
Abstract
While it is generally accepted that tendon healing following widespread extracellular matrix trauma is limited, tenocytes are thought to have the capacity to repair small amounts of microdamage generated through activities of daily living to maintain the mechanobiological function of the tendon. Despite this, few studies have directly studied the mechanisms governing this process. To address this, we developed a tunable ex vivo model of extracellular matrix microdamage in live tendon explants that enables us to track both clearance of denatured collagen and closure of a microsized defect in the tendon matrix. The purpose of this study was to controllably induce varying levels of localized microdamage to the tendon explants and identify (1) if thresholds for repair exist and (2) whether repair mechanisms are dependent on initial damage size. We found that within 3 weeks, all tendon explants were able to clear damaged matrix to some extent regardless of the damage size. Interestingly, larger injuries resulted in a more robust rate of damaged matrix clearance in the later weeks, while smaller injuries exhibited a more consistent rate that led to full clearance in two explants. While all groups demonstrated some closure of the hole over the course of culture, only a small subset of explants with smaller initial hole sizes demonstrated nearly complete closure. Greater than 50% clearance of denatured collagen was typically associated with an accompanying closure of the defect, suggesting a strong relationship between clearance and closure. Overall, our work characterizes a novel laser-induced microdamage model, which will be a powerful asset for investigating mechanisms of damage accumulation and/or repair important to maintaining the mechanobiological function of the tendon, as well as identifying local tendon-specific factors that can be leveraged for therapeutics.
Impact Statement
In this study, we developed a novel laser-based ablation system that produces tunable and reproducible micron-sized damage in live tendon explants. To our knowledge, this is the first direct demonstration that intrinsic cell-mediated responses can be sufficient to initiate tendon ECM repair in response to a localized microdamage injury. Moreover, this model provides a platform for direct visualization and analysis of local tissue repair, offering new opportunities to uncover mechanisms underlying microdamage accumulation and healing that maintain the mechanobiological function of the tendon, as well as to identify local tendon-specific factors that can be leveraged for therapeutics.
Introduction
Resident tendon cells, tenocytes, maintain tissue structure and function by continuously remodeling the extracellular matrix (ECM). Increased mechanical loading from daily activities or exercise induces microdamage in the matrix, which is thought to be repaired by resident cells. This repair is generally considered a multistep process, beginning with proteolytic clearance of damaged ECM followed by deposition and incorporation of new matrix proteins (neomatrix) to restore or improve mechanical function.1–4 Incomplete repair and the progressive accumulation of microdamage over time are the prevailing hypotheses for the development of chronic tendon degeneration. While macroscopic tendon responses to injury and exercise have been extensively studied, comparatively little is known about how tenocytes respond to localized ECM microdamage.
Most insights into tendon microdamage repair have come from models of widespread damage induced by repetitive fatigue loading. The most informative model to date was developed by Fung et al., in which the patellar tendon is clamped and directly loaded in vivo. 5 Using this approach, moderate levels of fatigue loading caused a 20% loss in stiffness and structural matrix damage that was unrecoverable even 8–10 weeks later, 6 implicating a limited repair capacity in the resident tenocytes. 7 At lower levels of fatigue-induced microdamage, tendons upregulated matrix metalloproteinase (MMP) and collagen expression at 7 days postinjury, suggestive of matrix turnover. 8 However, it remains unclear whether these responses reflect true microdamage repair or adaptation to loading. More recently, in vitro models have probed innate tenocyte responses to induced microdamage.9,10 These studies showed that tenocytes isolated from the systemic injury response mounted robust inflammatory and matrix degradation responses, yet were unable to restore matrix damage. The widespread damage used in these models may be too extensive to simulate an effective local response. This underscores the need for complementary models to study tenocyte-mediated ECM repair in the context of small-scale, localized microdamage.
Laser ablation offers a promising method to induce highly localized matrix damage in tendon. A focused laser pulse delivers a high amount of energy to a confined region generating micron-scale lesions whose size can be tuned by adjusting the focusing angle and pulse energy. 11 The resulting defects comprise both a discrete void and surrounding microdamage in the form of denatured ECM proteins, making this approach particularly well-suited to probe cellular responses to localized matrix damage and altered mechanical environments. Indeed, recent studies have applied laser ablation in engineered fibroblast-containing microtissues, revealing new insights into the cellular mechanisms that mediate damaged ECM clearance.12,13 While powerful, these microtissues do not fully recapitulate the complexity of native tendon ECM or its resident cell populations.
