
Abstract
Interspecific hybridization is an effective approach for creating novel Rubus germplasms, but early hybrid authentication is difficult when parental genetic backgrounds are complex and ploidy levels differ. In this study, we evaluated the F1 progeny derived from a cross between the hexaploid primocane-fruiting blackberry cultivar ‘Freedom’ (female parent) and the diploid Rubus hirsutus male parents via seven SSR primers. To account for parentage effects, hybrid identification was summarized by cross combination: Freedom × R-1/2 (n = 5), Freedom × R-3 (n = 35), and Freedom × R-4 (n = 41). Across all crosses, 62 seedlings (3 + 27 + 32) were authenticated as true hybrids (76.5%). Flow cytometry profiles of two vigorous F1 individuals revealed that the nuclear DNA fluorescence peak positions were intermediate between those of the diploid male parent and the hexaploid ‘Freedom’, which was consistent with a tetraploid level. From the authenticated hybrids, 26 vigorous seedlings were selected for phenotypic and physiological evaluation. Considerable variation was observed among the selected hybrids in terms of plant height, SPAD, total nitrogen content, and antioxidant-related traits (CAT, SOD, GSH, VC, TP, and TF). SSR-based diversity analysis revealed moderate polymorphism (18 polymorphic bands; expected heterozygosity 0.27–0.64, mean 0.44; Nei's genetic diversity 0.26–0.64). These results provide SSR marker references for early hybrid authentication in wide Rubus crosses and support subsequent selection of promising interspecific hybrid germplasms.
Introduction
Rubus L. is a large genus in the Rosaceae family, with more than 700 known species. 1 It is distributed on five continents around the world, most of which are distributed in the temperate zone of the Northern Hemisphere, North America and East Asia, and few species are distributed in the tropics and southern hemispheres. 2 The fruit of most plants in the genus Rubus are edible and have important economic, nutritional and medicinal value. 3 Among them, blackberry and raspberry are important fruit crops with delicious fruit tastes, unique flavors and high nutritional value. They can be eaten fresh or processed into juice, jam, ice cream or flavored tea, and are highly loved by consumers. 4
According to Wang's (2009) research, the adaptability of blackberry at the introduction site is better than that of raspberry. The main objectives of the development of new cultivars of blackberry and raspberry are high yield, good adaptability, thornless or thornfree, upright growth and excellent fruit quality. 5 The primocane-fruiting blackberry cultivar ‘Freedom’ has the advantages of early fruiting and a long harvest season, but fruit quality and disease resistance still need improvement, especially at the late fruiting stage. There are more than 200 species of Rubus widely distributed in China, among which some have high utilization value and development prospects. Paying attention to the introduction and domestication of excellent wild species can provide valuable resources for breeding excellent cultivars of raspberry and blackberry in the future.1,2 Rubus hirsutus Thunb. is a wild species native to China with attractive red fruits and a unique aroma. 6 Compared with traditional cultivated cultivars, wild varieties have higher secondary metabolite levels, antioxidant activity and nutritional value. 7 In recent years, raspberry and blackberry improvement programs have focused on disease and pest resistance, tolerance to hot and humid climates, and enhanced fruit quality. Common strategies for genetic enhancement include interspecific hybridization, ploidy manipulation, and mutagenesis, with the former two being the most prevalent (Wang, 2009). Hybridization has great advantages in operational costs, offspring segregation and trait combination, and the application of molecular marker technology has been found to be important and effective in the development of new cultivars. 8
Morphological identification is a traditional method for hybrid identification, its identification cycle is long, and the results are easily affected by environmental conditions, resulting in false positive results. 9 SSRs are “simple sequence repeats” and are also known as microsatellite DNA. It is usually a DNA sequence composed of 1 to 6 bases as motifs connected in tandem after multiple repetitions. 10 Zhang et al. 11 used SSR primers from Rubus to molecularly identify ‘Ningzhi 2’, the first blackberry hybrid cultivar bred in China. Among them, 70 pairs of SSR primers could amplify clear bands, and 7 pairs of primers had obvious amplification band differences for the tested materials, with a polymorphism percentage of 10%, but the amplified loci did not show co-dominance, 11 which indicated the heterozygosity and complexity of blackberry genetic background. Sharma et al. 12 obtained the genetic information of R. ellipticus through transcriptome sequencing, and developed a batch of SSR markers that can be used for genetic research and breeding. These SSR resources have also been applied to trait-oriented studies in Rubus, with berry pigmentation being a representative example. A nonsense mutation inside the anthocyanidin synthase gene controls pigmentation in yellow raspberry. 13 Furthermore, Lebedev et al. 14 assessed the genetic diversity in differently colored raspberry cultivars via SSR markers located in flavonoid biosynthesis genes. These SSR markers exhibited high polymorphism and transferability, effectively identifying allelic variations related to color traits. Overall, these examples highlight the broad utility of SSR markers in Rubus. SSR markers have been widely applied for cultivar identification and hybrid authentication in Rubus.15,16
Our team has obtained a number of hybrid offspring of double-cropping blackberry ‘Freedom’ and R. hirsutus Thunb. Understanding and mastering the genetic basis of excellent traits is the final way of heterosis combination selection, and it is also the premise and foundation of new cultivar development. To reduce breeding costs and shorten the breeding time, and to provide theoretical and technical support for germplasm innovation, utilization, and popularization of Rubus wild resources. In this study, the F1 generation hybrid plants obtained from double-cropping blackberry ‘Freedom’ and R. hirsutus were taken as the research material, and the research objectives were as follows: (1) analyze the variation traits of F1 generation hybrid plants; (2) identify the authenticity of the hybrids and explore the genetic relationship between hybrid offspring and parents through SSR molecular markers; (3) further study the genetic diversity level of hybrid offspring, to provide reference for breeding of excellent cultivars of raspberry and blackberry.
