscopy).
Election Microscopy
Dissected regions of mouse aortae from 2 mice were fixed for a minimum of 12 hours at 4°C in 1.5% glutaraldehyde/1.5% formaldehyde with .05% tannic acid then rinsed in Dulbecco’s serum free media. Subsequently, they were post-fixed for 24 hours in media buffered 1% osmium tetroxide. Assisted by a Pelco Biowave Pro microwave processor, tissues were dehydrated in a graded series of ethanol (30%–100% at 100W for 1 min/step), rinsed in propylene oxide (2x at 100W for 1 min/step), infiltrated in 1:1 and 1:3 propylene oxide: Spurr’s epoxy (250W for 15 min/step) then in 3 changes of 100% Spurr’s epoxy (250W for 15 min/step followed by overnight at 4oC) before embedding in fresh Spurr’s epoxy. Ultrathin sections were cut at 80 nm, mounted onto 1x2mm single hole formvar-coated slot grids, stained in Uranyl acetate and lead citrate, and then examined using a Thermo Ficher Scientific (Waltham, MA) G20 transmission electron microscope operated at 120 KV.
Quantitation of Breaks in the Elastic Lamellae
Breaks in the elastic lamella were counted by 2 independent, blinded investigators. Discordant counts were recounted by both observers. In order to distinguish breaks from areas with decreased or discontinuous lamellar content, breaks were defined according to the following criteria: 1. Elastin breaks are flanked by continuous, sharply delineated elastic lamellae. 2. Elastin breaks are 5 mm or less in length (40 X magnification viewed at 100% on Windows Photo Viewer, Redmond WA) to assure that the 2 elastin ends are part of the same lamella. 3. Sections with non-continuous elastic lamellae comprising greater than 25% of the section were excluded from analysis, as the presence of continuous elastic lamellae are needed in order to identify elastin breaks.
Cell Culture Experiments
Human aortic smooth muscle cells (HASMCs) were obtained from GIBCO (Cat# C-007-5C) (Waltham, MA) and were maintained in Medium 231 (GIBCO, M231, Carlsbad, CA), supplemented with smooth muscle growth supplement, penicillin, and streptomycin. Six-well plates (for Western blotting) or 1 mL chamber slides (for immunofluorescence) were seeded with 200 000 cells per well and cultured in medium 231 with differentiation supplement (Cascade Biologics, S-008-5, Frederick, MD) for 8 days. Cells were treated 4 days with 1/100 STS in PBS or an equivalent volume of PBS in medium 231 with differentiation supplement. Cells grown in chamber slides were incubated with antibodies to SMA or to MAC-2, followed by washing and incubation with secondary antibodies conjugated with Alexa Fluor 488.
Western Blot
Human aortic smooth muscle cells were harvested by directly lysing in SDS-PAGE loading buffer containing protease inhibitor cocktail (Sigma, P8340, St. Louis, MO), and lysates were stored at −20°C until western blots were performed. After incubating lysates at 95°C for 5 minutes, lysates were separated on 12.5% polyacrylamide gels at 140V for 2 hours. Transfer to nitrocellulose was performed in 10 mM sodium Borate (Sigma, S9640, St. Louis, MO) at 60V for 1hr in an ice bath. Membranes were blocked in 5% milk, TBST (.1% Tween-20). Anti-SMA (Sigma, A2547, St. Louis, MO) was used at 1/5000 in TBST, 1% milk for 3 hours; anti-MAC-2 (Cedarlane, CL8942AP, Burlington, NC) was used at 1/200 in TBST, 1% milk overnight. Secondary antibodies were incubated at room temperature for 1 hour: 1/1000 goat anti-mouse HRP (Millipore, AP308P) for the SMA blot, and 1/500 goat anti-rat HRP (Invitrogen, 62-9520) for the MAC-2 blot. Membranes were washed with TBST after primary antibody incubation and again after secondary antibody incubation. Blots were developed in Opti-4CN (Biorad, 170-8235, Hercules, CA). After drying, the membranes were scanned, and figures were created using Photoshop (Adobe, San Jose, CA).
Statistics
The number of elastin breaks between treatment groups was compared with a Kruskal–Wallis test with a Dunn’s post-test with Bonferroni correction. The number of elastic lamellae in distinct aortic segments was compared using a two-tailed Student’s t-test. Comparison of α-SMA downregulation between in STS and vehicle-exposed mice was performed using a Fischer’s exact test. A P value of less than .05 was used for statistical significance. All analyses were conducted using Stata IC for Windows, version 13.1 (StataCorp, TX, USA).