To overcome these limitations, we established a laser ablation approach in live tendon explants. Explants preserve the native ECM architecture as well as the cell–cell and cell–ECM interactions, enabling study of intrinsic repair processes in a physiologically relevant context. Our objective was to develop a laser ablation technique to reliably induce tunable levels of microdamage in live tendon explants and to use this model to characterize local repair responses. We hypothesized that resident tendon cells would be capable of repairing and remodeling microdamage in situ and that resident repair capacity would be dependent on the extent of damage.
Methods
Design of laser ablation bioreactor
Our novel technique required the design and fabrication of a custom bioreactor to ablate and culture whole tendon explants while maintaining physiological loading conditions. Our system was designed with a glass window at the bottom of the culture wells to visualize explants during ablation (Fig. 1A). The windows include etched lines to track the position of the ablated hole in reference to the grip edge. This ensured that the correct region was analyzed, even if the initial damage was fully repaired. Inclusion of an in-line load cell and a linear variable differential transformer enables precise and controlled cyclic loading for future studies. 14 The ability to ablate the explants directly in the bioreactor reduces excess handling and time outside of culture.

The ablation bioreactor system.
Sample preparation
Flexor digitorum longus (FDL) tendons were harvested from young (2–4 months) C57BL/6J male mice following sacrifice (BU IACUC PROTO202000046). Following previously described methods, 15 all explants were washed in 1× PBS supplemented with antibiotics (100 units/mL penicillin G, 100 µg/mL streptomycin (Fisher Scientific, Waltham, MA), and 0.25 µg/mL Amphotericin B (Sigma-Aldrich)). Explants were then immediately loaded into bioreactor grips using a loading rig to ensure all tendons were reliably held at a 10-mm gauge length. The tendons were oriented such that the joint-facing side of the tendon was ablated. The gripped tendons were then placed into our custom bioreactor and stretched manually to approximately 1% strain, which has been shown previously to be adequate for maintenance of tissue properties. 14 Left and right tendons from every mouse were used to conserve animal numbers, but tendons from the same animal were not attributed to the same experimental group. Throughout ablation and culture, explants were maintained in standard culture medium consisting of low glucose Dulbecco’s Modified Eagle’s Media (1 g/L; Fisher Scientific) supplemented with 10% fetal bovine serum (Cytiva, Marlborough, MA) and antibiotics.
Ablation
The bioreactor was transported to the stage of a Zeiss Axiovert S100 microscope (Zeiss, Oberkochen, Germany) equipped with a 10× objective and a 1064 nm q-switched pulsed-nanosecond Nd:YAG laser (Minilite, Continuum, San Jose, California). As previously described, 12 the beam is enlarged by a beam expander (Edmund Optics #39–739, Barrington, New Jersey) from 3 to 9 mm, directed through a dichroic mirror suitable for 1064 nm, and focused into the tissue through a 10× objective. The laser energy was measured directly after the beam expander and was adjusted to the desired laser power: 2, 5, 10, 15, or 25 mJ. All tendons were ablated near the bifurcation of the two fascicles of the FDL, in the middle of the larger fascicle. To ensure a uniform depth of the laser focal point, the microscope was focused on the outer edge of the large fascicle. Phase images were taken with a 20 Mp Blackfly camera (FLIR, Wilsonville, Oregon) before and immediately following microdamage injury (Fig. 1D).
A nonablated control injury (“puncture”) was also developed to account for differences in repair capacity attributed to heat damage generated by the laser. A 32-gauge needle was pushed through the large fascicle of the explant at a similar location along the length of the tendon as the ablations. The 32-gauge puncture produces a hole defect comparable to the 5 mJ laser pulse without producing heat-denatured collagen in the process.
Cell viability
Cell viability was assessed through live/dead staining in 1× PBS containing fluorescein diacetate (FDA; 4 mg/mL; Fisher Scientific) and NucBlue nuclear stain (2 drops/mL; Thermo Fisher Scientific, Waltham, Massachusetts). Dead cells were identified as positive for NucBlue staining but negative for FDA staining. To quantify, a circular region of interest (ROI) was drawn containing all dead cells around the injury. The diameter of that circular ROI was used to compare the extent of cell death generated by the laser ablation.