Materials and methods
Plant materials
The cross experiment and sample collection were carried out in Nanjing Lishui Research Base (31°35’ N, 119°09’ E) of Institute of Botany, Jiangsu Province and Chinese Academy of Sciences. The double-cropping blackberry ‘Freedom’ (hexaploid) was used as female parent, and four provenances of R. hirsutus (diploid) from different regions (Lishui, with red fruit; Nanjing Botanical Garden Mem. Sun Yat-Sen, one with red fruit, one with yellow fruit; Laoyaba, with red fruit) were used as male parent. Notably, the yellow-fruited R. hirsutus from Nanjing Botanical Garden Mem. Sun Yat-Sen is a bud sport of the red-fruited R. hirsutus and has low pollen production. Therefore, we mixed the pollen of these two fruited types for hybridization with the female parent, resulting in a total of three hybrid combinations in this study. The contemporary fruits were harvested, and the seeds were washed with pectinase, treated with hypochlorous acid for 12 h, and sown in early March of the following year. A total of 81 hybrid plants of F1 generation were obtained. After 6 months of age, 2–3 fresh leaves were collected randomly and stored in ice boxes and brought back to the −40°C laboratory.
DNA extraction and SSR primer design
A rapid plant genomic DNA extraction kit (BioTeke company, centrifuge column type) was used to extract DNA from leaves according to the instruction method, and 1% agarose gel electrophoresis and full-wavelength microplate reader (Thermo Scientific: Multiskan sky) were used to detect whether the DNA was successfully extracted, its purity and concentration. Then, DNA of the all samples to be detected was stored in a refrigerator at -20 °C for later use.
SSR primers were adopted from Zhang et al., 17 where primers were developed from the male parent transcriptome and screened/validated for interspecific hybrid identification (Table 1). Rh-11, Rh-20 and Rh-34 are polymorphic loci amplifying in both parents, whereas Rh-2, Rh-3, Rh-4 and Rh-16 act as paternal-diagnostic markers because the diagnostic band(s) are present in R. hirsutus but absent in ‘Freedom’, enabling direct authentication of hybrid seedlings. Moreover, because gel-based SSR profiling may occasionally yield non-amplification at single loci, the absence of a band in one primer was treated as missing or undetermined information for that locus rather than definitive evidence of non-hybridity. Hybrid authenticity was judged from the combined evidence of the seven SSR primer pairs, with priority given to the male-specific diagnostic band(s) produced by Rh-2/Rh-3/Rh-4/Rh-16; Rh-11/Rh-20/Rh-34 were used as supporting loci, and seedlings showing insufficient or inconsistent marker evidence were treated as undetermined and were not included in downstream diversity analyses.
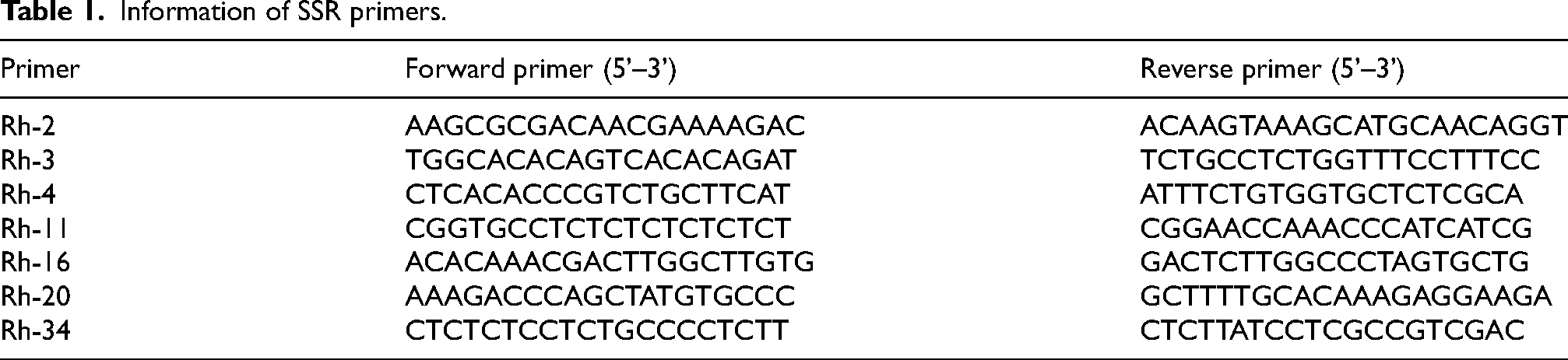
Information of SSR primers.
SSR-PCR reaction procedure
SSR-PCR amplification was performed on GE4852 T and XP cycle PCR instruments. The total volume of the amplification system was 10 μL, including 5 μL of 2 × Taq PCR Master Mix, 1 μL of forward primer (10 μM) and reverse primer (10 μM), 1 μL of template DNA and 2 μL of ddH2O. Reaction procedure for primer amplification: pre-denaturation at 95 °C for 2 min, denaturation at 94 °C for 40 s, annealing at 54 °C for 45 s, extension at 72 °C for 45 s, 30 cycles. The terminal was extended at 72 °C for 7 min, and the PCR amplification products were stored at 4 °C.
Polyacrylamide gel electrophoresis
Take 1 μL of the amplified product and add it to a 9% non-denaturing polyacrylamide gel for detection. Spaces on the left and right ends to avoid edge effects. A 50 bp marker was placed at each end. The electrophoresis buffer was 0.5× TBE. The electrophoresis was carried out at 120 V and 400 mA for 30 min, and then at 180 V and 400 mA for 1 h and 15 min. After electrophoresis, take out the gel, perforate and mark it with the tip of the gun, stain AgNO3 solution with silver for 10 min, then use the color developing solution to develop the color for 5 min, finally rinse it with distilled water twice, and place it on a light box to take pictures.
Flow cytometry
Flow cytometry was used to estimate the ploidy levels of the parents and selected F1 progenies. Young leaves from the male parent R. hirsutus, the female parent ‘Freedom’, and two vigorous F1 individuals were processed for nuclei preparation and DNA staining following a standard protocol, and nuclear DNA fluorescence was measured via a MoFlo XDP flow cytometer (Beckman Coulter, USA). The male parent R. hirsutus (2n = 2x = 14) was used as a control, and ploidy levels were determined by comparing the relative DNA fluorescence peak positions among samples.
