
Abstract
Background:
Women with polycystic ovary syndrome (PCOS) exhibit a low fertility by chronic hyperandrogenemia. Different evidence have shown that androgens could regulate the endoplasmic reticulum (ER) homeostasis and glucose metabolism. However, it is unclear whether androgens can exert these effects on human endometrial stromal cells. Our goal was to study the protein content of GRP78 (an ER homeostasis marker) in endometria from women with PCOS and healthy women and to assess the GRP78 protein levels and its relationship with glucose uptake on a human endometrial stromal cell line stimulated with testosterone.
Methods:
Immunohistochemistry assays for GRP78 were performed on endometrial samples obtained from women with PCOS (n = 8) and control women subjected to hysterectomy (n = 8). Western blot analysis for GRP78 and glucose uptake was assessed in a telomerase-immortalized human endometrial stromal cell line (T-HESC) exposed to testosterone for 24 or 48 hours and challenged to an insulin short-term stimulation. Tukey test was performed for human samples comparison. Student t test or ANOVA–Bonferroni test was carried out according to the in vitro experiment. P < .05 was considered as significant.
Results:
GRP78 stromal immunostaining was reduced in PCOS endometria compared to controls (P < .05). The T-HESC shows a testosterone-dependent downregulation of GRP78 protein content (P < .05), concomitant with half-reduction in glucose uptake compared to controls (P < .05). Moreover, enhanced small interfering RNA against GRP78 messenger RNA leads to a decrease in glucose uptake (P < .05). Such effects were reverted by hydroxyflutamide, an inhibitor of androgen receptor.
Conclusion:
These results suggest that hyperandrogenemic PCOS environment could compromise the endometrial homeostasis confirmed by the decrease in glucose uptake induced by testosterone and exhibited by stromal cells.
Introduction
Human endometrium is a cyclic steroid hormonal-sensitive tissue where steroids from ovarian origin command cell proliferation and differentiation in proliferative and secretory endometrial phases, respectively. 1 Endometrial function is crucial in processes such as recognition, adherence, and embryo implantation. It is constituted by luminal glandular epithelia plus stromal compartment with blood vessels, fibroblasts, and several immune cell types such as macrophages. 2 Functionally, endometrial stroma responds to ovarian steroids during the proliferative phase of the menstrual cycle, providing glycogen storage necessary for endometrial decidualization and embryo receptivity. In this context, protein expression of several glucose transporters in human endometrium, such as GLUT3, GLUT8, the insulin-dependent glucose transporter GLUT4, and specially GLUT1, endorses the energetic requirement in this tissue for a cyclic and dynamic metabolic demand, which is necessary for decidualization of stromal cells. 2,3 However, the energetic endometrial homeostasis could be altered as has been observed in women with polycystic ovary syndrome (PCOS). 4,5
Polycystic ovary syndrome is an endocrine–metabolic disorder present in 5% to 10% of women in fertile age, characterized by a clinical and/or biochemical hyperandrogenism. 6 Also, a higher pregnancy failure risk with a 30% to 50% probability of miscarriage in women with PCOS, compared to normal population, has been reported. 1 The endometria from these women exhibit several molecular alterations that disrupt the endocrine and metabolic function of the tissue. In this context, PCOS endometria have shown several alterations in the insulin signaling pathway, GLUT4 gene expression, and its protein content, suggesting impairment on the endometrial reproductive functions in these patients. Moreover, the hyperandrogenemic environment in women with PCOS could affect glucose homeostasis in endometrial cells 7 –9 and in other tissues such as adipose and skeletal muscle. 10,11 Nevertheless, how this environmental condition deregulates the endometrial homeostasis is still unclear. Several studies in animal and in vitro models show that endoplasmic reticulum (ER) stress is involved in the insulin resistance development and changes in energy metabolism. 12 –14 In this regard, androgens promote the expression of ER chaperones, including GRP78 in prostate cells, in vitro and ex vivo. 15,16 GRP78 is a Ca2+-dependent ATPase chaperone involved in the relief of ER stress, increasing the folding activity in the ER lumen. 17 Also, GRP78 is implicated in maintaining glucose uptake in glucose storage tissues. 12 However, it is unclear whether GRP78 regulates glucose uptake in human endometrial stromal cells and whether androgens can control GRP78 protein levels and glucose uptake in these cells.
Given the responsiveness of human endometrial tissue to androgens, as observed in endometria from women with PCOS having hyperandrogenemia, we studied ex vivo the GRP78 protein content in endometria from women with PCOS compared to fertile patients, as well as in vitro in a human endometrial stromal cell line exposed to testosterone where glucose uptake was assessed.
Materials and Methods
Participants
In this study, 16 patients were recruited: 8 women with PCOS having hyperandrogenemia and 8 fertile healthy patients as controls. The endometrial samples were obtained with a Pipelle suction curette from the corpus of the uteri from patients with PCOS. In the case of controls, the samples were obtained at the time of hysterectomy due to benign uterine pathology. The endometrial samples from both groups were obtained during the proliferative phase, using Noyes criteria 18 by an experienced pathologist. The diagnosis of PCOS was according to the Androgen Excess and PCOS Society criteria. 19 Hyperandrogenemia condition was considered in patients with a free androgen index (FAI) higher than 4.5. The FAI was calculated as the ratio between total testosterone and sex hormone-binding globulin (SHBG) plasma levels, as in previous studies. 9 –20 Blood glucose and insulin levels were evaluated by an oral glucose tolerance test (OGTT) with 75 g load of glucose. Additionally, homeostasis model assessment (HOMA) index and insulin sensitive index (ISI) composite were calculated for all patients. The exclusion criteria were women who presented hyperprolactinemia (prolactin >35 ng/mL), hypothyroidism (thyroid-stimulating hormone >5 mUI/mL), androgen-secreting tumors (total testosterone >2 ng/mL; dehydroepiandrosterone sulfate >3600 μg/dL), Cushing syndrome (urine cortisol concentration >50 μg/dL at 24 hours and fasting plasma concentration of cortisol >25 μg/dL), congenital adrenal hyperplasia (17-OH progesterone >2.5 ng/mL), diabetes or treatment with hormones within 3 months prior to the recruitment into the study and/or ovulation induction.
Ethical Committees from the Faculty of Medicine and Clinical Hospital from the University of Chile approved this research where all patients signed an informed written consent to be included in this study.
Tissue Assessment
Endometrial tissue samples were fixed in 4% buffered formaldehyde for 24 hours, embedded in paraffin, and cut into 5-μm thick sections before histological and immunohistochemical studies.
