
Abstract
In sexually colour dimorphic bird species, males can exhibit phenotypic variation, with males breeding in either dull female-like plumage or brightly coloured plumage. Two contradictory hypotheses predict that the male phenotype variation can influence the female investment in a given breeding attempt. Whereas females usually prefer males in bright coloured plumage, the “differential allocation hypothesis” predicts that females should invest more in their reproductive output when mating with them; while the “compensatory investment hypothesis” predicts that females should invest more when mating with non-preferred males. To test those predictions, we analysed reproductive data for two consecutive breeding seasons (2018–2019 and 2019–2020) of Lined Seedeaters Sporophila lineola. S. lineola is a socially monogamous songbird species in which males exhibit two breeding phenotypes, a black-and-white plumage being the most common, and a less common female-like brownish plumage. Our findings show that females mated with brownish males have a higher reproductive investment (i.e., egg volume) than those mated with black-and-white males. Despite the lower investment of females in egg volume, our results showed that black-and-white males have a higher nestling survival, producing more hatchings and fledglings per season than brownish males, which could suggest a higher fitness. Our findings indicate that investment allocation on the eggs and offspring survival rates can be attributed to the plumage colour of males.
Introduction
Males of sexually colour dimorphic bird species can exhibit high phenotypic variation in plumage aspect (e.g., dull and brightly coloured) that can be associated with their age, social status, and quality.1–3 In species with delayed plumage maturation, males are dull and/or female-like in appearance in their first breeding season (s), only exhibiting the adult plumage in their second or even subsequent moult cycles. These males are often socially subordinate to brightly coloured males.2,4 Moreover, adult plumaged males are usually more attractive to females, obtaining higher quality mates, higher nesting success 1 , and more extra-pair young than dull males 5 .
This intraspecific male phenotype variation can also influence the female breeding investment in species with biparental care, with females adjusting their reproductive investment according to the attractiveness of their mates. 6 The “differential allocation hypothesis” predicts that females invest more when paired with high-quality males since they provide better resources and/or genes.6–8 On the other hand, the “compensatory investment hypothesis” predicts that females paired with low-quality males increase their investment to compensate for their lowered offspring viability.9,10 The differential allocation hypothesis is most likely to occur in a mating system where females should preferentially invest in the highest quality offspring. On the other hand, the compensatory investment is the case in mating systems where even low-quality offspring will reproduce successfully. 10
In Lined Seedeater Sporophila lineola, a small songbird with marked sexual dimorphism and delayed plumage maturation, breeding males exhibit high phenotypic variation. Males breed at the end of the first moult cycle in the female-like dull brownish plumage, only acquiring the characteristic adult black-and-white plumage after the first breeding season. Some individual variations in this process, however, cannot be disregarded. 11 Males in the adult plumage are black above and white below, with conspicuous white patches on the head, wings, and rump. 12 Thus, we observe two distinct phenotypes of breeding males, the most common ornamented black-and-white males and the less common dull brownish males. 11 Differences in male quality and female preference between the two distinct phenotypes are unknown for Lined Seedeaters.
In sexually colour dimorphic birds, plumage ornamentation is often assumed to be a sexually selected trait,13,14 with more ornamented males being preferred by females.5,15,16 Moreover, the expression of ornaments depends on the quality of the individual.17,18 In this study, we investigated the influence of male plumage colouration on male quality and female breeding investment in Lined Seedeaters. Once mate phenotype can influence female investment, we predict that females invest differentially in the current brood according to male plumage. Assuming that the black-and-white plumage is a sexually selected trait and considering that it is the most common phenotype in the study species, we expected that the black-and-white males have higher nesting success and higher number of hatchlings and fledglings per nest.