Detection of phenotypic and physiological data
The following indicators were measured after the hybrid plants had grown for 6 months.
Plant height: A tape was used to measure the distance between the root neck of the plant and the top, where the top refers to the top of the main stem.
SPAD value and total nitrogen content: determined via a chlorophyll nutrient analyzer (Zhejiang Top Cloud-Agri. Technology Co., Ltd, TYS-4N).
The contents of catalase (CAT), superoxide dismutase (SOD), reduced glutathione (GSH), vitamin C (VC), total phenol (TP) and total flavonoid (TF) were detected using the relevant ELISA kit of Shanghai Yingxin Laboratory Equipment Co., Ltd, and finally by Infinite F50 enzyme labeling analyzer.
Data processing and analysis
Multiple allelic bands amplified by each primer pair were uniformly labeled across all samples. SSR banding profiles were scored manually from gel images: only clear and unambiguous bands were recorded, whereas faint, smeared, or ambiguous bands were treated as missing data. Band presence was coded as ‘1’ and absence as ‘0’, and a binary (0/1) SSR matrix was constructed for subsequent analyses. Genetic similarity was calculated in NTSYSpc v2.10e via the SIMQUAL module based on the binary matrix, with the Jaccard (J) similarity coefficient. Cluster analysis was then performed via the SAHN module with the UPGMA method, and the dendrogram was visualized via TREEPLOT. Genetic diversity parameters were calculated via POPGENE v1.32 based on the binary SSR dataset. Statistical analysis of phenotypic traits and physiological data was performed using Excel 2019. Differences among cross combinations were evaluated using the Kruskal–Wallis test (α = 0.05), treating each seedling as a biological replicate. Pearson correlation analysis and correlation heatmaps were generated using Origin 2022, and principal component analysis (PCA) was also conducted in Origin 2022.
Results
Hybrid identification
Hybrid authenticity of the F1 progeny derived from ‘Freedom’ (female parent) × R. hirsutus (male parent) was assessed using seven SSR primers (Rh-11, Rh-20, Rh-34, Rh-2, Rh-3, Rh-4 and Rh-16) reported in Zhang et al. 17 The male parents comprised four R. hirsutus genotypes from different origins: R-1 (red-fruited, Nanjing Botanical Garden Mem. Sun Yat-Sen), R-2 (yellow-fruited, Nanjing Botanical Garden Mem. Sun Yat-Sen), R-3 (Laoyaba), and R-4 (Lishui). To address parentage effects, hybrid identification results were summarized by cross combination: Freedom × R-1/2 (n = 5), Freedom × R-3 (n = 35), and Freedom × R-4 (n = 41) (Table 2).
Identification of hybrid F1 plants using SSR primers.
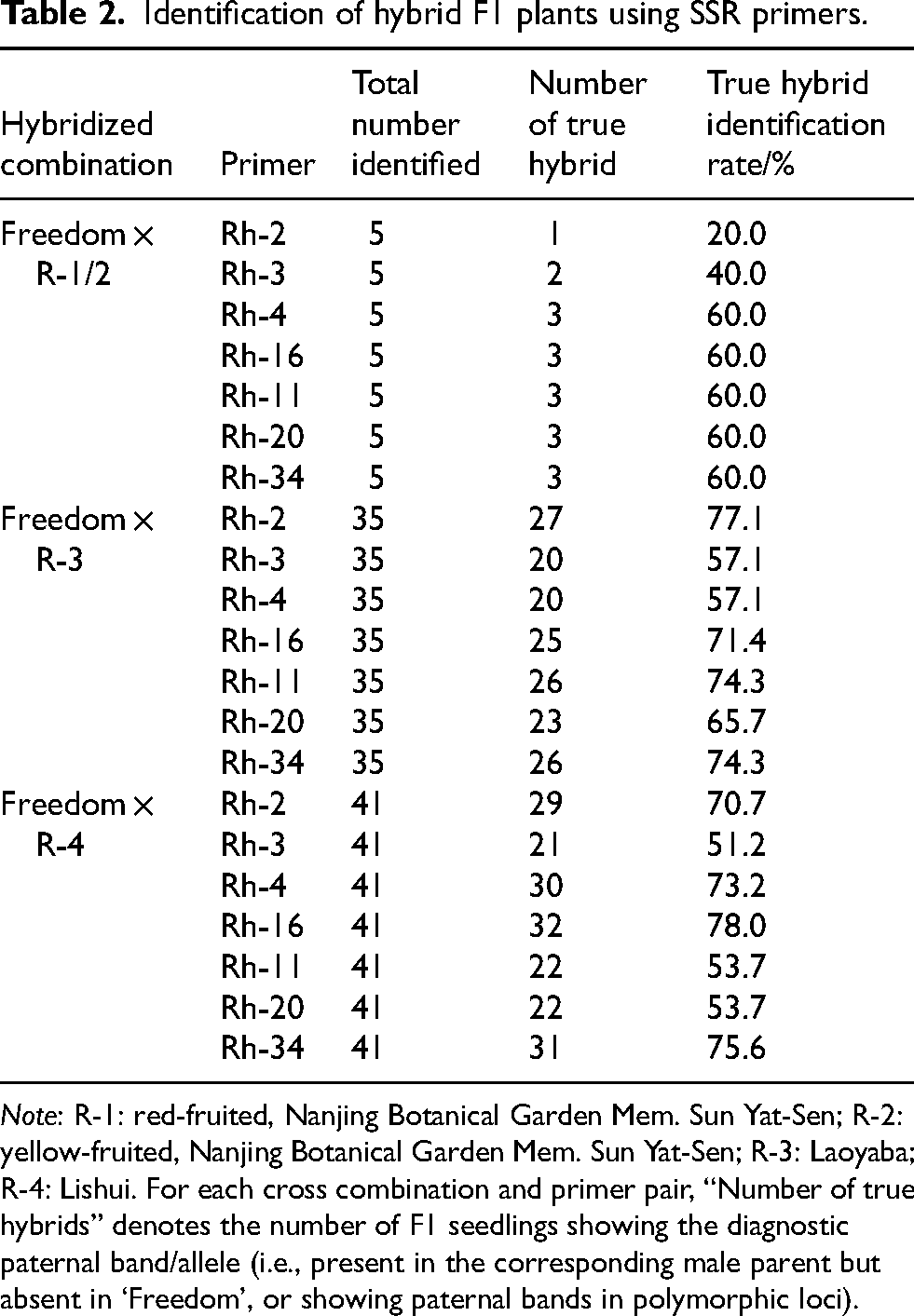
Note: R-1: red-fruited, Nanjing Botanical Garden Mem. Sun Yat-Sen; R-2: yellow-fruited, Nanjing Botanical Garden Mem. Sun Yat-Sen; R-3: Laoyaba; R-4: Lishui. For each cross combination and primer pair, “Number of true hybrids” denotes the number of F1 seedlings showing the diagnostic paternal band/allele (i.e., present in the corresponding male parent but absent in ‘Freedom’, or showing paternal bands in polymorphic loci).