Immunohistochemical Assay
Paraffin sections of human endometrial tissue were deparaffinized in xylene and gradually hydrated through graded alcohols. The sections were incubated in 10 mmol/L sodium citrate buffer (pH 6.0) at 95°C for 30 minutes, incubating the samples in 3% hydrogen peroxide for 15 minutes to prevent endogenous peroxidase activity. Nonspecific antibody binding was inhibited by incubating samples with a commercial blocking solution (Histostain-SP; Invitrogen, Maryland) for 15 minutes. Primary antibody to GRP78 (1:500, monoclonal; Cell Signaling Technology, Massachusetts) was applied to the samples and incubated overnight at 4°C. The internal control was carried out on adjacent sections incubated without the primary antibody. A biotinylated horseradish peroxidase secondary antibody was used for the detection of immune signal. Chromogenic revealed was developed by the streptavidin-peroxidase system, and 3,3′-diaminobenzidine was used as the substrate; counterstaining was performed with hematoxylin (Dako, California). The slides were evaluated on an (Olympus BX51, Japan) optical microscope. Slide analysis was performed by the measurement of positive pixel intensity with the use of the semiquantitative analysis tool integrated optical density, from Image Pro Plus 6.0 program. Equally sized areas were taken at random in the stroma and epithelia in different regions of the sample.
Cell Culture and Treatments
A commercial telomerase human endometrial stromal cell line responsive to steroid hormones (T-HESC; ATCC, CRL-4003), obtained from primary cultures of endometrial stroma, 21 was used for in vitro studies, as reported previously. 7,20,22,23 The T-HESC cells were cultured in growth media (Dulbecco modified eagle medium [DMEM] Ham F-12 medium, 10% fetal bovine serum treated with dextran carbon, puromycin 500 ng/mL, and 1× antimycotic/antibiotic) at 37°C in 5% CO2 atmosphere until confluence was achieved. After 80% confluence, cells were cultured in 6-well plates, 700 000 cells/plate in growth media with 2% fetal bovine serum treated with dextran carbon, at 37°C in a 5% CO2 atmosphere. The cultures were further subjected for 24 or 48 hours to testosterone 100 nmol/L (Sigma-Aldrich, Germany) according to previous studies. 7,10,20 Furthermore, after testosterone treatment, some cell cultures were challenged with 100 nmol/L insulin short term (20 minutes) to assess glucose uptake. Once the cultures were stimulated, the media was aspirated and the cells were lysated with radioimmunoprecipitation assay buffer and processed as described previously. 7,8,20 . To determine the activation of androgen receptor by testosterone, the cells were incubated with hydroxyflutamide 10 μmol/L for 30 minutes previous to the addition of testosterone 100 nmol/L, as reported. 7 To silence the GRP78 translation, T-HESC cultures were incubated with an enhanced small interfering RNA (esiRNA) overnight (200 nmol/L) in Opti-MEM® (California) and replaced with DMEM Ham F-12 medium, 2% of fetal bovine serum treated with dextran carbon, at 37°C in a 5% CO2 atmosphere prior to hormonal stimulation.
Western Blot Assay
Total proteins (30 μg from cultured cells) were denatured and fractionated using 10% or 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels. The protein content was transferred to a nitrocellulose membrane, and the protein transference was confirmed by Ponceau S staining. The membranes were blocked for 1 hour in TTBS (20 mmol/L of Tris, pH 7.5; 137 mmol/L of NaCl; and 0.1% Tween-20) with 5% non-fat milk. The membranes were washed twice for 5 minutes with TTBS and incubated with anti-GRP78 (1:500, #3177 rabbit monoclonal; Cell Signaling) overnight at 4°C. The membranes were then washed twice for 5 minutes with TTBS and incubated for 1 hour at room temperature with anti-rabbit immunoglobulin G (IgG) peroxidase-linked species-specific antibody (1:5000; Kirkegaard & Perry Laboratories [KPL], Maryland). β-Actin was detected as loading control (1:20000, mouse monoclonal; Sigma-Aldrich and 1:10000, antimouse IgG peroxidase linked; KPL, 30 minutes at room temperature). After antibody incubation, the membranes were washed with TTBS 3 times for 5 minutes, and the bound antibodies were detected with enhanced chemiluminescence substrate (Western Lightning Plus-ECL; Perkin-Elmer, Massachusetts). Band intensities were quantified by scanning densitometry utilizing the UN-SCAN-IT software, Automated Digitizing System, version 5.1.
Glucose Uptake Assay
Cells were plated in 12-well plates at 1.5 × 104 cells/well. After hormone treatment described previously, the cell cultures were incubated in transport media containing 15 mmol/L 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid buffer pH 7.6, 135 mmol/L NaCl, 5 mmol/L KCl, 1.8 mmol/L CaCl2, 0.8 mmol/L MgCl2, and 10 mmol/L deoxy-
Statistical Analysis
Comparison groups were analyzed by Tukey test for human samples comparison, and Student t test or ANOVA and Bonferroni test was carried out according to the in vitro experiment. P value <.05 was considered as significant. Statistical tests were performed using GraphPad Prism 6 software.
Results
Clinical and Endocrine Features of Controls and Patients With PCOS
Clinical and hormonal characteristics of women participants in this study are summarized in Tables 1 and 2. Women with PCOS exhibit hyperandrogenemia, according to higher levels of total testosterone and a decrease in circulating SHBG levels, both expressed in higher FAI in the PCOS group compared to control patients (Table 1; P < .05). Body mass index was similar between patients with PCOS and controls (Table 1). Levels of steroid ovarian hormones, 17β-estradiol and progesterone, in the PCOS group were similar to controls (Table 1). Moreover, PCOS and control groups showed similar glucose and insulin values, basal and post OGTT, as well as HOMA-insulin resistance (IR) and ISI composite (Table 2). Histology of endometrial samples was similar in both studied groups and correlate with the proliferative phase of endometrium (data not shown), as we have reported previously. 9 –26
Age, Body Mass Index, and Circulating Steroids Hormones of Controls and Patients With PCOS.a
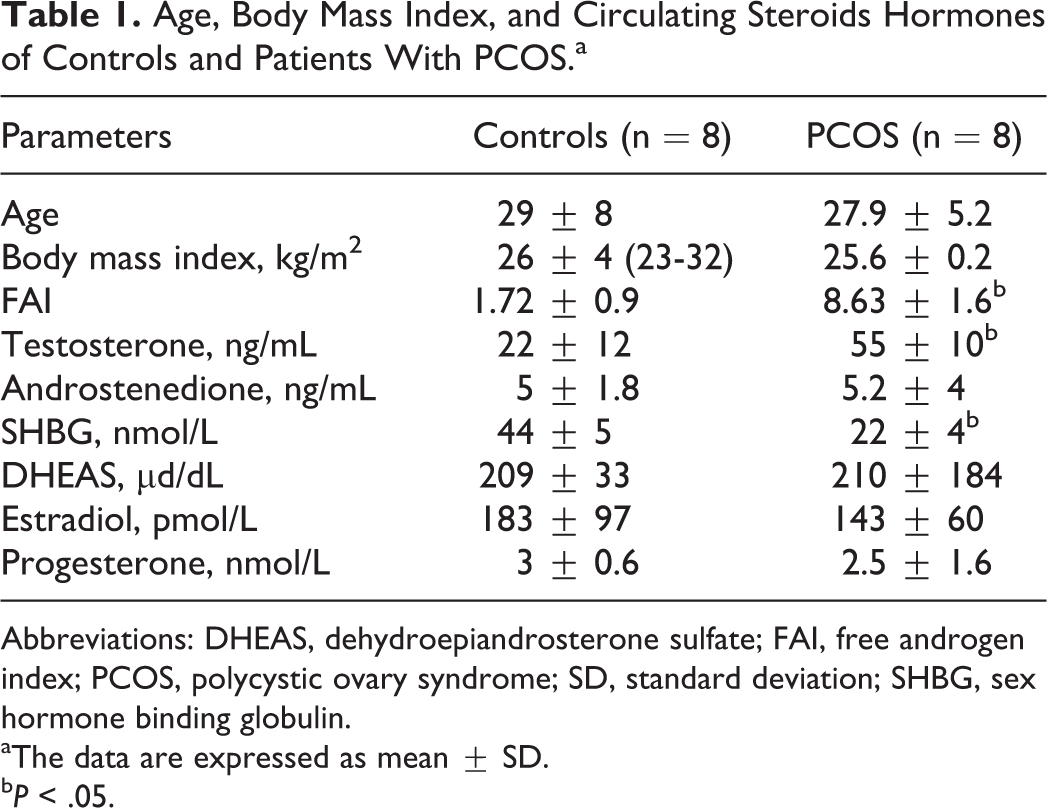
Abbreviations: DHEAS, dehydroepiandrosterone sulfate; FAI, free androgen index; PCOS, polycystic ovary syndrome; SD, standard deviation; SHBG, sex hormone binding globulin.