Materials and methods
Study site
We conducted a field study at the Campus Florestal of the Federal University of Viçosa (19°52’51’’S and 44°24’49’’W), south-eastern Brazil. Lined Seedeaters inhabit pastures dominated by African grasses (Panicum and Brachiaria), crops, and gardens around buildings and ponds. 19
Studied species
The Lined Seedeater is a socially monogamous intra-tropical migratory songbird which occurs in open habitats across South America. 12 Members of this species arrive in the study area in late November, and the laying dates span from early December to late April. 19 Males defend small territories by singing from sunrise to sunset. 20 The distinction between females and similar-looking brownish males is possible based on their behaviour (males sing, while females do not) or in the presence of brood patch. 11
Only the females build the nests and incubate the eggs, while both sexes feed the nestlings.19,21,22 The typical clutch size is two or three eggs and individuals can make up to six attempts per season (unpublished data). The mean incubation period is of 12.7 days and average nestling period is of 11.2 days. 19 In the study site, Lined Seedeaters exhibited a lower nesting success (less than 40%) and predation was the main cause of nesting failure (more than 50%). 19
Field methods
We captured individuals with mist-nets in feeding sites and breeding territories, during two consecutive breeding seasons, from November to April (2018–2020). Individuals were ringed with a unique combination of colour-rings and a numbered metallic ring. We measured the length of the right wing (“Wmax”) using a ruler (to 0.5 mm accuracy) for each individual captured. 23 We used wing length as representative of the structural body size.23,24 Wing length is routinely used as a measure of body size in passerines and is particularly important for migratory birds as a flight-related trait. Moreover, the reproducibility of wing length was generally better than for other linear measures. 24
To find nests, we observed the reproductive activity of adults, such as the transport of material for nest building or food for nestlings. We registered the clutch size and the number of hatchings and fledglings for each nest monitored whenever possible. We took the length and width of each egg using a calliper (to 0.1 mm accuracy) and calculated the egg volume (mm3) using the formula 0.51 x length x width2. 25 We then calculated the mean egg volumes per clutch. We monitored nests systematically at 1–2 days intervals. Nests were monitored from their encounter until success or failure. We also estimated for each nest the egg survival (the proportion of eggs laid that hatched) and the nestling survival (the proportion of nestlings that fledged). Nests were considered successful when at least one nestling fledged.
Statistical analysis
We used the R software and the R packages “lme4”,26,27 “lmerTest”, 28 “RVAideMemoire”, 29 for the statistical analyses, and “ggplot2” to make the visual data representation. 30 We used linear mixed-effects models to test: (i) whether male plumage type correlates with body size using wing length as response and male plumage as predictor; and (ii) whether male plumage type or body size explain variation in female breeding investment, using mean egg volume per clutch as response variable and male plumage or body size as fixed effects. We use residual plots to check the assumptions to validate these models.
We used generalized linear mixed models (glmm) to test: whether male plumage colour or body size explains variation in (iii) female breeding investment in clutch size; and (iv) the reproductive output as measured by number of successful breeding attempts, number of hatchlings and fledglings per season, (Poisson error distribution) egg survival, nestling survival, and nesting success (Binomial error distribution). Since the same pair can have multiple breeding attempts per season, when necessary, we include male ID and female ID as random effects in the models.
Results
Outcome of the 252 nests of Lined Seedeaters Sporophila lineola monitored in south-eastern Brazil during two breeding seasons per male plumage colouration.