Among the seven primers, Rh-2/Rh-3/Rh-4/Rh-16 were treated as paternal-diagnostic markers (diagnostic bands present in the male parent but absent in ‘Freedom’), whereas Rh-11/Rh-20/Rh-34 provided polymorphic profiles in both parents and served as additional evidence for paternal contribution. The proportion of seedlings showing diagnostic paternal evidence varied among primer pairs and among cross combinations (Table 2). For Freedom × R-1/2, 20.0–60.0% of seedlings were scored as hybrid-positive depending on the primer used. For Freedom × R-3 and Freedom × R-4, hybrid-positive rates ranged from 57.1–77.1% and 51.2–78.0%, respectively (Table 2), indicating that primer performance differs among male genotypes and supporting the need to present results by parentage. In general, we identified a total of 62 true hybrids (3 + 27 + 32), accounting for 76.5% of all F1 generations from the cross combinations.
Genetic diversity analysis
The SSR marker band data of hybrid offspring and parents were analyzed via POPGENE v1.32 software, and a total of 18 polymorphic bands were detected. A total of 18 alleles (Na) were detected, with an average value of 2.57 and a variation range of 2 ∼ 3. The number of effective alleles (Ne) varied from 1.36 to 2.75, with an average value of 1.89, among which primer Rh-20 had the most Ne, and primer Rh-4 had the least Ne. The Shannon information index (I) varied from 0.48 to 1.05, with an average value of 0.70, the largest value appeared at primer Rh-20 and the smallest value appeared at primer Rh-4. The expected heterozygosity (He) varied from 0.27 to 0.64, with an average value of 0.44. The variation of Nei's genetic diversity ranged from 0.26 to 0.64 (Table 3). On the basis of multiple indices, the seven primers selected can be used to identify the genetic diversity of interspecific hybrid offspring of Rubus, among which Rh-20 may be the preferred primer for constructing the fingerprint of hybrid offspring.
Genetic diversity analysis of 7 SSR loci.

Cluster analysis
To further investigate the genetic relationships between the parents and the hybrid-positive seedlings, UPGMA cluster analysis was conducted based on SSR data using NTSYSpc v2.10e, and phylogenetic trees were constructed for the F1 individuals identified as true hybrids in the three cross combinations (Figure 1). Among the three offspring derived from the cross between ‘Freedom’ and R. hirsutus from Nanjing Botanical Garden Mem. Sun Yat-Sen (R-1 and R-2), all clustered in the same group as the red-fruited male parent (R-1); R. hirsutus from Laoyaba (R-3) and R. hirsutus from Lishui (R-4) clustered into one group; and the yellow-fruited male parent (R-2) clustered into the same group as ‘Freedom’ (Figure 1A). Among the 27 true F1 hybrids from the Freedom × R-3 cross, 25 individuals clustered in the same group as the male parent, accounting for 92.6% (Figure 1B). Among the 32 true F1 hybrids from the Freedom × R-4 cross, six individuals clustered with the male parent in Group I (18.8%), whereas Group II contained 25 individuals (78.1%), and R-1 was located within this group (Figure 1C). Overall, the F1 progeny of the Freedom × R-1/2 and Freedom × R-3 crosses exhibited a stronger clustering tendency toward the male parent, whereas the F1 progeny of the Freedom × R-4 cross presented a more dispersed clustering pattern on the phylogenetic tree, indicating a more complex genetic composition and potentially higher genetic diversity and greater selection potential in this cross combination.