aThe data are expressed as mean ± SD.
bP < .05.
Metabolic Parameters of Controls and Patients With PCOS.a
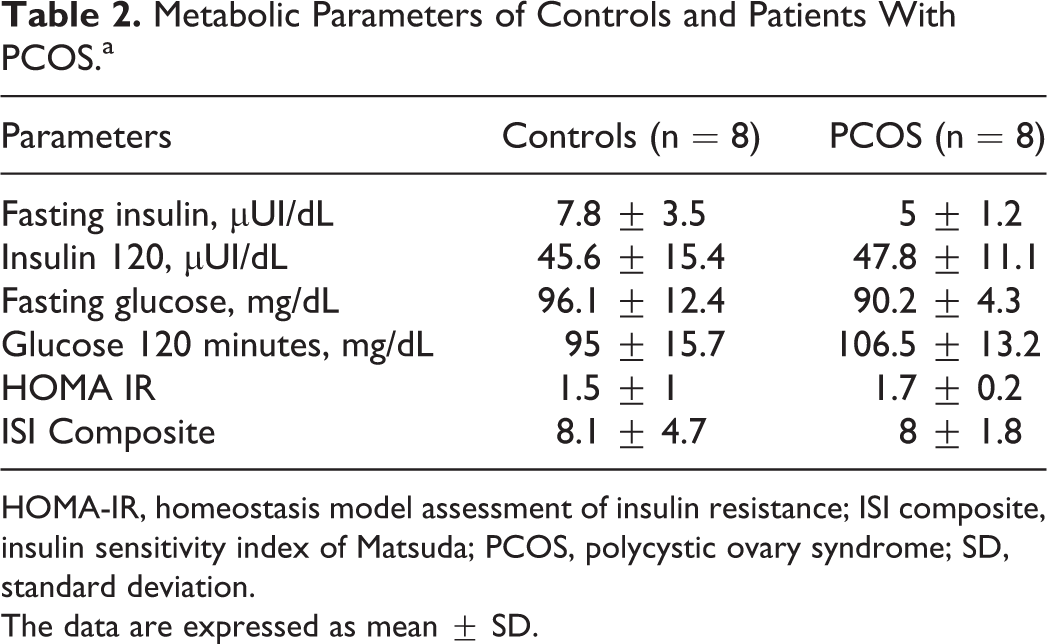
HOMA-IR, homeostasis model assessment of insulin resistance; ISI composite, insulin sensitivity index of Matsuda; PCOS, polycystic ovary syndrome; SD, standard deviation.
The data are expressed as mean ± SD.
GRP78 Protein Expression in Human Endometrial Samples From Women With PCOS
To evaluate the levels and distribution of GRP78 protein in the endometrial compartments, immunohistochemistry assays were carried out in both patients with PCOS and controls. Protein levels of GRP78 were similar in epithelial compartment of endometria from both studied groups of participants. However, the percentage of GRP78-positive cells (controls: 88% ± 10%; PCOS: 35% ± 9%) as well as the median value of GRP78 protein levels decreased around 50% in stromal cells from PCOS endometria compared to the control group (Figure 1; P < .05).
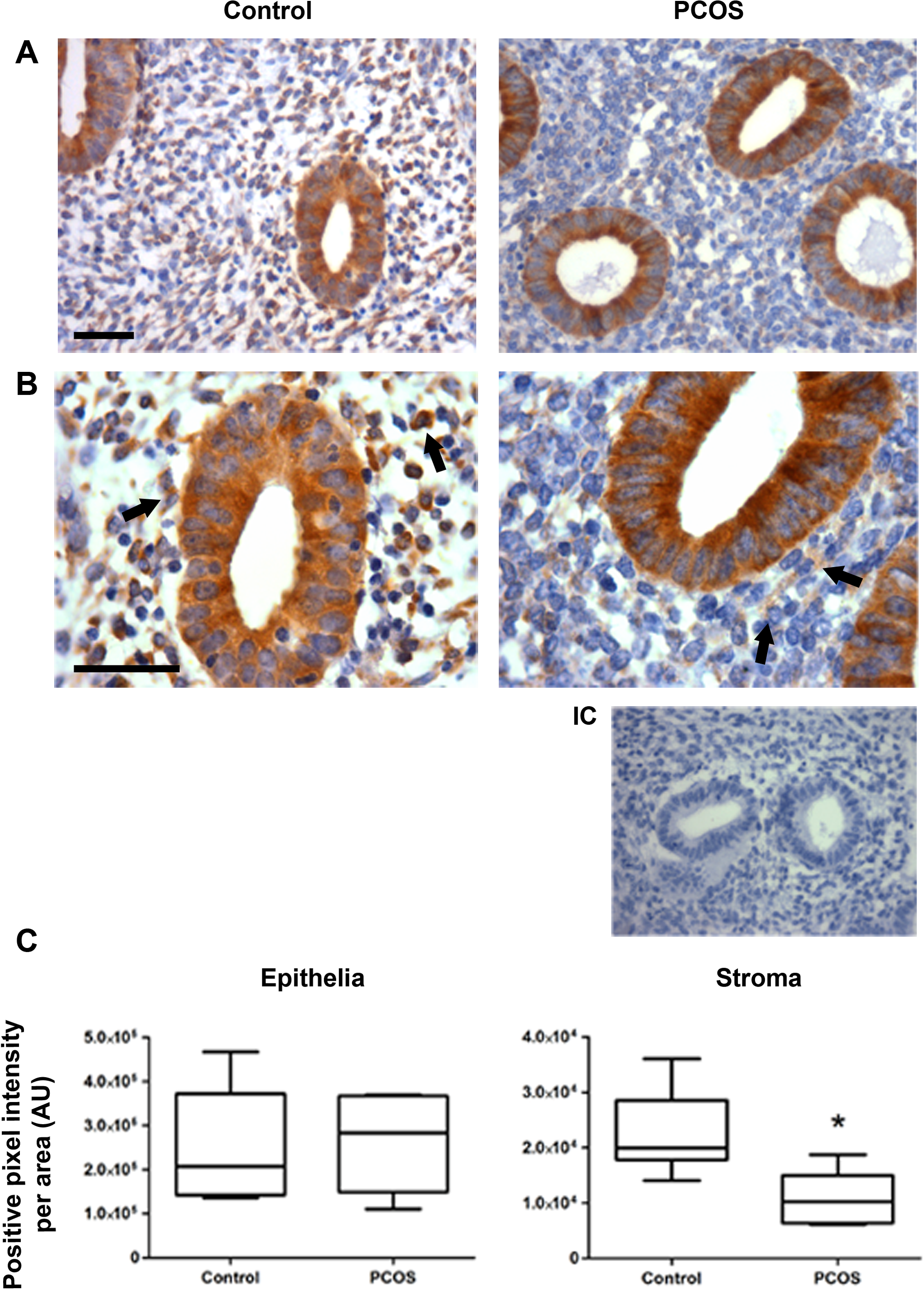
Immunohistochemical assay of GRP78 protein in endometrial samples. A, Representative immunostaining of GRP78 protein in controls (n = 8) and polycystic ovary syndrome (PCOS) endometria (n = 8) magnified at ×400. Bar = 50 μm. B, Magnification of control and PCOS images at ×1000. Bar = 50 μm (IC, internal control without primary antibody). C, Semiquantitation of GRP78 protein levels in endometrial epithelia and stroma from controls and PCOS endometria, expressed in arbitrary units (AU). Arrows indicate positive GRP78 immunostaining. The data are shown as median ± standard deviation (SD). *P < .05.