Results of the Linear and Generalized Linear Mixed Model of breeding variables in relation to male’s plumage colour (brownish or black-and-white) and male’s wing length (used as a proxy of body size), the values showed have brownish plumage colour as the reference level. Data from 252 nests monitored during two breeding seasons (2018

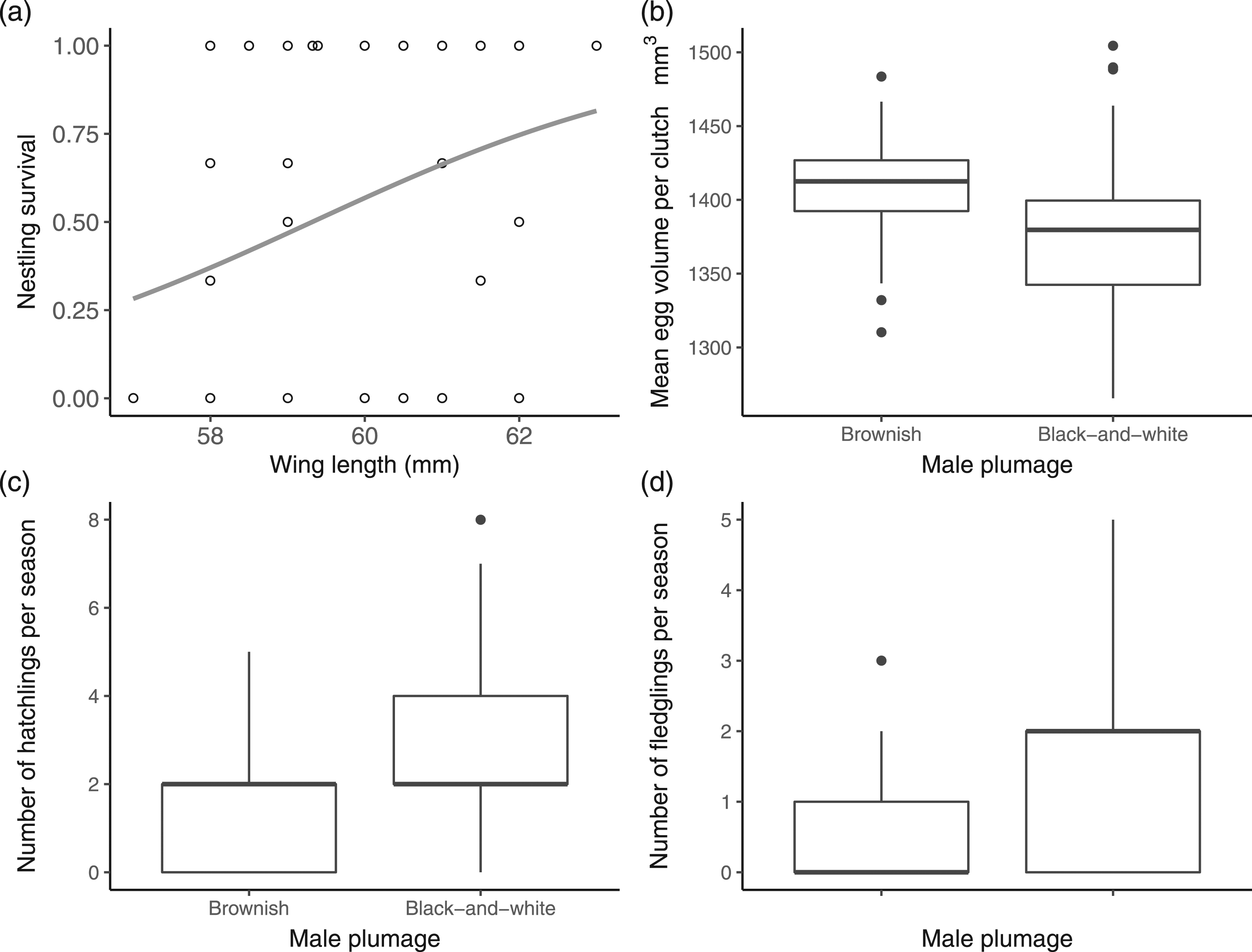
(a) Nestling survival in relation to wing length with a fitted binomial curve and 95% confidence interval (n= 71 nests of 41 breeding males). (b) Plot of predicted values of the model build using mean egg volume per clutch in relation to male plumage. (c) Number of hatchlings per season in relation to male plumage from 68 ringed breeding males. (d) Number of fledglings per season in relation to male plumage from 68 ringed breeding males. Data from nests monitored during two breeding seasons (2018-2020) of the Lined Seedeater Sporophila lineola in south-eastern Brazil.
Females paired with brownish males laid clutches with higher volume per egg (1407.7 ± 92.7) than females paired with black-and-white males (1373.8 ± 115.6) (Table 2). However, the clutch size was similar between females paired with males of different plumages. Egg survival, nesting success, and the number of successful breeding attempts per season were also independent of males’ plumage (Figure 1). Nonetheless, black-and-white males had a significantly higher number of hatchlings and fledglings per season when compared to brownish males (Table 2) (Figure 1).