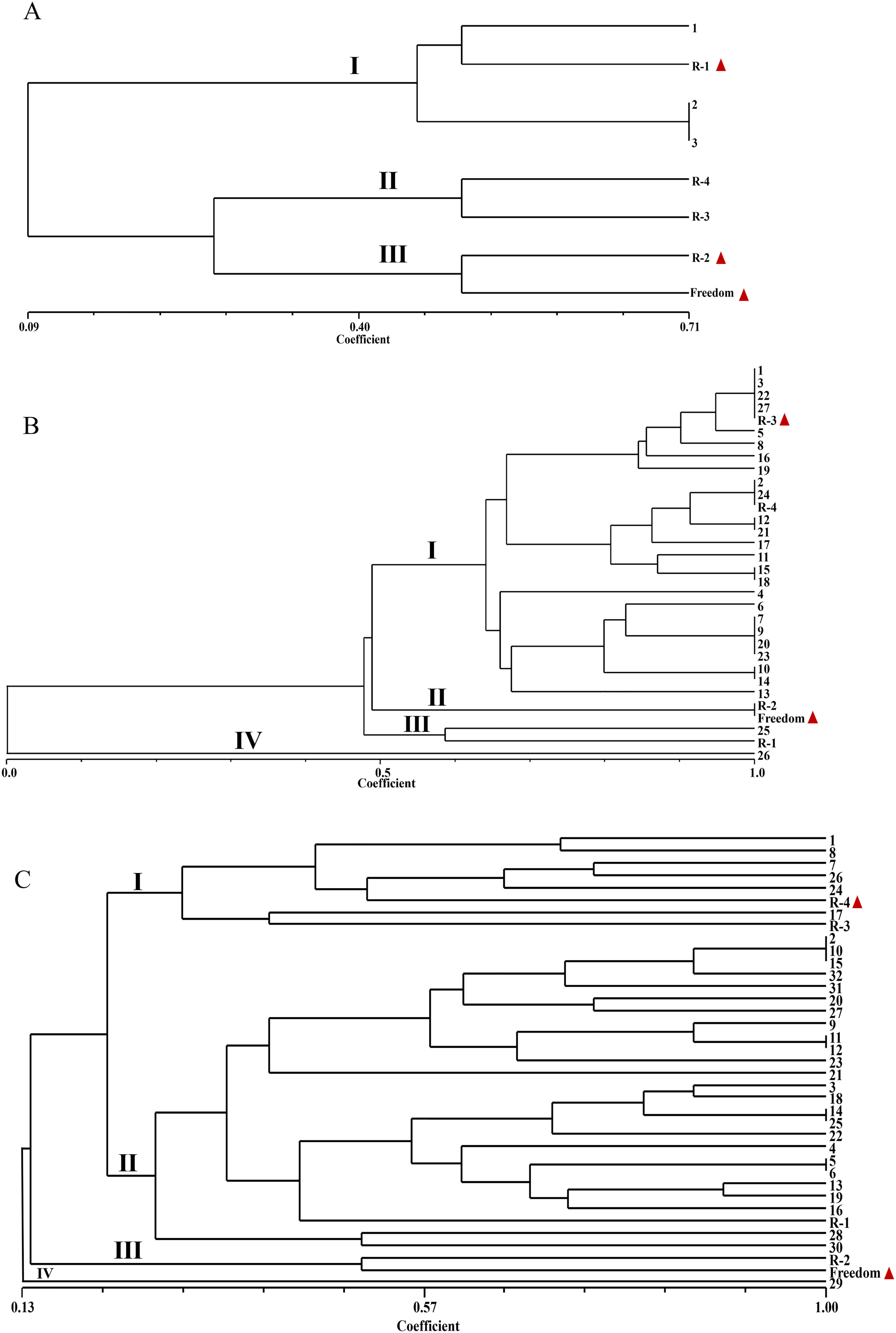
Cluster analysis of three Rubus hybrid combinations.
Ploidy analysis of the parents and hybrid progeny
Based on previous reports, 18 the female parent ‘Freedom’ is a hexaploid cultivar, whereas R. hirsutus is a diploid species (2n = 2x = 14). In the present study, flow cytometry profiles of the parents revealed a single major DNA fluorescence peak for each sample, and the peak position of ‘Freedom’ clearly shifted to a higher fluorescence channel relative to that of R. hirsutus, which is consistent with their known ploidy difference. For the two randomly selected vigorous F1 individuals, their major peaks were located between those of the diploid male parent (R. hirsutus) and the hexaploid female parent (‘Freedom’), and were approximately at the position expected for a tetraploid level when compared with the diploid control (Figure 2).
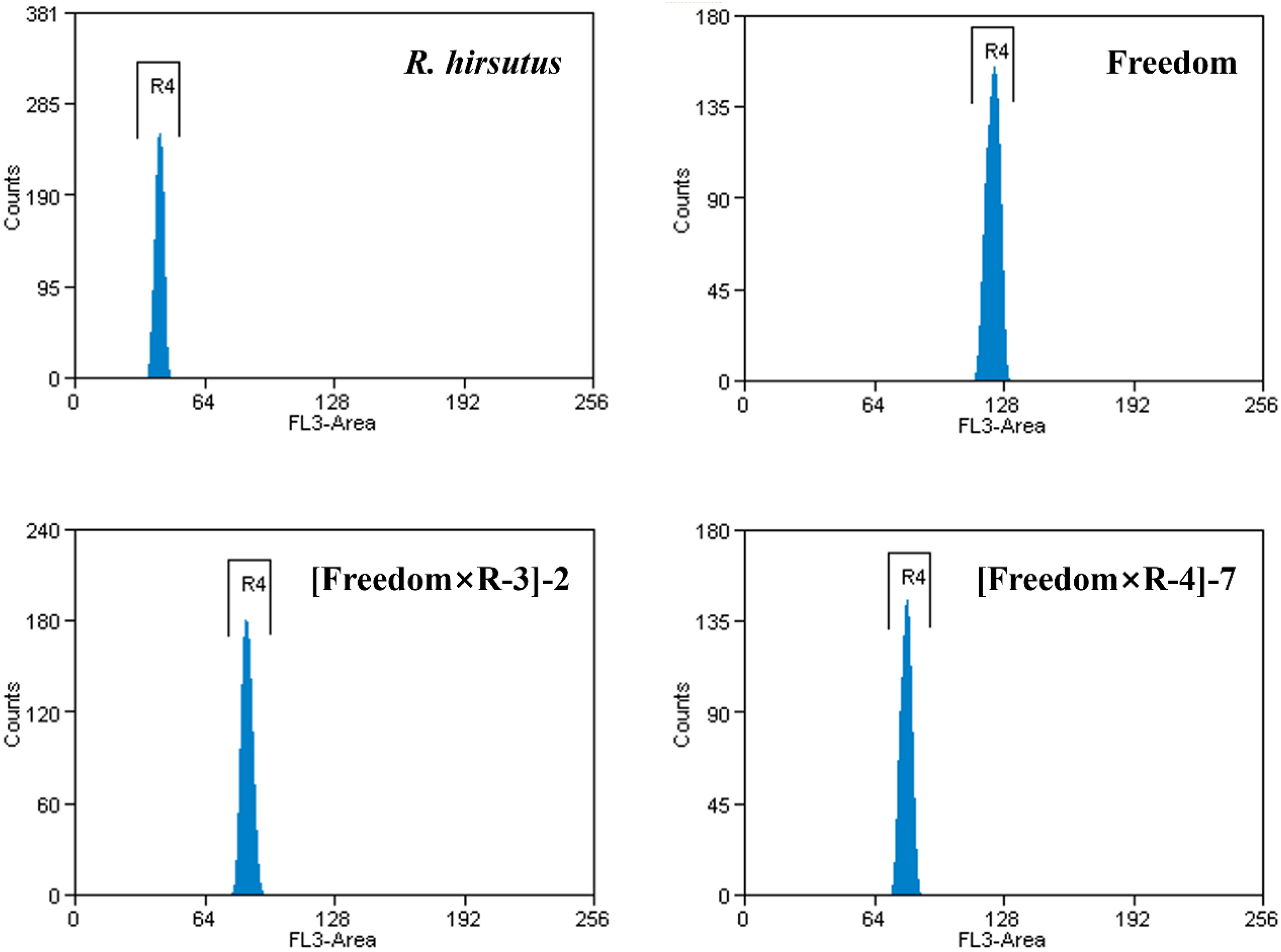
Flow cytometry-based ploidy estimation of the parents and two F1 hybrids.