GRP78 Protein Levels and Glucose Uptake in Testosterone-Treated T-HESC Cells
To mimic the hyperandrogenic environment, the effects of testosterone were tested protein levels of GRP78 in T-HESC cell cultures incubated with testosterone 100 nmol/L for 24 or 48 hours, and GRP78 protein content was evaluated by Western blot assays. As shown in Figure 2, after 48 hours of testosterone treatment, GRP78 protein levels were reduced around 40% by testosterone, compared to untreated cells (Figure 2C and D; P < .05). Nevertheless, testosterone stimulation had no effect on GRP78 protein levels at 24 hours of culture (Figure 2A and B). In order to test whether testosterone regulates glucose uptake in T-HESC cultures, insulin short-term stimulation and tritiated 2-deoxyglucose uptake assays were carried out in the same cell passages utilized for GRP78 Western blots, after 24 or 48 hours of testosterone treatment. After 48 hours of testosterone stimulus, glucose uptake decreased by 50% compared with control cells (Figure 3B; P < .05), whereas, 24 hours after testosterone stimulation, cell cultures exhibit similar levels of glucose uptake compared to untreated cells (Figure 3A).
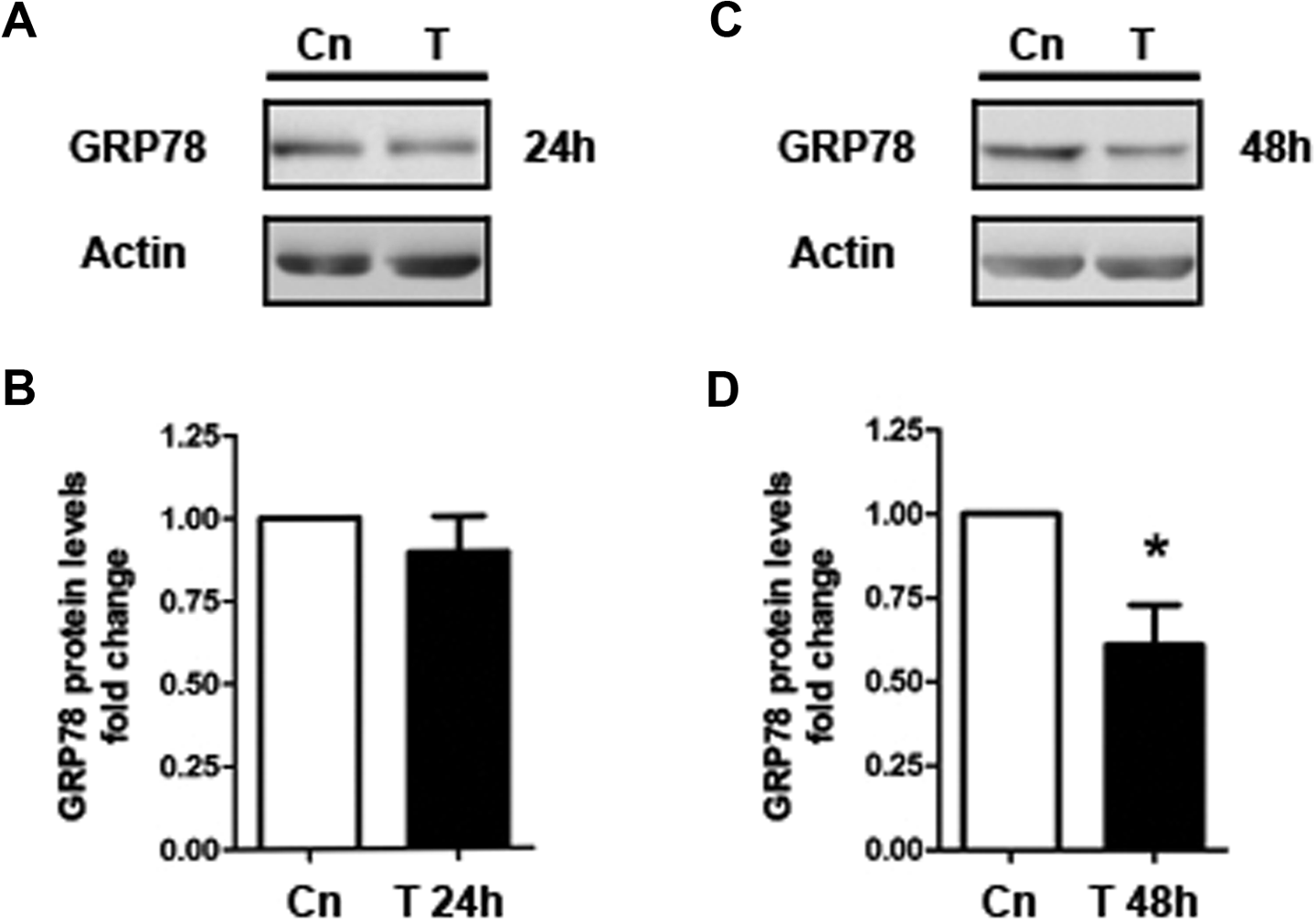
GRP78 protein levels in testosterone-treated telomerase-immortalized human endometrial stromal cell line (T-HESC) cultures. A, Representative gel of Western blot assay of GRP78 after 24 hours of testosterone stimulation (T). B, Semiquantitation of GRP78 protein content after testosterone treatment for 24 hours normalized with β-actin protein levels. C, Western blot assay for GRP78 after 48 hours of testosterone stimulation. D, Semiquantitation of GRP78 protein content after testosterone treatment for 48 hours normalized with β-actin protein content. The data from 3 independent experiments in duplicate are expressed as mean fold change with respect to untreated controls (Cn) ± standard error (SE). *P < .05.