Results
Characteristics of Solubilized Tobacco Smoke
The absorbance spectra of STS were compared to that of known concentrations of solubilized nicotine (figure 1). Solubilized tobacco smoke demonstrates a wide absorbance spectrum including a peak at 260 nm corresponding to the absorbance peak for solubilized nicotine. The concentration of nicotine within the smoke solution was estimated by comparing the absorbance peak of nicotine in the smoke solution to the absorbance peak of known concentrations of nicotine. The maximum nicotine concentration of the saturated smoke solution was 310 μg/mL, as 10 μL of smoke solution dried and diluted into 1 mL PBS yielded an equivalent peak as nicotine solution at a concentration of 3.1 μg/mL. The wider absorbance spectrum of smoke solution demonstrates the presence of multiple other tobacco smoke components in addition to nicotine (Figure 1).
Tobacco Smoke Infusion Causes Increased Fragmentation of the Elastic Lamellae in the Descending Thoracic and Suprarenal Abdominal Aorta Compared to Control Infusion
Fifteen cross-sections of aorta at each anatomic location (descending thoracic, suprarenal abdominal, and infrarenal abdominal) were examined after VVG staining for elastin. 3/240 thoracic sections and 5/190 suprarenal abdominal sections were excluded due to artifact or discontinuous elastic lamellae in greater than 25% of the section.
Peritoneal STS infusion significantly increased the number of elastin breaks in the thoracic aorta compared to control infusion. Both peritoneal and subcutaneous STS infusion increased the number of elastin breaks compared to control infusion in the suprarenal aorta. The smoke treated animals demonstrated a greater number of