Quantification of damaged ECM and clearance
The denaturation of collagen by the thermal energy released in the tissue during ablation allows the damage generated to be stained via collagen hybridizing peptide (CHP). Therefore, collagen damage and cell location were stained by Cy3-CHP (Advanced Biomatrix, Carlsbad, California) and NucBlue, respectively. Briefly, a 200 μL CHP solution of 4.32 μg/mL CHP was heat shocked at 80°C for 10 min and then cooled in a water bath for 1.5 min. After cooling, tendons were added to the solution and stained at 4°C for 18 h. Tendons were then removed from the CHP stain and transferred to a NucBlue stain containing 2 drops of NucBlue stock in 1 mL of PBS for 5 min. Confocal image z-stacks of the ablation site were captured on an Olympus FV3000 with a 20× objective.
Custom software was used to quantify damage intensity and dimensions, as well as the number and area of cell nuclei in the damaged region. First, the background signal was removed through user-defined thresholding of the stack of CHP images. The max projection image of the image stack was then used to identify an ROI. The ROI was defined by the code as the pixels that fell within CHP stain regions verified by user input. The depth of the denatured collagen was assessed by identifying the number of slices containing CHP stain in the ROI. The width and the length of the denatured collagen ROI were obtained by taking the distance between ROI edges in the direction perpendicular and parallel to the long axis of the tendon, respectively. The volume of denatured collagen was determined by quantifying the area of CHP stain in the ROI on each plane of the image stack and multiplying by the image slice height of 3.94 μm. Average collagen damage intensity was determined by summing the intensity of each pixel of the ROI across all slices, including pixels without stain, and taking the average of the summed intensity of all the pixels in the ROI. For studies tracking changes in microdamage over time, clearance is presented as the percent difference from initial injury (day 0). Positive values represent clearance of denatured collagen or closure of the matrix defect when compared to the initial day 0 mean value of the metric. Negative values represent accumulation of denatured collagen or expansion of the matrix defect when compared to the initial day 0 mean value of the metric. The daily rate of clearance is presented as the change from the previous time point divided by seven days. A positive rate shows faster clearance or repair, and a negative rate represents slower clearance or an accumulation of damage.
Quantification of hole morphology and closure
The mechanical failure of the surrounding ECM and destructive tissue ablation leaves a hole in the tendon ECM that is detectable via second harmonic generation (SHG) imaging. Tendons were imaged for SHG using a two-photon microscope with the laser tuned to 1190 nm, a dwell time of 1.2 ms, and an imaging window of 1024 × 1024 pixels. Pockels were interpolated exponentially from 130 at the tendon surface to 180 at 150 µm into the tendon, with frame-averaging every two frames. Custom software was then used to quantify the volume of the hole left in the tendon ECM. Briefly, a Gaussian filter was applied to the SHG images to reduce noise, and the images were auto-thresholded using a locally adaptive threshold with high sensitivity. The user then selected the frame where the ECM hole first begins, the frame where clear collagen fibers disappear, and the position of the ECM hole. The software then tracks the hole across the z-stack and calculates the number of pixels without an SHG signal. Hole volume is calculated by quantifying the area of continuous dark pixels on each plane of the image stack and multiplying by the image slice height. For studies tracking changes in hole size over time, closure is presented as the percent difference from initial injury (day 0).
Birefringence imaging
Local collagen structure was assessed at 0 and 21 days postinjury. After the explants were assessed for clearance and closure using whole-mount imaging, explants were frozen at minus 20°C until they could be embedded for sectioning. Explants were embedded and frozen in optimal cutting temperature compound (Fisher Scientific), and 10 µm thick serial sections were collected on glass slides. After sectioning, one section containing clear damage from the ablation injury was selected from each sample. The selected sections were then rinsed in 1× PBS for 5 min and fixed in 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA) for 10 min at room temperature. Following fixation, the sections were rinsed in 1× PBS for 5 min and mounted in 60% glycerol.
The slides were imaged using a custom-built quantitative birefringence microscope in a configuration that is an inverted version of the system described in Blanke et al.