Phenotypic and physiological analysis
From the identified true hybrids, we selected 26 vigorous seedlings for phenotypic and physiological measurements, including two individuals from Freedom × R-1/2, twelve individuals from Freedom × R-3, and twelve individuals from Freedom × R-4 (Table 4). Considerable variation was observed among the 26 selected hybrids. Plant height ranged from 2 to 55 cm, SPAD readings ranged from 23.3 to 45.6, total nitrogen content ranged from 7.5 to 15.1 g/100 g, and SOD content ranged from 1605.83 to 4314.26 ng/g (Table 4). When the data were analyzed by cross combination, the trait ranges largely overlapped among crosses; nevertheless, the Freedom × R-3 progeny showed broad segregation, with plant heights ranging from 2 to 44 cm and SPAD readings ranging from 23.3 to 45.6. The Freedom × R-4 progeny also displayed pronounced variation, with plant heights ranging from 3 to 55 cm and SOD contents spanning a wider range than those of Freedom × R-3 (Table 4). In contrast, the Freedom × R-1/2 progeny (n = 2) showed limited within-group variation due to the small sample size.
Main phenotype and physiological data statistics of hybrid progeny.
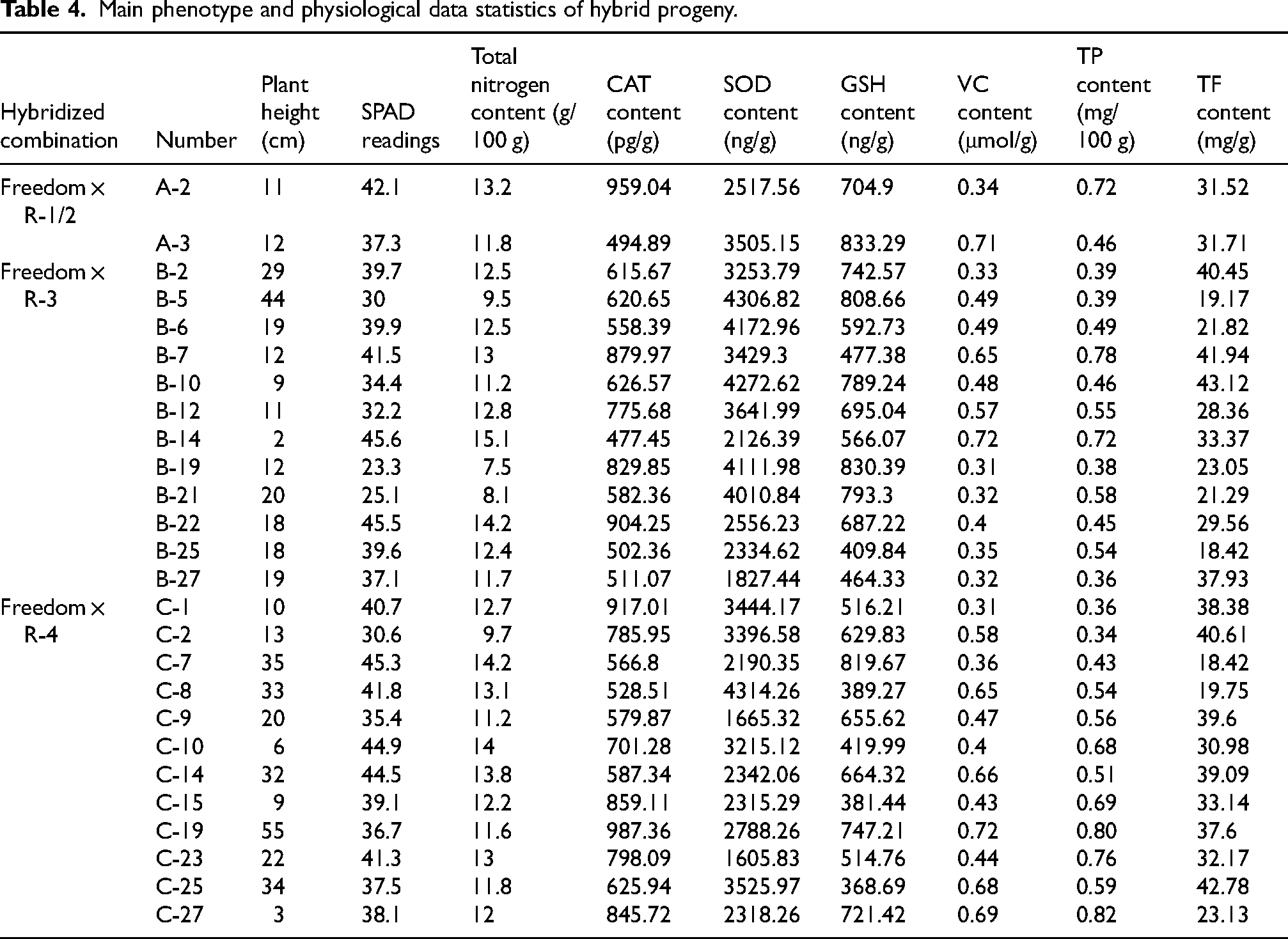
Variation analysis further confirmed the diversity of these traits (Table 5). Among the measured indices, plant height showed the largest coefficient of variation (CV = 0.66), whereas SPAD readings and total nitrogen content exhibited relatively low CV values (0.16 and 0.15, respectively). The CV values of the CAT, SOD, GSH, VC, TP and TF contents ranged from 0.23 to 0.30, indicating moderate variability in the antioxidant-related and secondary-metabolite traits. Overall, these results indicate substantial individual-to-individual variation across the hybrid population, providing a useful basis for early selection of superior genotypes. Kruskal–Wallis tests detected no significant differences among cross combinations for any trait (P > 0.05), indicating that the observed variation is predominantly continuous among individuals rather than forming clearly separated cross-specific groups.