elastin breaks in the suprarenal abdominal aorta compared to the descending thoracic aorta. Control animals demonstrated no difference in the number of elastin breaks between the suprarenal abdominal and descending thoracic aorta (Figure 2). Mice were exposed to vehicle-alone minipumps (n = 5), peritoneal STS-filled minipumps (n = 8), or subcutaneous STS-filled minipumps (n = 5). The thoracic aorta of mice exposed to peritoneal STS and the suprarenal abdominal aorta of mice exposed to either peritoneal or subcutaneous STS demonstrate increased elastin breaks compared to mice exposed to vehicle (P < .0001). The number of elastin breaks in the abdominal aorta is greater than the number of elastin breaks in the thoracic aorta for both peritoneal and SC smoke exposure groups (P =.013 and < .0001, respectively). Data represent median, 25th percentile, and 75th percentile. The whiskers are 75th percentile + 1.5*IQR and 25th percentile − 1.5*IQR. Outlier values not shown in plot.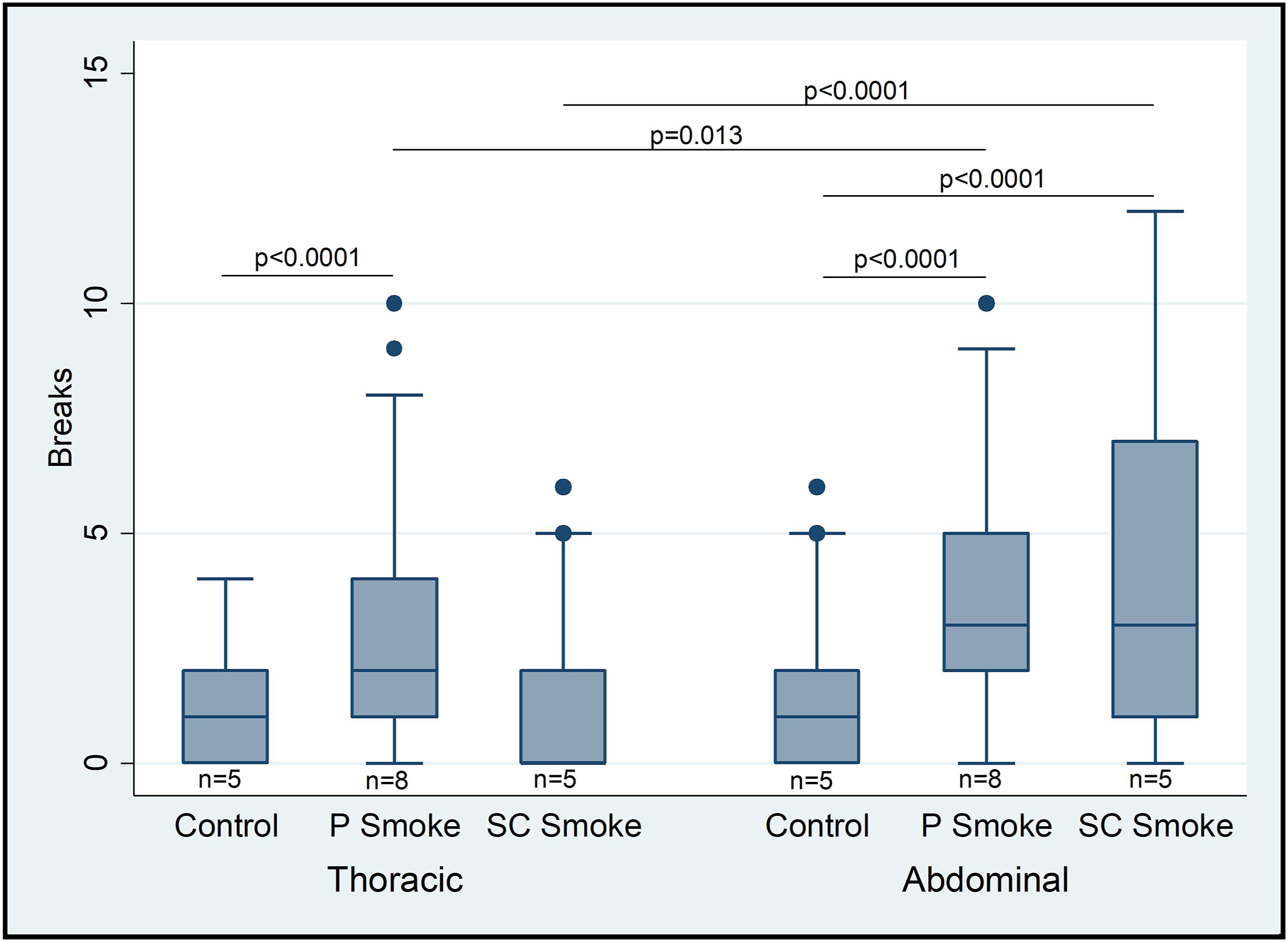
Elastic lamellar breaks occurred in the absence of inflammatory infiltrates. Immunohistochemical staining for myeloperoxidase, CD-3, and CD-20 were negative. Immunofluorescence microscopy also demonstrated no appreciable Mac-2 staining in the region of lamellar breaks (Figure 3). A. Breaks in elastic lamellae (white arrows) were assessed with Verhoeff-Van Gieson stain. 400X magnification, scale bar 50 µm. B. Immunofluorescent microscopy demonstrates no Mac-2 antigen expression in the vicinity of elastin breaks. 400X magnification, scale bar 50 µm.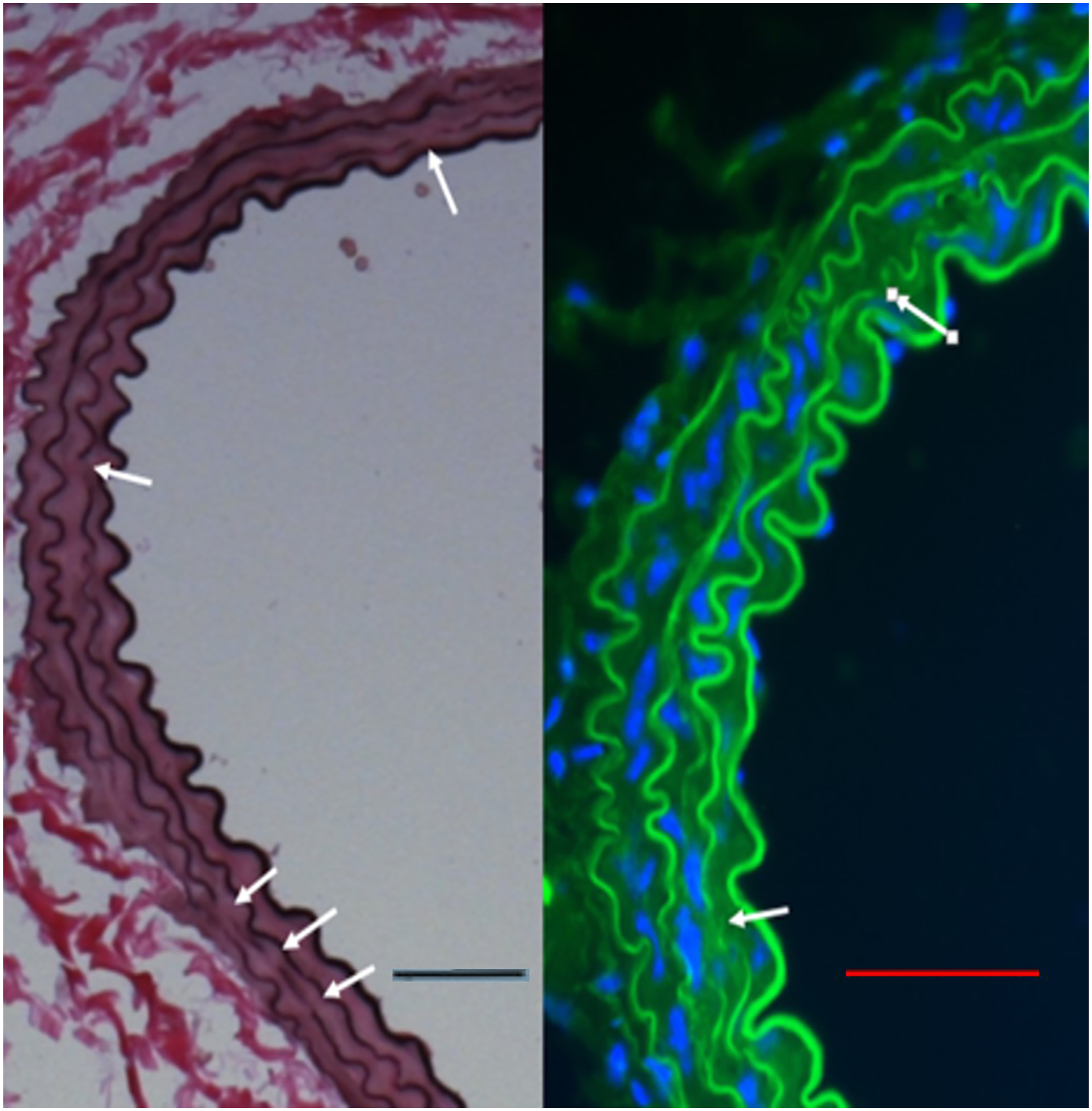