16
The tendon sections were imaged at a magnification of 20×, at 9 different evenly spaced orientations of the linear polarizer, and the method described in Glazer et al. was used to extract the relative retardance and optic axis orientation from these measurements,
17
and the redundancy present in this method was eliminated. With this method, it is assumed that Jones calculus can be used to model the sample at each pixel imaged as a variable retarder with unknown retardance, sin(δ), and optic axis, φ. The resulting equation models the change in intensity as follows:
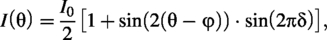
MMP activity
To understand how the resident cells were facilitating the removal of denatured collagen from the injury site, we assessed MMP (1, 2, 3, 7, 8, 9, 10, 13, and 14) activity via analysis of spent culture medium (n = 2–6/group/time point) using a commercially available Förster Resonance Energy Transfer-based generic MMP cleavage kit (SensoLyte 520 Generic MMP Activity Kit Fluorimetric, Anaspec, Fremont, CA). Media was collected during standard media changes on days 6, 14, and 20 of culture. MMP activity is represented as the concentration of MMP cleaved product (5-FAM-Pro-Leu-OH).
Data and statistics
All data are presented as violin plots with the median, first quartile, and third quartile marked. Data points more than two standard deviations outside of the mean were removed as outliers. Statistical evaluation was performed in GraphPad Prism 8 (GraphPad, San Diego, CA) using individual one-way analysis of variances (ANOVAs) for each laser power level with effects for time in culture. Bonferroni corrected post hoc t tests were then used to identify differences within each time point where appropriate. Bartlett’s test for heteroscedasticity and D’Agostino-Pearson omnibus test for normality were used to assess the data sets. In instances where a data set was heteroscedastic, a Brown–Forsythe ANOVA and Dunnett’s T3 multiple comparisons test were used. In instances where a data set was not normally distributed, a Kruskal–Wallis test and Dunn’s multiple comparisons test were used. A significant increase in the rate of clearance was determined through a one-tailed t test of the timepoint compared to a hypothesized zero rate of clearance. For all comparisons, significance was noted at *p < 0.05. Correlations between the denatured ECM cleared and the ECM hole closure were assessed using simple linear regression analysis.
Results