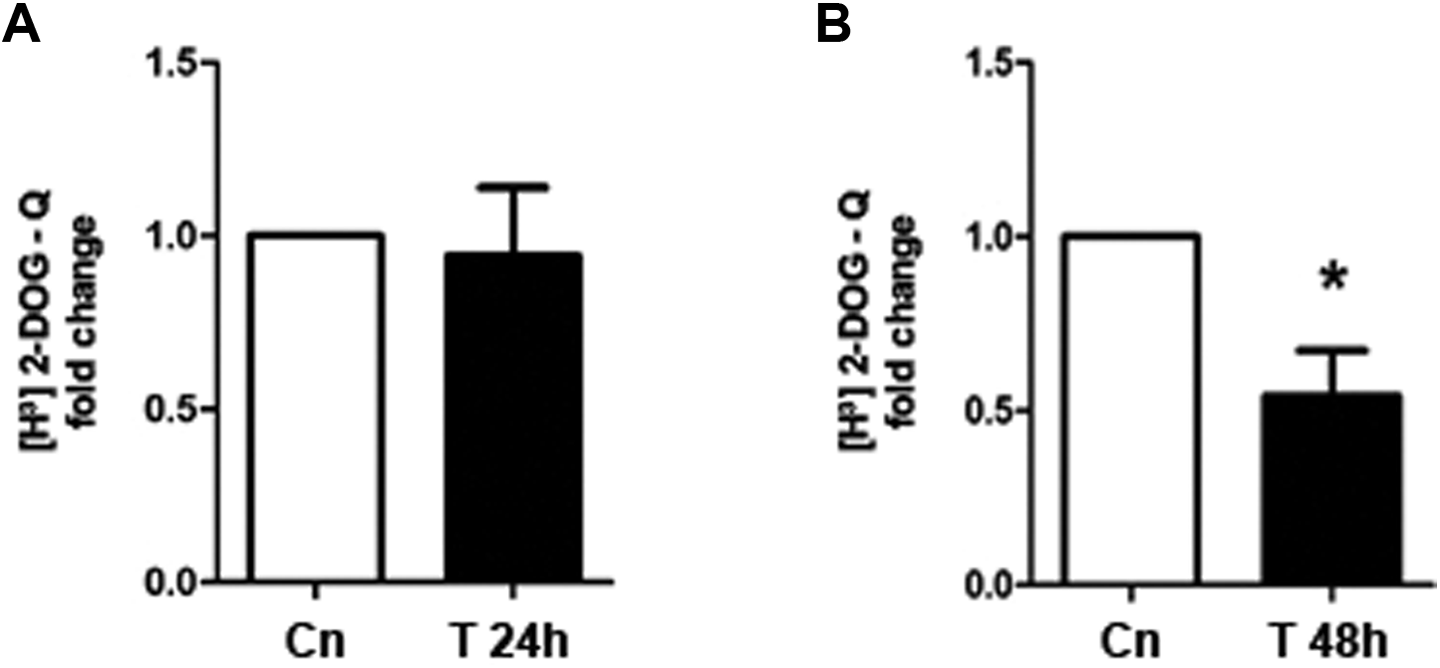
[H3]-2-Deoxyglucose uptake in testosterone-treated telomerase-immortalized human endometrial stromal cell line (T-HESC) cultures. Cell cultures from the same cell passage used for immune blots of GRP78 were incubated with testosterone (T) for 24 or 48 hours followed by 20-minute insulin stimulation (100 nmol/L). A, [H3]-2-Deoxyglucose uptake after 24 hours of testosterone treatment. B, [H3]-2-Deoxyglucose uptake after 48 hours of testosterone treatment. The data from 5 independent experiments in duplicate are expressed as mean fold change with respect to untreated controls (Cn) ± standard error (SE). *P < .05.
GRP78 Protein Levels and Glucose Uptake in T-HESC Cells Treated With Hydroxyflutamide
To confirm the specificity of testosterone effects on GRP78 protein levels and glucose uptake, T-HESC cultures were preincubated with hydroxyflutamide, an inhibitor of the androgen receptor, previously to 48 hours of testosterone stimulation. As expected, GRP78 protein levels decreased 30% by testosterone 100 nmol/L, compared to untreated cells, but not in testosterone-stimulated cells exposed to hydroxyflutamide, where protein levels of GRP78 were similar to cell cultures treated with the inhibitor alone (Figure 4A and B; P < .05). Likewise, glucose uptake was reduced to 50% by testosterone stimulation and was unchanged when androgen receptor activity was blocked (Figure 5; P < .05).