The paucity of continuous elastic lamellae in the infrarenal aorta did not allow for adequate comparison of elastin breaks at this location, with 71/240 sections demonstrating insufficient continuous elastic lamellae for analysis. Light microscopy with VVG staining of elastic lamellae as well as electron microscopy were used to demonstrate that elastic lamellae of the abdominal aorta are thinner, fewer in number and less often continuous than the elastic lamella of the thoracic aorta (Figures 4 and 5). Additionally, the abdominal aorta contains areas of extralamellar elastin which are not associated with organized lamellae (Figure 4). The mean number of elastic lamellae in the descending thoracic, suprarenal abdominal and infrarenal abdominal aorta were 4.7 ± .45, 3.1 ± .51, and 2.6 ± 1.0 lamellae, respectively (P < .01 for all groups) (Figure 5). Differences in elastic lamellae of thoracic and abdominal aorta. Lower panel: Light and electron microscopy of thoracic aorta (A and C) demonstrate thicker, more numerous, and more continuous elastic lamellae than the infrarenal abdominal aorta of the same animal (B and D). The abdominal aorta also demonstrates regions of extralamellar elastin (arrows in D). The boxed areas in A and B (scale bar, 100 µm) are shown as electron micrographs in C and D (scale bar, 10 µm). The descending thoracic, suprarenal (SR) abdominal and infrarenal (IR) abdominal aorta demonstrate a progressive decrease in the number of elastic lamellae (P <.01 for all groups).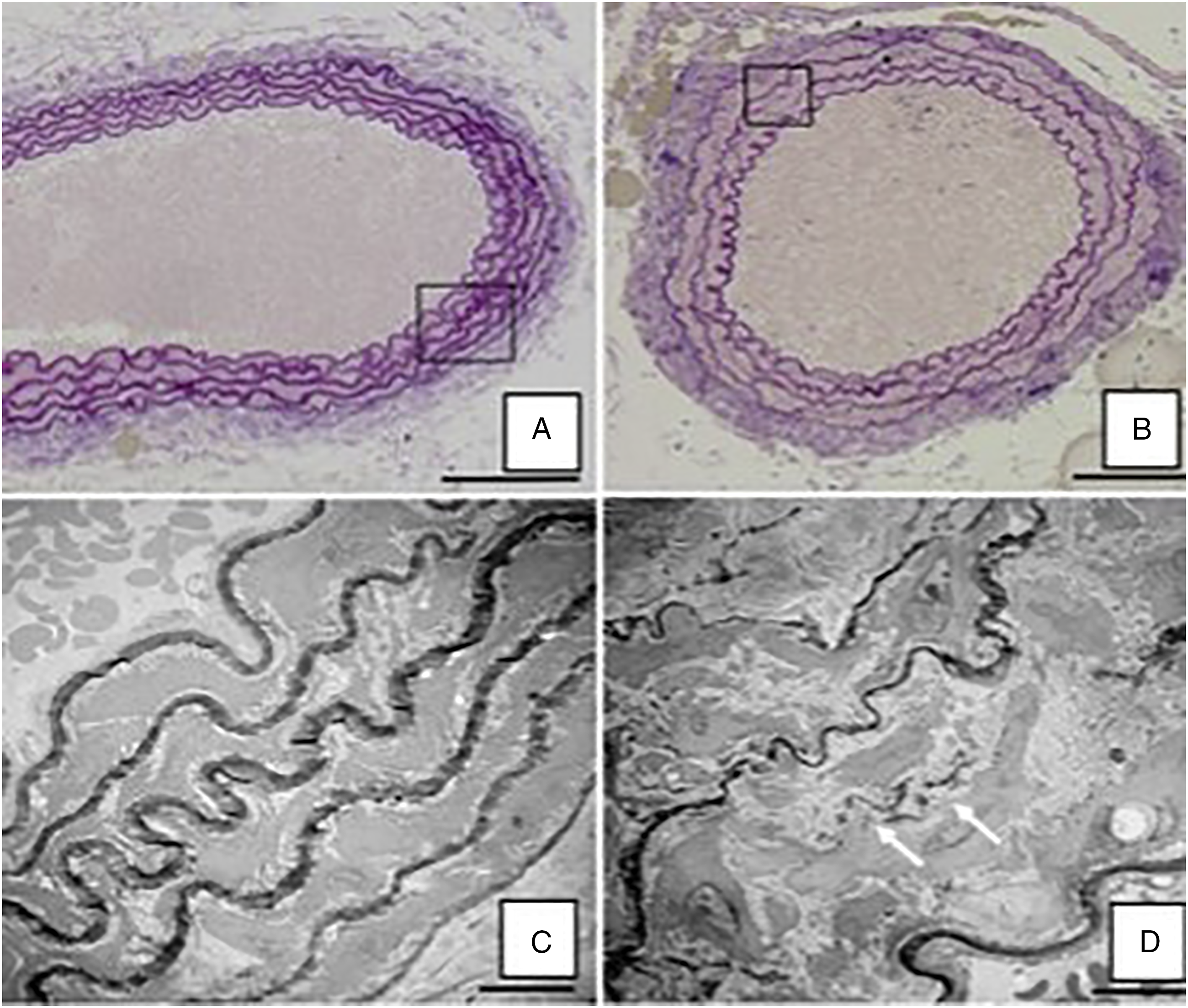

Tobacco Smoke Exposure Causes Downregulation of α-smooth Muscle Actin Expression in Vascular Smooth Muscle Cells In Vivo