We first established that laser ablation could reproducibly generate tendon injuries of varying sizes by modulating pulse energy. Consistent with prior studies, 11 higher pulse energies produced proportionally larger regions of cell death and collagen denaturation (Fig. 2A,B). This increase in denatured collagen volume is primarily attributable to greater damage width (transverse dimension of the tendon) and depth (Supplementary Fig. S1), while damage length along the tendon axis remained largely unchanged. Correspondingly, the volume of the ECM defect, quantified by the hole size via SHG, also scaled with laser power (Fig. 2C).

Characterization of the damage generated by the ablation system
Having validated the ability to generate graded injury sizes, we next asked how the extent of initial injury size affects intrinsic repair response. To this end, tendon explants were ablated at three distinct laser powers (2, 5, or 10 mJ). Representative images show a progressive reduction in CHP staining around the ablation site over time across all groups, indicating clearance of denatured collagen (Fig. 3A). Quantitative analysis confirmed that collagen clearance increased with culture duration; however, significant reductions from baseline were observed only in the 5 mJ and 10 mJ groups at day 21 (Fig. 3B). Remarkably, two tendon explants in the 2 mJ group achieved complete clearance of denatured collagen, with one clearing by day 14 of culture and the other by day 21.
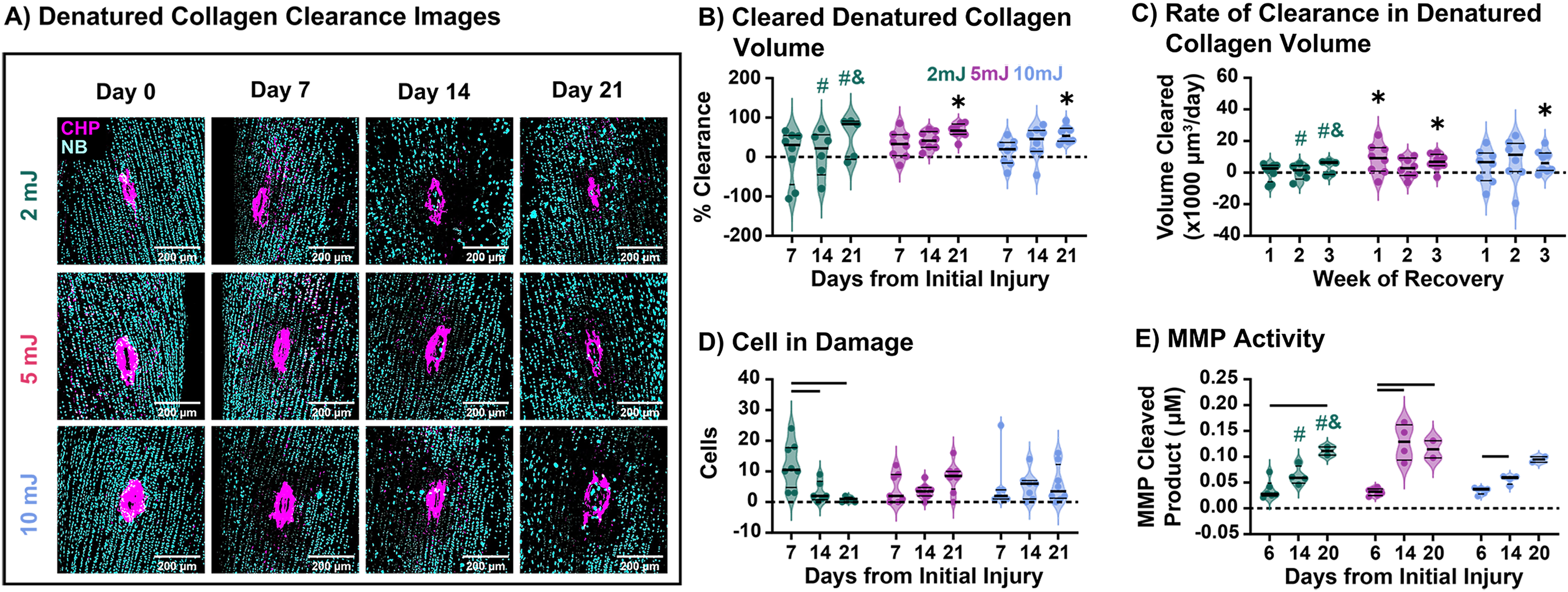
We then asked whether cells were recruited to participate in the clearance process or whether clearance was a passive process. No significant clearance occurred in explants from frozen tendons, suggesting a cell-mediated process (data not shown). Indeed, all live groups exhibited a low level of cellularity at the injury site (Fig. 3D), but the timing of cell appearance was dependent on initial damage. While small damage sites (2 mJ) showed a progressive reduction in cell number at the site over time, larger sites exhibited slight increases over culture. This could indicate that early cell recruitment is important for efficient clearance. Looking at how these cells clear the tissue, we found that MMP activity increased over the culture period in all groups (Fig. 3E). However, this did not show time-dependence, suggesting other potential proteolytic mechanisms at play.
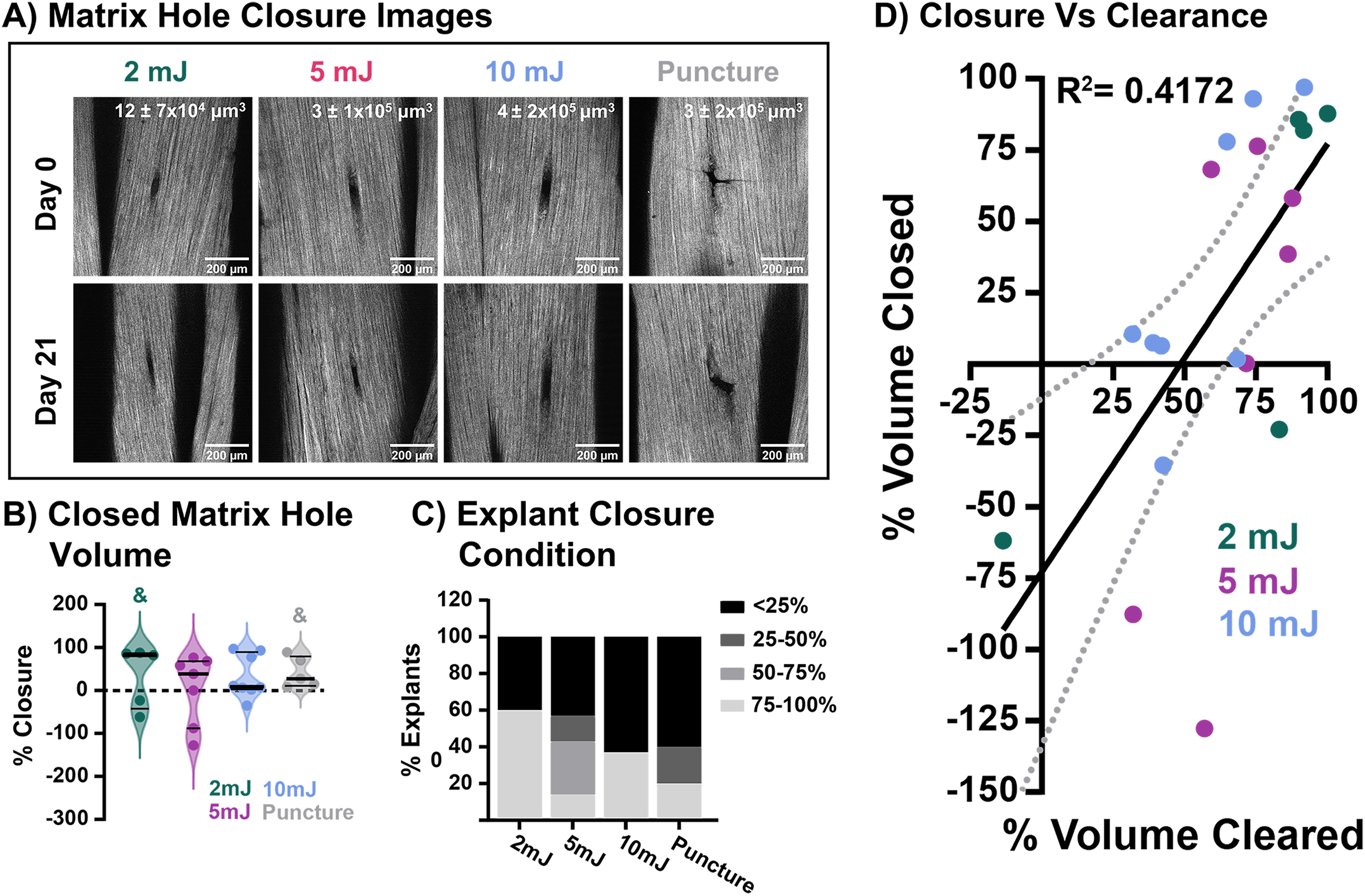