Discussion
In this study, we used field nesting monitoring data to investigate how phenotypic variation in the plumage colouration of breeding males correlates with their body size and influence female investment and nesting success in Lined Seedeaters. Males in different plumage colouration showed similar body sizes estimated by wing length. Contrary to studies that showed that the wing length increases after the first complete moult in many passerines,31–33 brownish males on their first moult cycle showed similar wing length to males in black-and-white plumage that had already undergone a complete moult. However, independent of male plumage, the wing length influenced nestling survival. Males with longer wings yielded a higher nestling survival on their nests. Wing length may impact the male’s ability to acquire and defend higher quality territories which may influence the nestling survival.
Females varied their investment according to the male's phenotype. Clutch size was similar between nests of males with different phenotypes, but females paired with brownish males laid larger eggs than females paired with black-and-white males. We assume that black-and-white male plumage is preferred by females, as it is a product of mate choice and is the more common phenotype in the population. Females of Lined Seedeaters invested more in egg volume if paired with not preferred males, which is in line with the “compensatory investment hypothesis”. 10 Similar results were reported for Collared Flycatchers (Ficedula albicollis), as females paired with younger males showed higher yolk testosterone concentration in their eggs compared to the eggs laid by females paired with older males. 34 Likewise, Barn Swallow (Hirundo rustica) females mated with unattractive males laid eggs with higher lutein concentration than eggs laid by females mated with attractive males. 35
Investing more in breeding when paired with less attractive males may indicate an adaptive maternal strategy to enhance offspring viability. 35 A review study reported a positive relationship between egg size and offspring fitness, hatching mass, nestling growth, and nestling survival. 36 In other words, larger eggs produce higher-quality nestlings. Due to the higher egg volume, nests of brownish males would be expected to have higher nestling survival rates and, therefore, be more successful than those of black-and-white males. However, our results showed the opposite: brownish males achieve low nestling survival despite higher female investment compared to black-and-white males. If we consider nestling survival rates as a proxy for the ability to raise young, lower nestling survival in nests attended by brownish males may suggest that they are poor resources provider.
Black-and-white males are older than brownish males. 11 Thus, black-and-whites could perform better due to previous breeding experience providing their offspring with better resources. Studies showed that more experienced birds are more efficient at feeding and caring for their nestlings and often obtain higher quality territories, which makes it possible for them to deliver a high energy rate per visit to the nest.37,38 Moreover, breeding in underprivileged circumstances might benefit from a reduction in egg investment and making more attempts per breeding season. 39 This might explain why females of Lined Seedeaters invest less when paired with black-and-white males, which may contribute more to nestling quality than brownish males.
Predation is the major cause of offspring mortality in our studied population, affecting 58.5% and 52% of the nests during the two breeding seasons monitored. 19 Thus, choosing a safe nest site to raise young is essential because it can influence the probability of nest predation. Lined Seedeater males are responsible for suggesting nest site to females. 40 Hence, due to their inexperience in choosing better nesting sites, brownish males may offer lower reproductive output. However, still little is known about nest predators and which factors influence nest predation in the study species. In brief, older males (black-and-white) may choose better nesting sites, allowing them to raise young successfully.
In conclusion, it is remarkable that males in female-like plumage can attract mates and breed as satisfactorily as adult males in black-and-white plumage, reaching similar nesting success. Female-like plumage is usually a signal of subordinate status and a low-cost strategy to breed adopted by less experienced breeding males.2,4,5,41 Our study suggests that females are willing to invest more in egg volume if paired with brownish males. One possible explanation is that this relation may reflect an adaptive maternal response to compensate for the likely lower quality of paternal care provided by the younger and inexperienced brownish males due to their little breeding experience. The impact of a larger egg volume in the long-term survival of Lined Seedeater fledglings are not yet understood. Higher reproductive outputs exhibited by black-and-white males can be linked to the better parental care provided to offspring. However, whether black-and-white males are better parents than brownish males still needs to be tested.
Footnotes
Acknowledgements
We thank the Lined Seedeater Project team for their help during data collection. UFV supported our fieldwork.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Financial support has been provided by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior- Brasil (CAPES) [Finance Code 001 to DFF], Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) [309660/2017-3 and 316960/2021-7 to LEL].
Ethical and legal aspects
UFV allowed us to conduct this study in the area under their care. Permits were granted by ICMBio (61078-1) and CEUA-UFV (03/2018).