Variation analysis of major traits of true hybrids.

Correlation and principal component analysis
Correlation analysis of 9 traits in the F1 generation revealed associations between multiple indicators (Figure 3A). The SPAD value was positively correlated with total nitrogen content, TP content was positively correlated with VC content, and CAT content was positively correlated with TP and TF contents. In contrast, the SPAD value and total nitrogen content were negatively correlated with SOD and GSH contents, and TP content and TF content were also negatively correlated with SOD and GSH contents. Overall, these traits showed varying degrees of correlation, indicating that they may change together as part of the physiological state of the seedling. Notably, correlation does not imply causation, and these measured traits are more likely to reflect the outcomes of growth and physiological conditions.
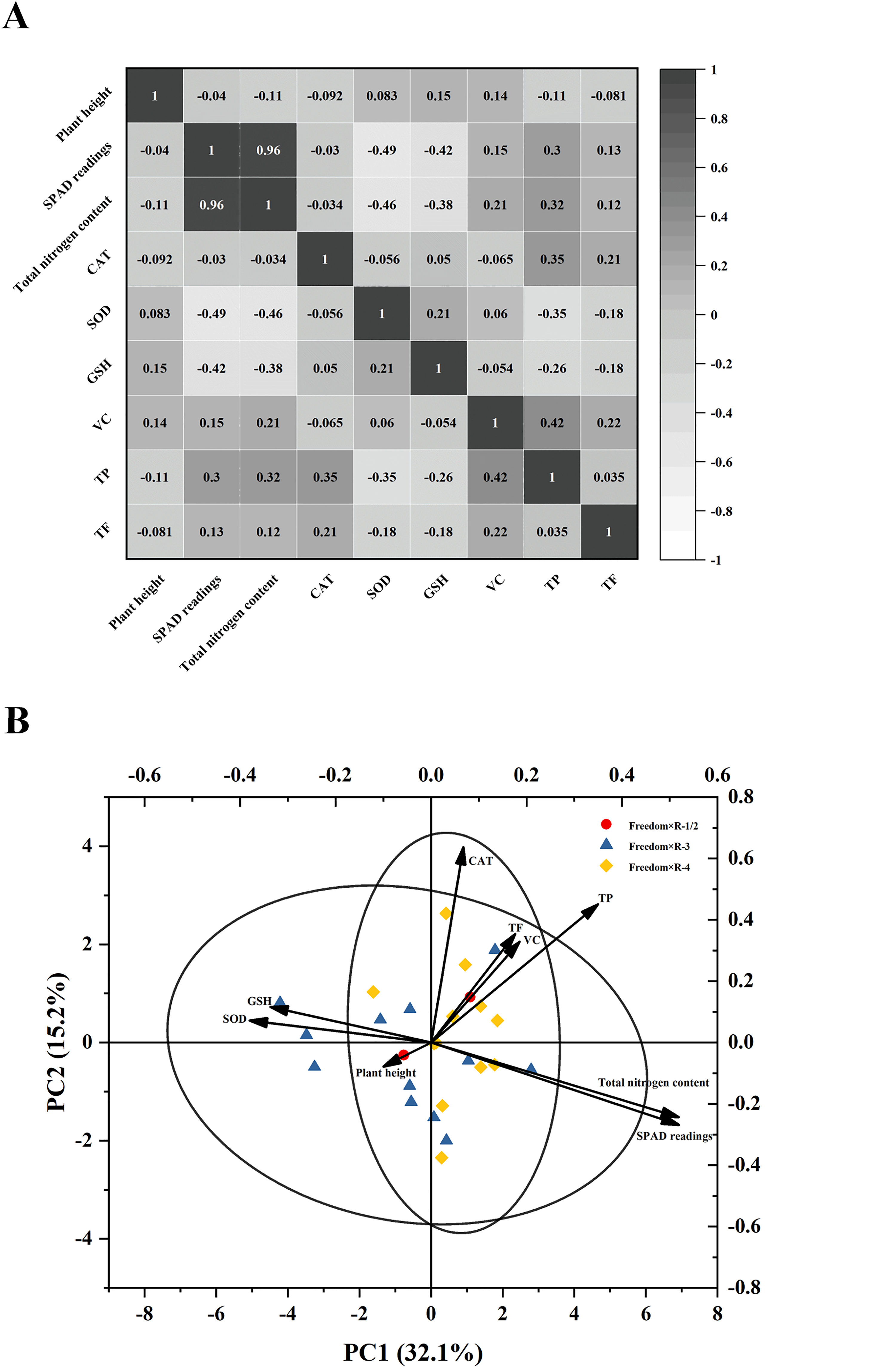
Correlation heatmap (A) and principal component analysis diagram (B) of 9 traits of F1 hybrid generation.
Principal component analysis showed that the contribution rates of PC1 and PC2 were 32.1% and 15.2%, respectively (Figure 3B). The SPAD value, total nitrogen, and TP content contributed significantly to PC1, whereas the SOD and GSH contents were strongly negatively correlated with PC1. CAT mainly contributed to PC2, and TP, VC, and TF were also positively correlated with PC2. As shown in Figure 3B, the three hybrid combinations (Freedom × R-1/2, Freedom × R-3, and Freedom × R-4) had a large amount of overlap, and no obvious separation was observed between the groups, indicating that the variation among individuals was continuous and scattered.