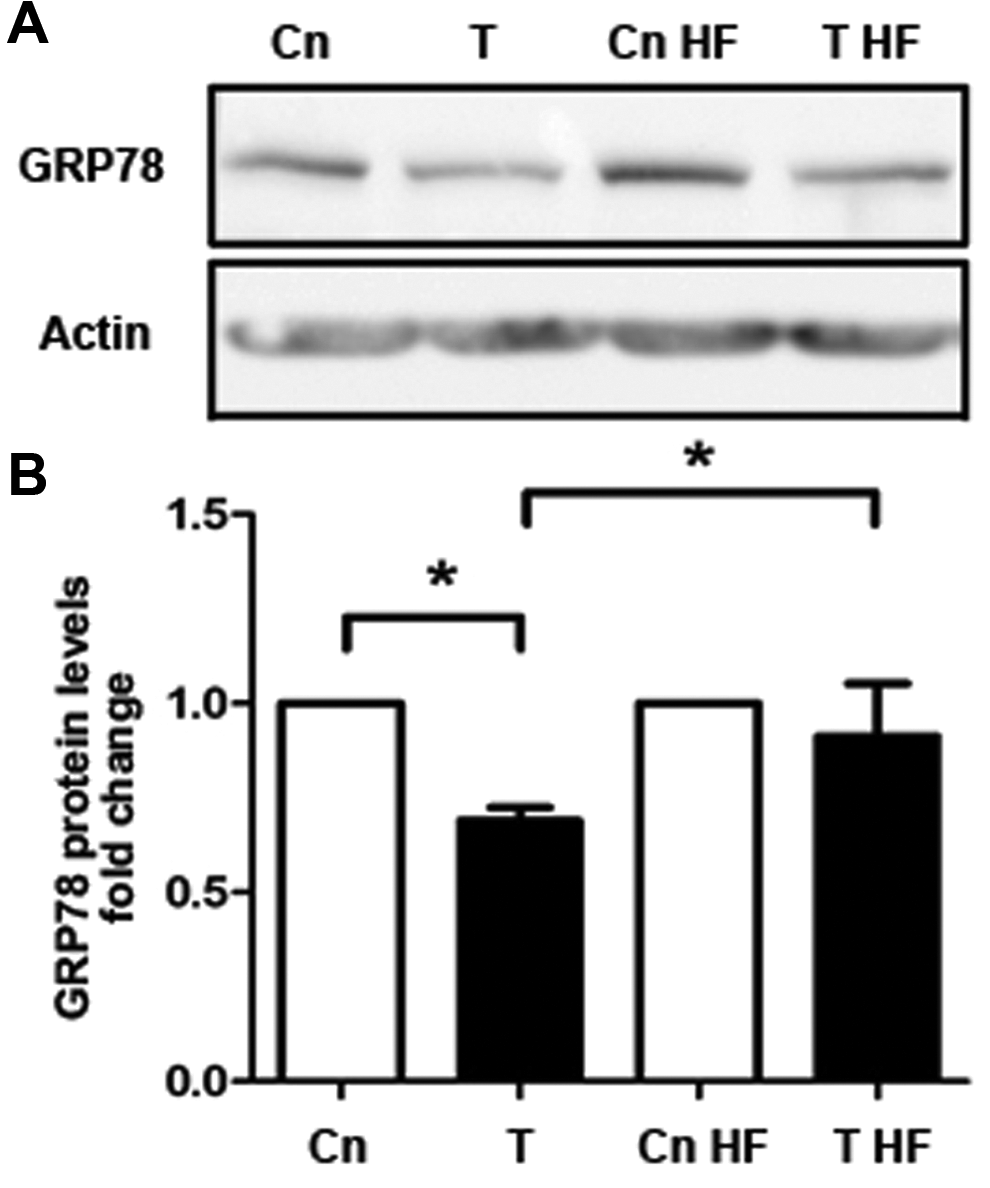
GRP78 protein levels in telomerase-immortalized human endometrial stromal cell line (T-HESC) cultures treated with hydroxyflutamide (HF) and testosterone. Cell cultures preincubated for 30 minutes with hydroxyflutamide were stimulated with testosterone (T, 100 nmol/L) for 48 hours followed by short-term insulin (20 minutes, 100 nmol/L) stimulation. A, Representative immunoblot of GRP78 in cells stimulated with testosterone with or without HF. B, Semiquantitation of GRP78 protein levels. Western blot assays were normalized with β-actin protein content. The data from 5 independent experiments in duplicate are expressed as mean fold change with respect to untreated controls (Cn) ± standard error (SE). *P < .05.
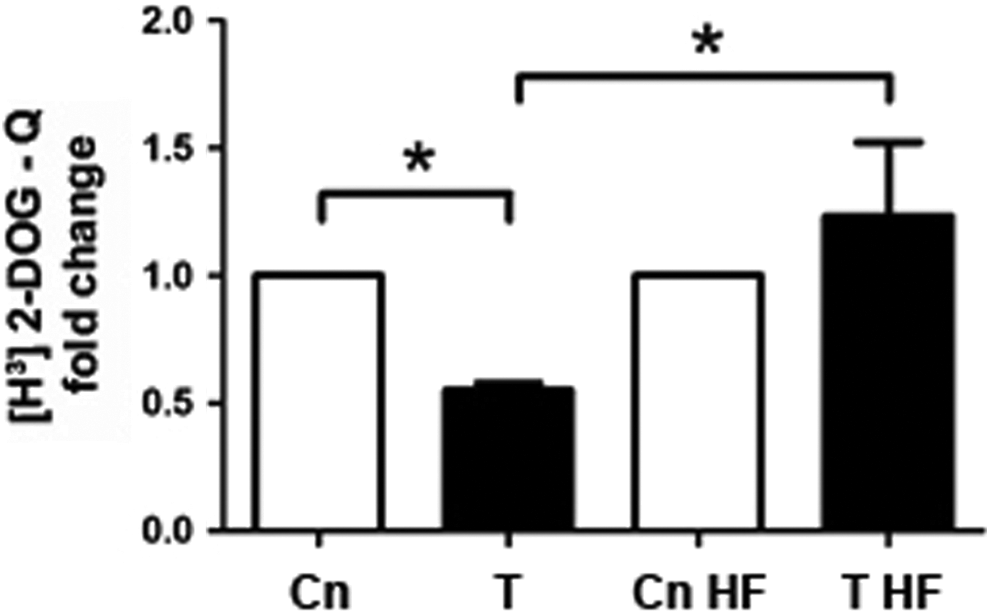
[H3]-2-Deoxyglucose uptake in telomerase-immortalized human endometrial stromal cell line (T-HESC) treated with hydroxyflutamide and testosterone. Cell cultures preincubated for 30 minutes with hydroxyflutamide (HF) were stimulated with testosterone (T, 100 nmol/L) for 48 hours followed by short-term insulin (20 minutes, 100 nmol/L) stimulation. [H3]-2-Deoxyglucose uptake assays were assessed in the same cell passage for GRP78 immunoblots, with or without hydroxyflutamide. The data from 5 independent experiments in duplicate are expressed as mean fold change with respect to untreated controls (Cn) ± standard error (SE). *P < .05.
Glucose Uptake in T-HESC Cultures Incubated With an siRNA to GRP78 Transcript
To elucidate a possible role of GRP78 on glucose uptake in stromal endometrial cells, T-HESCs were transfected with a commercial esiRNA to reduce the GRP78 translation, prior to a short-term insulin stimulus (100 nmol/L for 20 minutes). As shown in Figure 6, in T-HESC cultured cells incubated with the esiRNA with or without insulin for 20 minutes, GRP78 protein levels diminished by 50% compared to non-esiRNA–incubated cell cultures (Figure 6A and B; P < .05). On the other hand, glucose uptake was increased a 30% in short-term insulin-stimulated cells compared to untreated cell cultures (Figure 7A). However, in silenced GRP78 cells and stimulated with a short-term insulin, glucose uptake was inhibited by 75%, compared to insulin-treated cells without esiRNA (Figure 7A and B; P < .05). Interestingly, insulin-untreated cultures showed a diminished glucose uptake when GRP78 translation was reduced by the esiRNA (Figure 7A and B; P < .05).
