
Abstract
Glypican-3 (GPC3) belongs to the glypican family of heparan sulfate proteoglycans and is frequently overexpressed in hepatocellular carcinoma (HCC). The overexpression of GPC3 is associated with the poor clinical outcomes, suggesting its potential as a clinically relevant biomarker and therapeutic target. Therefore, anti-GPC3 monoclonal antibodies (mAbs) have been developed in various modalities for tumor diagnosis and therapy. In this study, 88 clones of novel anti-GPC3 mAbs were established using a flow cytometry-based high-throughput screening, the Cell-Based Immunization and Screening (CBIS) method. Among them, a clone G3Mab-25 (IgG1, κ) recognized GPC3-overexpressed Chinese hamster ovary-K1 (CHO/GPC3) but not parental CHO-K1 in flow cytometry. Furthermore, G3Mab-25 recognizes endogenous GPC3 in GPC3-expressing HCC cell lines, including HepG2, HuH-7, and JHH-5. G3Mab-25 specifically recognized only CHO/GPC3, but not other GPC family-overexpressed CHO-K1. The dissociation constant values of G3Mab-25 for CHO/GPC3, HepG2, HuH-7, and JHH-5 were determined to be 1.8 × 10−8 M, 7.3 × 10−9 M, 3.9 × 10−9 M, and 1.4 × 10−9 M, respectively. Moreover, G3Mab-25 detects the N-terminal fragment of GPC3 in western blotting. In immunohistochemistry, G3Mab-25 showed a diverse staining pattern for GPC3 in HCC tissues. G3Mab-25, established by the CBIS method, is a versatile mAb for basic research and is expected to contribute to tumor diagnosis and therapy.
Keywords
Introduction
Glypican-3 (GPC3) belongs to the glypican family of heparan sulfate (HS) proteoglycans and is tethered to the plasma membrane through a glycosylphosphatidylinositol (GPI) anchor.1–3 GPC3 is a ∼70-kDa protein that undergoes endoproteolytic cleavage by Furin at the Arg358–Ser359 position, generating a 40-kDa N-terminal fragment and a 30- kDa C-terminal fragment bearing HS–type glycosaminoglycan side chains.4–6 The N- and C- subunits are covalently linked via disulfide bonds formed by highly conserved cysteine residues. 7 The mature glypican can be released from the cell surface through cleavage of the GPI anchor by the lipase Notum.2,3
GPC3 expression is tightly regulated in a stage- and tissue-specific manner during development, with high transcript levels detected in fetal liver, lung, and kidney, but absent in most adult tissues.3,8,9 GPC3 modulates embryonic development by regulating key signaling cascades such as Hedgehog (HH), Wnt, fibroblast growth factor, and bone morphogenetic protein pathways.9–14 In adult tissues, nonpathological re-expression of GPC3 has been reported in regenerating liver following partial hepatectomy, where it promotes parenchymal regeneration through activation of the HH signaling pathway.15,16 Furthermore, GPC3 is frequently overexpressed in hepatocellular carcinoma (HCC). The overexpression is associated with undifferentiated, more aggressive HCC, and poor clinical outcomes, suggesting its potential as a clinically relevant biomarker and therapeutic target in HCC. 17
Soluble GPC3 (sGPC3), generated by enzymatic cleavage of GPC-3, is detectable in the serum of HCC patients but undetectable in that of hepatitis patients and healthy individuals,18–20 making it a valuable diagnostic marker in HCC.21,22 Using monoclonal antibodies (mAbs) against the N-terminal fragment of GPC3, sGPC3 was shown to be superior to α-fetoprotein in sensitivity, and the combination measurement increased overall sensitivity in well- or moderately-differentiated HCC, suggesting that sGPC3 is a serological marker essential for the early detection of HCC. 18
Codrituzumab (derived from mouse anti-GPC3 mAb clone GC33) recognizes the C-terminal fragment (amino acids 524–563) of GPC3. 23 Codrituzumab induced tumor cell death through antibody-dependent cellular cytotoxicity (ADCC)23,24 and exhibited antitumor efficacy in preclinical human tumor xenograft models. 25 Furthermore, codrituzumab was evaluated in phase I and II clinical trials for HCC. Despite demonstrating favorable tolerability, these studies did not reveal significant therapeutic efficacy.26,27 Codrituzumab was further investigated in phase I clinical trials for pediatric solid tumors, including hepatoblastoma, where it has shown good tolerability with minimal toxicity. 28 However, the monotherapy targeting GPC3 is unlikely to achieve sufficient antitumor efficacy. 29
Biodistribution, tumor targeting, and pharmacokinetics of radiolabeled codrituzumab with iodine-124 (124I) were assessed using positron emission tomography (PET) imaging, which demonstrated favorable pharmacokinetics and tumor uptake in HCC patients.30,31 For radioimmunotherapy, GC33 conjugated with the α-emitting radionuclide actinium-225 (225Ac) through the chelator macropa has been evaluated in a preclinical study. 32
GC33 has been developed as a chimeric antigen receptor (CAR) T cell therapy. 33 However, the therapeutic efficacy of CAR T cells in patients with solid tumors has been constrained by the immunosuppressive nature of the tumor microenvironment (TME).34,35 The TME not only provides inhibitory signals that attenuate antitumor immune responses but also lacks essential supportive elements, such as cytokines—particularly interleukin-15 (IL-15)—that are critical for the persistence and optimal functionality of tumor-specific T cells. Recently, GC33-based CAR T coexpressing IL-15 increases the expansion, survival in tumors, and antitumor activity in patients with liver cancers. 36 Therefore, the development of a new anti-GPC3 mAb is essential for tumor diagnosis and therapy.