Following clearance of damaged collagen, cells then facilitate the deposition and replacement of the missing collagen matrix. The initial hole in the collagen matrix due to laser pulse or puncture injury, as well as any potential cell-mediated repair or infill after 21 days of culture, was visualized through SHG imaging (Fig. 4A). No group showed significant closure from their respective initial hole volume (Fig. 4B). However, categorizing the tendon explants by percent closure, it becomes clear that as initial injury size increases, fewer explants can initiate closure (Fig. 4C). Notably, explants in the 2 mJ and 10 mJ groups either had nearly complete closure (above 75% reduction of the initial hole volume) or had not closed (below 25% reduction) and, in some cases, had increased above the initial hole volume. Looking at the relationship between damage clearance and hole closure, the amount of defect closure in the explant is moderately correlated with the amount of denatured collagen (R2 = 0.4172; Fig. 4D). While no group exhibited significant closure of the injury site, there was no further degeneration of the tissue over the culture period. Birefringence imaging, a form of polarized light microscopy, demonstrated groups were able to maintain the organized collagen structure surrounding the injury site (Supplementary Fig. S2).
Discussion
In this study, we developed a novel laser ablation bioreactor system capable of producing tunable and reproducible micron-sized damage in live tendon explants. By varying pulse energy, this system provides a customizable platform to study cell responses to localized microdamage. Although the timing and rate of denatured collagen clearance varied based on the size of injury, the extent and ability to clear denatured collagen was consistent across groups. These findings suggest that while the kinetics of repair may depend on the initial severity of injury, the capacity of clearance is conserved. Importantly, the presence of cells within the bounds of the injury at all damage levels suggests that clearance of acute microdamage is an intrinsic, cell-mediated process.
The cellular origin of this repair response remains to be determined. It is possible that resident immune cell populations, such as resident macrophage-like cells, contribute to clearance. 18 However, given their low abundance, it is equally plausible that tenocytes migrate from surrounding regions of the explant in response to injury-induced inflammatory signaling. 19 Indeed, acute tendon injuries are known to trigger a proinflammatory environment characterized by the release of cytokines such as interleukin-1β, tumor necrosis factor-α, and interleukin-6 and the recruitment of immune cells within 2–3 days. 20 Ongoing studies will quantify the levels of secreted proinflammatory cytokines near the injury site. Regardless of their origin, our results establish that cells near the microdamage site play a central role in the clearance of denatured collagen.