Four of the 13 tobacco exposed mice demonstrated multiple areas with both Mac-2/α-SMA expression within the aortic media and in dissected areas of the aorta (Figure 6A and B). In severe lesions (Figure 6A and B), downregulation of α-SMA was observed in conjunction with increased staining for Mac-2 antigen, including some cells that expressed Mac-2 antigen without any discernable α-SMA expression. The control group did not show any evidence of Mac-2 expression, downregulation of α-SMA or any areas of aortic dissection (P = .28). Area of aortic injury in an STS-exposed mouse (thick arrow, A and B) demonstrating aortic dissection (*), marked expression of Mac-2 (green in A) and minimal α-SMA expression (red in B). The adjacent area of normal aorta (thin arrow, in A and B) shows normal α-SMA expression and no Mac-2 expression. 400X magnification (A and B). Vehicle-exposed mice did not express Mac-2 (C) and demonstrated normal α-SMA expression (D). Decreased α-SMA expression was seen in 4/13 STS-exposed animals and 0/5 vehicle-exposed animals (P = .28). Scale bars 50 µm.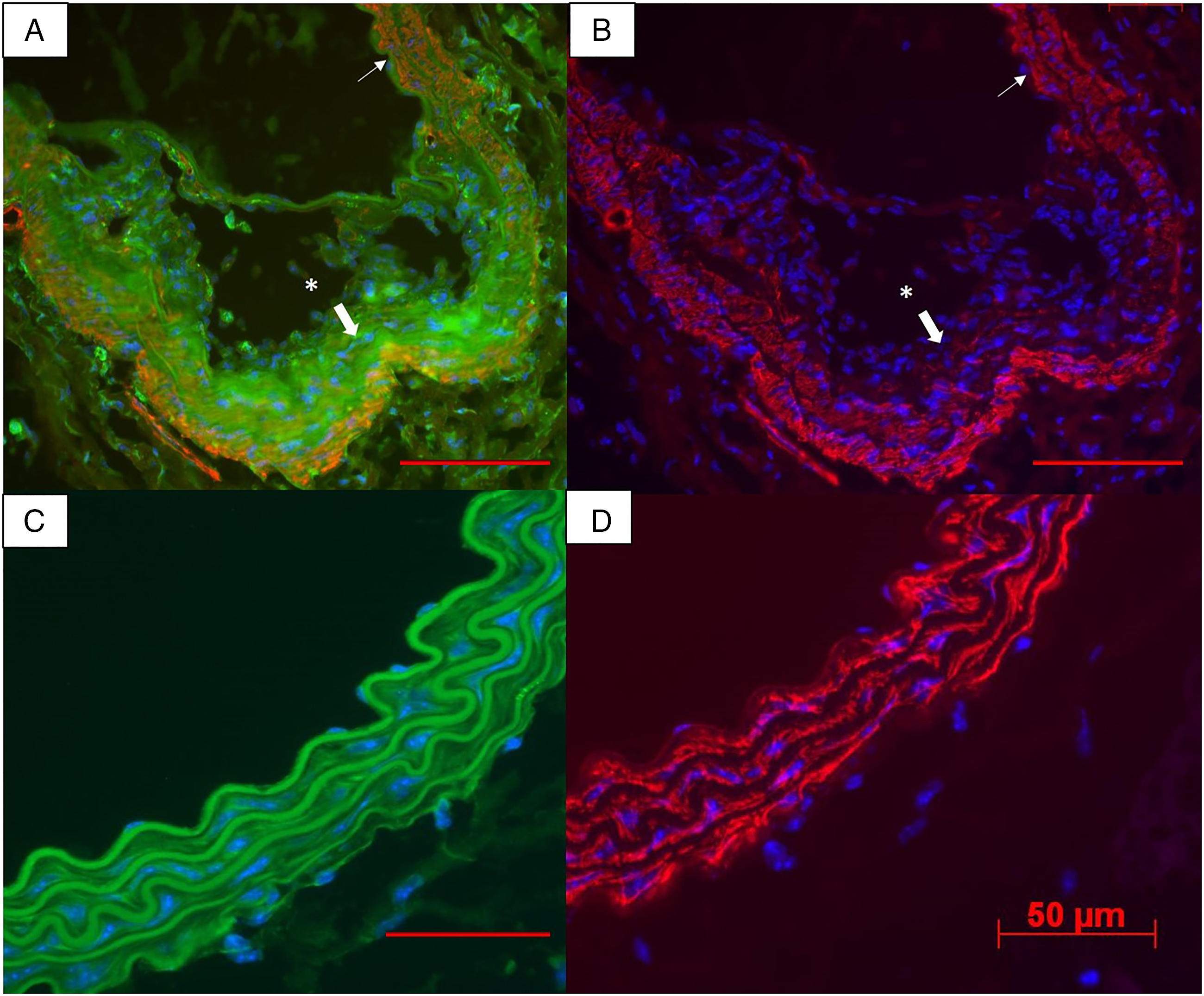
Tobacco Smoke Exposure Downregulates Expression of α-SMA in Cultured HASMC
Cultured HASMCs grown on media containing STS were compared with cells grown without addition of STS. Immunofluorescence microscopy demonstrates the loss of intracellular SMA filaments in STS-treated cells compared to controls (Figure 7A and B). The loss of α-SMA was verified using Western blot analysis (Figure 7C) Cultured human aortic smooth muscle cells demonstrate normal α-SMA expression in standard media (A) and downregulation of α-SMA after 4 days in media containing solubilized tobacco smoke (B). (C) Western blot analysis (left) of HASMC confirms loss of α-SMA expression in cells exposed to STS-containing media compared to cells in control media. Ponceau slide (right) showing equal loading of samples.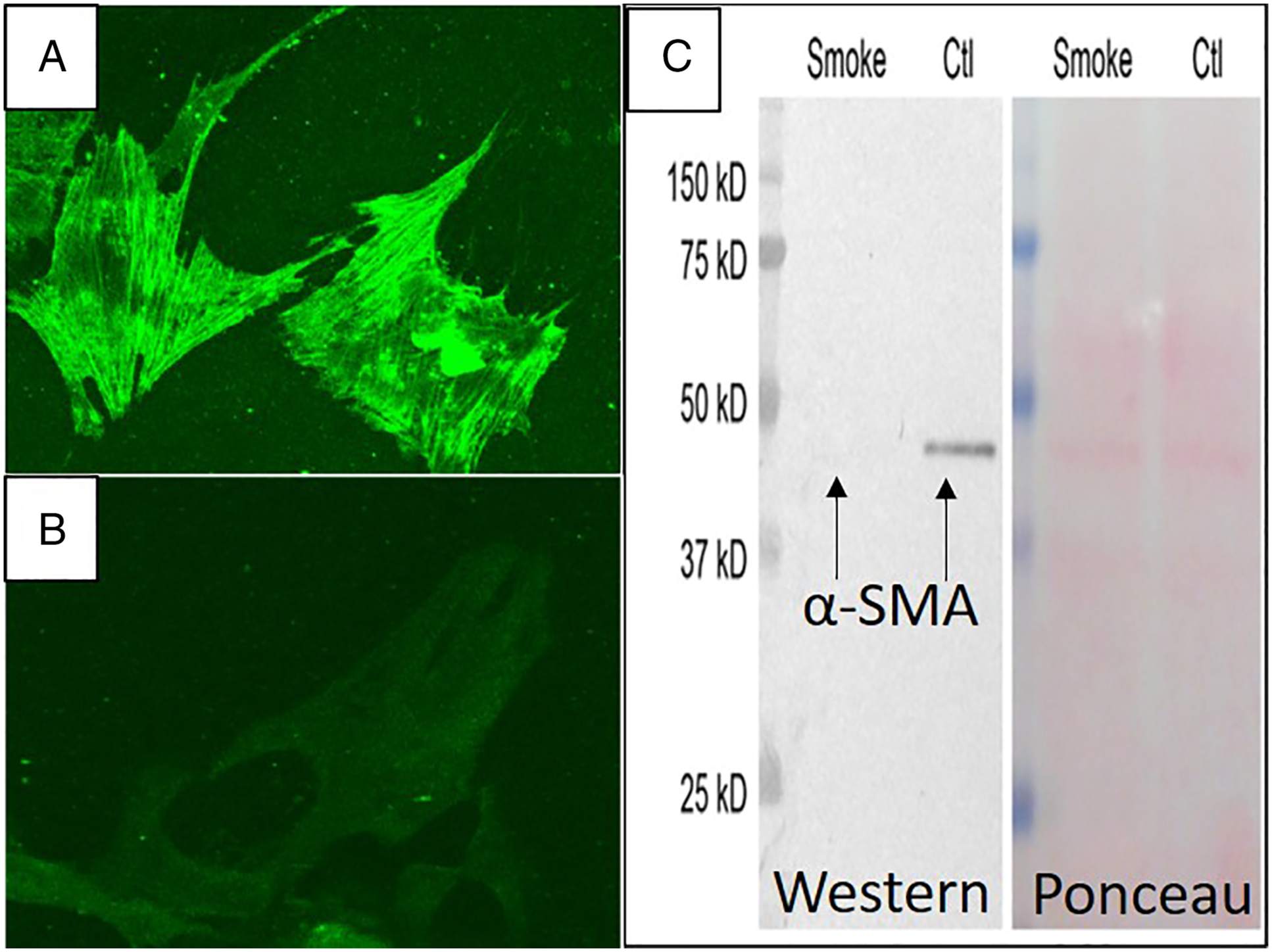