Discussion
Hybrid breeding plays an important role in blackberry and raspberry breeding. Through interspecific hybridization, the excellent genes of different Rubus plants can be identified to develop new cultivars with higher yield, better quality or stronger adaptability. Hybrid breeding practices are key measures for the creation of new blackberry germplasms. The genetic background of different Rubus plants is complex. 19 Not only do germplasms of different ploidy levels have low hybrid affinity, but the post-representative types also vary greatly. It is difficult to judge whether they are hybrids based on phenotype, which brings limitations to hybrid breeding. Therefore, reliable marker-based hybrid authentication is essential for early selection in wide Rubus crosses.
SSR is rich in polymorphisms and has good reproducibility of PCR amplification results. It is a widely used as molecular marker in Rubus plant. 12 The excellent hybrid plant ‘Ningzhi 4’ is thornless, semi-upright or upright, and has the advantages of large fruit, high yield, vigorous growth and good disease resistance. 20 Wu et al. 20 identified the parents of ‘Ningzhi 4’ through SSR markers, providing a realistic basis for the construction of its fingerprint.
In this study, 7 pairs of SSR primers 17 were used to perform hybrid authenticity identification, genetic diversity analysis and cluster analysis on the F1 generation of the hybrid between R. hirsutus and double-cropping blackberry ‘Freedom’. We found that the hybrid offsprings had both heredity and variation. Among the 81 putative F1 seedlings, 62 were confirmed as true hybrids (76.5%), and the hybrid rates differed among cross combinations (Freedom × R-1/2: 3/5; Freedom × R-3: 27/35; Freedom × R-4: 32/41). The observed hybrid rate in this hybridization experiment may be influenced by cross-ploidy barriers (hexaploid × diploid), which can reduce compatibility and increase developmental instability in some interspecific combinations. Similar difficulties have been reported in wide Rubus crosses, where ploidy barriers reduce hybrid fertility and stability. For example, colchicine-induced chromosome doubling has been successfully used to stabilize interspecific hybrids in Rubus, providing a practical approach to facilitate the subsequent selection and utilization of hybrid progeny. 21 Crosses between different ploidy levels may cause meiotic irregularities and affect the stability of hybrid progeny. 22 These findings also indicate that the genetic background of Rubus plants is complicated compared with that of other plant species because of high heterozygosity breeding and the lack of polyploid genome information for Rubus plants, which has led to slow progress in interspecies cross-ploidy breeding of Rubus plants. 23 Zhang et al. 24 used SSR molecular markers to identify six superior strains of blackberry hybrid combinations, and found that the superior strains of the offspring of ‘Kiowa’ × ‘Arapaho’ hybrid combination showed both heterozygous band patterns and band deletion or abnormality. This discovery also implies that blackberry has a complex genetic background, and the hybrid preparation and gene recombination will lead to serious separation of traits, which makes the performance of hybrid offspring full of uncertainty. In this study, the genetic diversity of F1 generation of hybrids varied from 0.26 to 0.64.
Oxidative reactions such as respiratory metabolism in organisms produce active free radicals, and the production of free radicals damages DNA, lipids and proteins. 25 The contents of CAT, SOD, GSH, VC, TP and TF can reflect the antioxidant capacity of plants. Evaluating its oxidative stress status is particularly important. 26 In this study, 9 traits such as plant height, SPAD value, total nitrogen content and antioxidant capacity of hybrid offsprings were analyzed. The results showed that these important traits have rich diversity and high variability in offsprings, but the growth potential of plants may differ significantly due to cross-ploidy hybridization, and their adaptability needs to be further investigated in the future. In the process of cross breeding, the application of correlation analysis and principal component analysis is helpful for extracting key information from complex data, providing a scientific basis for breeding work, and better carrying out variety selection. 27 This study showed that SPAD value, total nitrogen content and TP content have great contributions to PC1, and plant height have great contributions to PC2. All traits influence and correlate with each other. These findings can provide reference for the study of interspecific hybridization of Rubus plants. Future work combining expanded marker sets and multi-year field evaluation will further support efficient selection of superior Rubus interspecific hybrids.
Conclusion
Using seven SSR primers, we authenticated 81 hybrid F1 individuals from ‘Freedom’×R. hirsutus. In total, 62 seedlings (3 from Freedom × R-1/2, 27 from Freedom × R-3, and 32 from Freedom × R-4) were confirmed as true hybrids, representing 76.5% of all tested F1 individuals. Flow cytometry analysis of two vigorous hybrids revealed that the fluorescence peak positions were intermediate between those of the diploid male parent and the hexaploid female parent, supporting a tetraploid level for the tested F1 plants. Among the authenticated hybrids, 26 vigorous seedlings were further evaluated and showed substantial phenotypic and physiological variation, indicating rich segregation and selection potential in this interspecific population. SSR diversity indices (18 polymorphic bands; expected heterozygosity 0.27–0.64) suggest moderate genetic diversity detectable with the selected primer set. Overall, this study provides practical SSR marker information and a workflow for early hybrid authentication and preliminary evaluation of Rubus hybrid progeny, facilitating subsequent selection and utilization of promising germplasms.
Footnotes
Abbreviations
Author contributions
Conceptualization: Yaqiong Wu and Chunhong Zhang; investigation: Lianfei Lyu and Wenlong Wu; Formal analysis: Yaqiong Wu, Mingyue Wang and Haiyan Yang; writing—original draft preparation: Mingyue Wang and Yaqiong Wu; writing—review and editing: Fuliang Cao and Weilin Li. All authors have read and approved the published version of the manuscript.
Funding
The research was supported by the “JBGS” Project of Seed Industry Revitalization in Jiangsu Province (JBGS[2021]021), the National Natural Science Foundation of China (32572105), the Jiangsu Key Laboratory for the Research and Utilization of Plant Resources (JSPKLB202313), and the China Postdoctoral Science Foundation (2025M781855).
Role of the funding source
The funding bodies had no role in the design of the study; the collection, analysis, and interpretation of data; or writing of the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The data and materials supporting the conclusions of this study are included within the article.