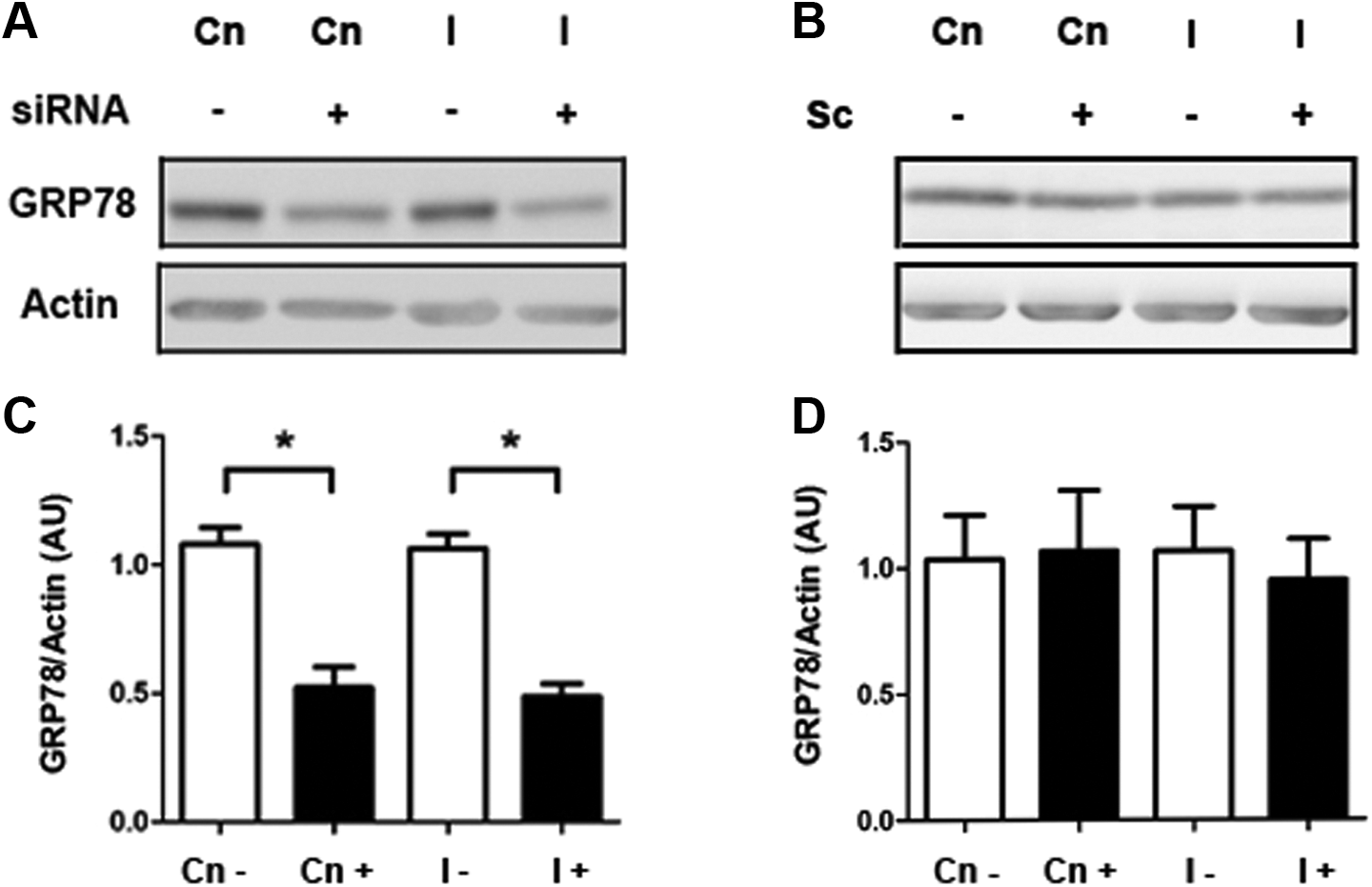
GRP78 protein levels in telomerase-immortalized human endometrial stromal cell line (T-HESC) cultures treated with a GRP78-enhanced small interfering RNA (esiRNA). A, Representative immunoblot of GRP78 in cell cultures preincubated with an esiRNA to GRP78 overnight followed by insulin short-term stimulation (100 nmol/L, 20 minutes). B, Semiquantitation of GRP78 protein levels in cells incubated with an esiRNA to GRP78. C, Immunoblot of GRP78 in cell cultures preincubated with a scrambled siRNA (Sc). D, Semi-quantitation of GRP78 protein levels in cells incubated with Sc, followed by insulin short-term stimulation (20 minutes). Western blot assays were normalized with β-actin protein content. The data from 3 independent experiments in duplicate are expressed as mean fold change with respect to untreated controls (Cn) ± standard error (SE). *P < .05.
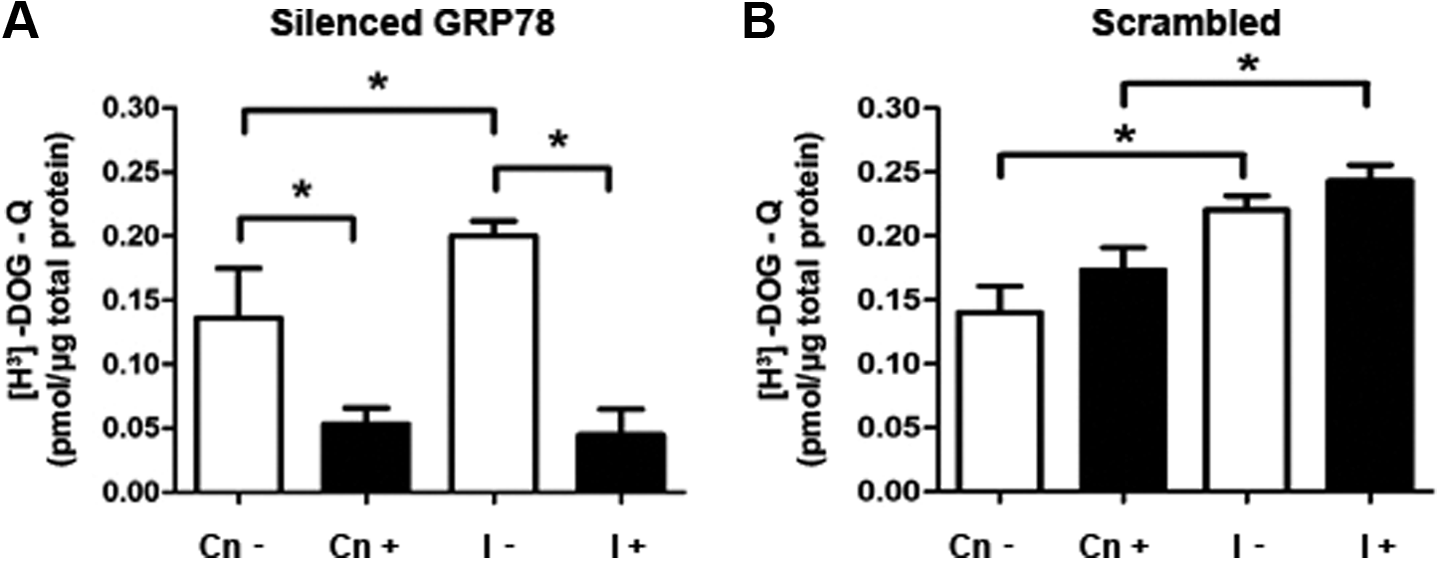
[H3]-2-Deoxyglucose uptake in telomerase-immortalized human endometrial stromal cell line (T-HESC) cultures treated with an enhanced small interfering RNA (esiRNA) to GRP78. A, [H3]-2-Deoxyglucose uptake assays in cell cultures preincubated with an esiRNA overnight followed by insulin short-term stimulation (20 minutes). B, [H3]-2-Deoxyglucose uptake assays in cells treated with a scrambled siRNA (Sc). [H3]-2-Deoxyglucose uptake assays were assessed in the same cell passage for GRP78 immunoblots with or without esiRNA. The data from 5 independent experiments in duplicate are expressed as mean fold change with respect to untreated controls (Cn) ± standard error (SE). *P < .05.