Discussion
Delivery of STS to mice via osmotic minipumps and the addition of STS to differentiated HASMC culture may be an effective model for investigating early tobacco smoke–mediated aortic damage. Tobacco smoke is thought to contribute to aortic aneurysm formation through inflammatory pathways; however, it is unknown whether structural changes or changes to aortic vascular smooth muscle cells (VSMCs) precede the onset of inflammation. It is also unknown why smoking is more strongly associated with abdominal aortic aneurysms than with thoracic aortic aneurysms.1,10-13 To address these gaps in knowledge, we developed a novel model of tobacco-induced aortic injury by utilizing STS in an in vivo model. The model was designed to simulate moderate tobacco smoke exposure in humans. Unlike the infusion of nicotine solution, 8 the wide absorbance spectra of STS demonstrates that STS contains multiple tobacco smoke components in addition to nicotine, and likely resembles tobacco smoke exposure more closely than the administration of nicotine solution alone. The use of a continuous smoke infusion, rather than intermittent exposure to inhaled tobacco, provides a simple way to provide prolonged, daily tobacco smoke exposure and mimics the smoking behavior of human smokers, who adjust smoking frequency to maintain a constant level of plasma nicotine.14,15 The concentration of nicotine in the STS was quantified so that physiologically relevant doses of tobacco smoke corresponding to roughly 5–11 cigarettes/day in a 70 kg human were used. Finally, the use of wild-type mice allowed us to discern the effects of tobacco smoke without confounding genetic factors. The surgical components of the model are straightforward and safe, with all mice surviving until the end of the study.