Cells employ a variety of molecular mechanisms for ECM clearance, including phagocytosis and direct enzymatic degradation via MMPs, serine proteases, and cystine proteases.21,22 Our data provides compelling evidence that MMP activity contributes to matrix clearance of local ECM damage. It is important to note that the expression of MMPs responsible for clearing denatured collagen from the damage area is known to be affected by mechanical loading.9,23–25 The injuries produced lead to the removal of ECM and unloading of collagen fibrils and fibers in the tissue, which is known to increase MMP production in tendon.15,24,26,27 However, there are potential clearance mechanisms outside of MMPs that have yet to be explored. For example, previous studies have shown that fibroblasts have the capacity to recycle collagen through an endocytic process and that removal of denatured ECM from an injury site relies on ROCK and dynamin activity, suggesting a phagocytic or endocytic process.12,28 Future studies will seek to more deeply characterize the local molecular clearance mechanisms and how the cells are driving denatured collagen clearance.
Following clearance of damaged collagen, cells facilitate the deposition and replacement of the missing collagen matrix. Previous studies support tenocyte-produced neomatrix during healing, including collagen III and collagen I, 29 both of which can be visualized using SHG imaging. 30 The puncture group closed the ECM hole to a similar extent as the laser injury groups, indicating that the presence of denatured collagen does not affect ECM hole repair. With more viable cells near the injury site, the explants with smaller injuries may have been able to start laying down new neomatrix and contract existing matrix sooner, supported by the observed decrease in initiation of closure with increased injury size. Alternatively, the closure exhibited may be due to a contraction of the surrounding matrix in response to changes in the local mechanical environment. Resident tendon cells have been shown to be able to drastically contract the ECM in response to changes in mechanical load through an α-smooth muscle actin-mediated cellular mechanism. 31 Future studies will aim to identify the presence of contractile cells and the deposition of neomatrix proteins like collagen III and fibronectin. Interestingly, correlations demonstrate that clearance and closure are connected. This would suggest that before the defect can be contracted or new collagen matrix can be deposited at the injury site, the injury site must be debrided of damaged ECM.
This study and model system are not without their limitations. While thermal denaturation is not physiological, this model is more controllable and allows for quantification of damage over time. In addition, microdamage generated by the laser ablation is comparable in size to that generated during overload (∼0.47% of total surface area in laser ablation compared to as high as 13% of the total tendon area for overload).32,33 The average size of “microtears” in tendons can range from 0.33% to 3% in loaded tendons to 0.44% to 2.66% in unloaded tendons. 34 We were also unable to track the same injury across all time points in this study, adding variability to our measurements. To address this, we are currently building a longitudinal system to enable real-time tracking of cell behavior, damage clearance, and neomatrix deposition. Finally, we used mouse tendon explants to preserve intact tissue architecture and the full complement of resident cells. We acknowledge that species differences may limit direct quantitative extrapolation to human tendons. Therefore, our conclusions emphasize mechanistic principles of intrinsic, cell-mediated clearance and defect closure, rather than direct equivalence across species. Future work should validate these mechanisms in vivo and in human samples if possible.
We present here a novel model for studying innate tissue healing processes following a microscale injury without interference from extrinsic factors. To our knowledge, our model system has, for the first time, provided the ability to directly identify that intrinsic cell-mediated responses may be sufficient to initiate ECM repair in response to localized microdamage injury. We have identified that larger levels of microdamage affect the microenvironment enough to alter the timing of matrix repair, with delayed increases in the rate of damage clearance. We have also shown that closure of a microdamage hole defect may rely on the extent of damage clearance. Nevertheless, our novel laser ablation-induced microdamage model enables the direct visualization and study of local tissue repair, which will be a powerful asset for understanding mechanisms of microdamage accumulation and/or healing important to maintain the mechanobiological function of the tendon, as well as identifying local tendon-specific factors that can be leveraged for therapeutics.
Authors’ Contributions
A.N.A. and M.T.L. have contributed to all aspects of this study, including research design, data acquisition, interpretation/analysis of data, and drafting/revision of the article. M.B. and I.J.B. have contributed to data acquisition, interpretation/analysis of data, and drafting/revision of the article. J.E. and B.K.C. have contributed significantly to research design, interpretation/analysis of data, and drafting/revision of the article. All authors have read and approved the final submitted article.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by NIH/NIA
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.