Discussion
Polycystic ovary syndrome is characterized by a chronic hyperandrogenism and a higher risk to develop insulin resistance with compensatory hyperinsulinemia, metabolic syndrome, and type 2 diabetes mellitus in the absence of treatment. In this context, the excess of androgens exerts a wide type of molecular alterations in several tissues including the endometrium, leading to an insulin signaling and glucose uptake deregulation. 4,5,7,9 Besides, androgen effects have been related to ER homeostasis, promoting gene expression of ER chaperones and cytoplasmic proteins involved in the ER stress response, as reported in prostate cancer. 16 In this regard, ex vivo and in vitro studies in prostate cancer cells and LNCaP prostate cell line, respectively, show that androgens lead to an increase in the transcriptional activity of genes that codify for ER stress-induced proteins such as NDRG1 and HERPUD1, 17 besides an increase in messenger RNA (mRNA) and protein levels of GRP78. 15 As known, GRP78 is a key chaperone involved in ER protein folding and relieving the ER stress. 17 Nevertheless, it is still unclear how androgens regulate the ER function and glucose uptake in the endometrial tissue, particularly in the endometrial stroma, which maintains the glucose storage for an appropriate endometrial function. 2 To address this, we evaluated ex vivo the protein levels and pattern expression of GRP78 (considered an early ER stress marker) 17 in endometrial samples from patients with PCOS and compared with endometrial tissue from fertile women. Also, we developed an in vitro cell model to evaluate testosterone effects on GRP78 protein levels and glucose uptake in human endometrial stromal cells.
The results of PCOS endometria show a significant decrease in GRP78 protein levels and a diminution in GRP78-positive stained cells in the stromal compartment from patients with PCOS. We attribute these effects to the hyperandrogenemic state of these women in the absence of obesity and insulin resistance, given their normal body mass index and the euglycemic and normoinsulinemic status assessed by different measurements, such as fasting and circulating insulin levels after OGTT, HOMA-IR, and ISI composite. 27,28 Nonetheless, it is useful to compare these findings with endometria from obese and women with PCOS having insulin resistance, where such abnormal conditions could alter insulin-induced cell signaling and probably glucose uptake into the tissue. 4,5,7,8,29
On the other hand, in vitro results showed that testosterone exposure induces a reduction in GRP78 protein content, concomitant with a reduced glucose uptake in the stromal cell model. Our findings are in agreement with the fact that testosterone leads to 50% diminution in glucose uptake in subcutaneous adipocytes from women without PCOS, although GRP78 was not included in that study. 10 Importantly, GRP78 protein has been addressed to stimulate the energetic metabolism in different cell models, as well as the ER stress response. 12,30,31 Therefore, the evidence obtained in the present work highly suggests that GRP78 could be necessary for glucose availability in human endometrial stromal cells and that GRP78 protein content could be downregulated by androgen action. To corroborate these observations, an esiRNA was used against the transcript of GRP78 in T-HESC stimulated with a short-term insulin dose, which leads to a decrease in glucose uptake. Interestingly, such effect was also observed in insulin-untreated cell cultures incubated with GRP78-esiRNA, suggesting a role for GRP78 in the constitutive glucose uptake process in endometrial stromal cells. In addition, androgen activity has been implicated in the regulation of glucose homeostasis in several cell models. For instance, dehydroepiandrosterone or testosterone stimulation induces GLUT1 and GLUT4 translocation to the plasma membrane in human adipocytes and skeletal muscle cells and increases IRS-1 total protein content in T-HESC. 23,32 –34 In contrast, several evidence have associated testosterone as a negative effector of energetic metabolism, such as glucose uptake and insulin signaling, where testosterone stimulation leads to a decrease in mRNA and protein levels of IRS-1 with an increase in its serine phosphorylation in human endometria explants and in mouse skeletal muscle. 35–36 In addition, long exposure to testosterone of 3T3-L1 and C2C12 cell lines reduces the protein levels of IRS-1 and GLUT4. 37 Nevertheless, none of the above-mentioned reports include GRP78 as part of testosterone-regulated glucose uptake mechanism. Besides the role of GRP78 as signaling mediator in ER stress, the present investigation describes a novel regulation of testosterone on GRP78 protein content, which is negatively involved in glucose uptake in immortalized endometrial stromal cells. Furthermore, androgen actions also involve the ER homeostasis regulation by stimulating the expression of calcium-dependent ER resident chaperones (GRP78 included), improving the ER protein folding in human prostate tissue. 16 Unlike these findings, the present study is the first evidence that testosterone could negatively regulate the protein levels of GRP78. Moreover, testosterone was unable to increase the mRNA levels of GRP78 (data not shown). Probably, the discrepancy of data between prostate and endometrial models is due to the different origin of both tissues.
In conclusion, testosterone exposure could deregulate glucose availability by reducing GRP78 protein levels in endometrial stromal cells. Given that most of the patients with PCOS are obese and exhibit biochemical hyperandrogenemia plus hyperinsulinemia, further studies are necessary to know how these metabolic features influence on GRP78 protein levels in the endometrium of these women. Moreover, androgen effects on the glucose uptake could be reflected on the endometrial glucose storage and compromise the normal development of the endometrium through the menstrual cycle. Potentially, these alterations could be detrimental to cell decidualization or for the uterine morphological and molecular adaptation for embryo receptivity and implantation, which could explain in part the higher spontaneous miscarriage rate observed in patients with PCOS.
Footnotes
Acknowledgments
The authors thank the laboratory team for its collaboration and Dr Juan Carlos Vera from University of Concepcion (Chile) for the training in glucose uptake assays. We dedicate this article to Dr Vera, whose sudden death was a great loss for us all. We are also grateful to women who donated tissue.
Authors’ Note
C. Rosas contributed to design and execution of experiments, analysis of data, and manuscript redaction. L. Oróstica contributed to execution of experiments and analysis of data. C. Poblete performed cell cultures. R. Carvajal contributed to the execution of experiments. F. Gabler contributed to analysis and classification of histological samples. C. Romero contributed to analysis of data and discussion of results. S. Lavandero and M. Vega contributed to design of experiments, analysis of data, discussion of results, and manuscript redaction.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by FONDECYT 1130053 (M.V.), FONDAP 15130011 (S.L. and C.M.), and doctoral thesis support (CONICYT 24121256).