Although fragmentation of aortic elastic lamellae is a hallmark of aortic aneurysms, 16 it is unknown whether elastin loss precedes aneurysm formation or reflects end-stage disease processes. 17 Our findings suggest that aortic elastin damage can result from tobacco smoke exposure in wild-type mice and precedes aneurysm formation and inflammatory infiltration of the aorta. These findings suggest that initial tobacco-induced damage of aortic elastic lamellae may not be mediated through inflammatory pathways and may not depend on the presences of proteases produced by macrophages. These findings are consistent with those of Jin et al, who demonstrated that inhaled tobacco smoke exposure combined with elastase perfusion exacerbates elastic lamellar injury and aneurysm formation, and that this effect is not inhibited by protease inhibition or protease deficiency. 6 Our model also investigated 2 routes of STS delivery, subcutaneous and peritoneal. Both the subcutaneous and peritoneal routes of STS delivery were effective in producing elastin breaks within the abdominal aorta, but only the peritoneal route of STS delivery was associated with increased elastin breaks within the thoracic aorta, suggesting that the peritoneal route of delivery may be more effective producing elastin breaks. Future studies will assess blood nicotine concentration in order to better delineate the differences between peritoneal and subcutaneous STS delivery and will utilize mice of a single-age group in order to reduce potential age-related differences in response to tobacco smoke exposure.
Our study also highlights differences in elastic lamellae within different aortic regions and the difficulty of quantifying elastin breaks within the infrarenal aorta. It has been well documented that the human thoracic aorta consists of more numerous and thicker elastic lamellae than the abdominal aorta.18,19 The mean number of elastic lamellae in the ascending thoracic, descending thoracic, suprarenal abdominal, and infrarenal abdominal aorta of humans is 80, 57, 39, and 32 lamella, respectively. 20 Our study demonstrated a similar decrease in the number and thickness of elastic lamellae in the aorta of mice (Figures 4 and 5). Tobacco smoke–induced elastin breaks were most easily quantified in the thoracic and suprarenal abdominal aorta, where the large majority of the elastin is in the lamellar configuration. In our study, the frequency of elastin breaks caused by tobacco smoke exposure was greater in the suprarenal abdominal aorta compared to the thoracic aorta. It is possible that the thicker more numerous elastic lamellae of the thoracic aorta are more resistant to tobacco smoke–mediated damage compared to the thinner elastic lamellae of the abdominal aorta, which may explain why smoking is more strongly associated with abdominal rather than thoracic aortic aneurysms.
The paucity of continuous elastic lamellae in the infrarenal abdominal aorta made it difficult to quantify the number of elastin breaks in the infrarenal abdominal aorta using our pre-defined criteria. It is possible that the decreased amount of lamellar elastin in the abdominal aorta predisposes this region to the damaging effects of tobacco smoke, but this effect cannot be quantified using elastic fiber breaks. It is possible to apply existing grading scales for elastin damage to the infrarenal aorta,21,22 but the presence of non-continuous elastic lamellae and extralamellar elastin may result in overestimation of elastin damage with such methods. In future studies, the effect of tobacco smoke on the elastin content of the infrarenal aorta may be more objectively assessed through the use of imaging software to measure elastin content relative to aortic cross-sectional 20 area or by measuring total aortic elastin, similar to studies of human aorta. 23
While quantification of elastin breaks or elastin damage may be subject to interobserver variation, our study applied fairly rigorous, pre-defined criteria to identify elastin breaks and used 2 separate, blinded observers to determine the number of breaks in each section. Furthermore, our study defined and compared the same aortic segments (descending thoracic, suprarenal abdominal, and infrarenal abdominal) between animals. The uses of defined criteria for elastin breaks minimizes the subjectivity in counting elastin breaks and helps distinguish true breaks in elastic lamellae from areas of sparse elastic content and areas of artifact. Comparing the same aortic segment between animals controls for the fact that the baseline content, structure, and continuity of elastic lamellae differs between the thoracic and abdominal aortic segments.
Vascular smooth muscle cells loss is another hallmark of human aortic aneurysm and VSMC death has been implicated in animal models of aortic aneurysm as well.24-26 Our study suggests that tobacco smoke can cause decreased expression of α-SMA within VSMC. Previous studies have demonstrated a similar VSMC phenotypic switch in angiotensin II-induced aortic aneurysms, cerebral aneurysms and in response to hyperlipidemia.27-29 Our study is the first to describe downregulation of α-SMA in VSMC in response to tobacco smoke exposure. Within areas of microdissection, Mac-2 staining was observed in addition to α-SMA downregulation. It is unclear if the Mac-2 staining occurs due to a phenotype change in VSMC or represents an influx of inflammatory cells in these structurally damaged regions. The exposure of cultured HASMC to smoke solution verified the loss of α-SMA, but failed to verify expression of Mac-2 antigen. More complex lineage tracing animal models will be needed to determine if the Mac-2 expressing cells in dissected areas of the aorta are transformed VSMCs or infiltrating inflammatory cells. Additionally, larger sample sizes or doses of STS may be needed to demonstrate a statistically significant difference in number of mice with areas of α-SMA downregulation and Mac-2 expression across exposure groups.
Our animal model did not produce a gross aneurysm. We intentionally used a relatively low amount of tobacco smoke exposure and wild-type mice in order to determine the first discernable arterial changes in response to tobacco smoke exposure, prior to aneurysm formation. Future experiments using higher doses and duration of STS exposure, hyperlipidemic mice, genetically susceptible mice and additional insults, such as angiotensin II infusion, may produce more exaggerated aortic injury and aneurysm formation.
Conclusion
Delivery of STS to mice via implanted osmotic minipumps may provide a model for studying early tobacco smoke–mediated aortic changes that occur prior manifestation of aortic disease. The initial aortic response to tobacco smoke exposure appears to be elastic lamellar damage and downregulation of α-SMA in aortic VSMCs. Further characterization of early tobacco smoke–induced aortic injury may identify biomarkers of subclinical aortic injury and may provide therapeutic targets to prevent progression to aortic aneurysm formation.
Footnotes
Acknowledgments
We thank Marjorie Grafe MD, PhD and Xiao Jing Nie, MS for their assistance with tissue processing and slide preparation
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the Medical Research Foundation of Oregon (AA) and the Knight Cardiovascular Institute (AA).