The Cell-Based Immunization and Screening (CBIS) method includes immunizing antigen-overexpressed cells and flow cytometry-mediated high-throughput screening. Using the CBIS method, we have developed various mAbs against membrane proteins including receptor tyrosine kinases,37,38 and chemokine receptors.39,40 MAbs obtained by the CBIS method possess a variety of epitopes, including conformational epitopes, linear epitopes, and glyco-epitopes, and are suitable for flow cytometry. Furthermore, some of these mAbs are also suitable for western blotting and immunohistochemistry (IHC). This study employed the CBIS method to develop highly versatile anti-GPC3 mAbs.
Materials and Methods
Cell lines
Mouse myeloma P3X63Ag8U.1 (P3U1), Chinese hamster ovary (CHO)-K1, and human glioblastoma LN229 were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). Human HCC cell lines, HepG2 and HuH-7, were obtained from the Cell Resource Center at Tohoku University (Miyagi, Japan). Another HCC cell line, JHH-5, was obtained from the Japanese Collection of Research Bioresources (Osaka, Japan).
JHH-5 was maintained in Williams’ E medium (Sigma-Aldrich Corp., St. Louis, MO, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc., Kyoto, Japan). LN229, HepG2, HuH-7, CHO-K1, and P3U1 were maintained as described previously. 41
Establishment of stable transfectants
The expression vectors of GPC3v2 (designated GPC3) and other GPC members were reported previously. 42 The plasmids were transfected into CHO-K1 and LN229, and stable transfectants were established by sorting with an anti-GPC3 mAb (clone SP86; Abcam, Cambridge, UK), an anti-GPC1 mAb (clone 1019718; R&D systems Inc., Minneapolis, MN, USA), an anti-GPC2 mAb (clone CT3; Cell Signaling Technology, Inc., Danvers, MA, USA), an anti-GPC4 mAb (clone A21050B; BioLegend, San Diego, CA, USA), an anti-GPC5 mAb (clone 297716; R&D Systems Inc.), and an anti-PA16 tag mAb (clone NZ-1 for PA16-tagged GPC6). 42 The GPCs-overexpressed CHO-K1 (e.g., CHO/GPC3) and GPC3-overexpressed LN229 (LN229/GPC3) were finally established.
Production of hybridomas
Animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001). They were carried out following the NIH (National Research Council) Guide for the Care and Use of Laboratory Animals. Two female BALB/cAJcl mice (CLEA Japan, Tokyo, Japan) were intraperitoneally immunized with LN229/GPC3 cells (1 × 108 cells/mouse, five times), and the hybridomas were generated as described previously. 38
Flow cytometry and determination of dissociation constant values
The cells were treated with mAbs in blocking buffer (0.1% bovine serum albumin in phosphate-buffered saline) for 30 minutes at 4°C. The cells were treated with anti-mouse IgG or anti-rabbit IgG conjugated with Alexa Fluor 488 (2000-fold dilution, Cell Signaling Technology, Inc., Danvers, MA, USA). The data were collected and analyzed as described previously. 38
The cells were treated with serially diluted G3Mab-25. Subsequently, the cells were treated with anti-mouse IgG conjugated with Alexa Fluor 488 (200-fold dilution) for 30 minutes at 4°C. The dissociation constant (KD) values were determined as described previously. 38
Western blotting
Western blotting was performed using G3Mabs (1 μg/mL), SP86 (1 μg/mL), or an anti-isocitrate dehydrogenase 1 (IDH1) mAb (RcMab-1 43 ) (1 μg/mL) as described previously. 38
Immunohistochemistry
All procedures of IHC were performed using VENTANA BenchMark ULTRA PLUS (Roche Diagnostics, Indianapolis, IN, USA). Cells were fixed with 4% paraformaldehyde, and the cell blocks were prepared as described previously. 38 The formalin-fixed paraffin-embedded (FFPE) cell sections were stained with G3Mab-25 (0.5 μg/mL) or SP86 (0.5 μg/mL) using the ultraView Universal DAB Detection Kit (Roche Diagnostics and BenchMark ULTRA PLUS.
Solid tumor screen panel (CC00-10-001, Cybrdi, Rockville, MD, USA) and liver tumor tissue arrays (BS03013b, T032d, LV242a, and LV243, US Biomax Inc., Rockville, MD, USA) were stained with G3Mab-25 (2 μg/mL) or SP86 (2 μg/mL) as described above. Staining intensity, pattern, and cumulative GPC3 expression scores (0–7) were determined as described previously.27,36
Results
Development of anti-GPC3 mAbs
To develop anti-GPC3 mAbs, LN229/GPC3 was used as an antigen and immunized in two female BALB/cAJcl mice (Fig. 1A). Hybridomas were generated by fusing LN229/GPC3-immunized splenocytes with mouse myeloma P3U1. After forming colonies, the CHO/GPC3-positive and CHO-K1-negative supernatants were screened. (Fig. 1B). Subsequently, anti-GPC3 mAb-producing hybridomas were cloned by limiting dilution. We obtained 88 anti-GPC3 mAb-producing clones, and their supernatants were further screened for several applications, including western blotting and IHC using the CHO/GPC3 block. Finally, a clone, G3Mab-25 (IgG1, κ), was shown to be applicable to these applications (Fig. 1C).
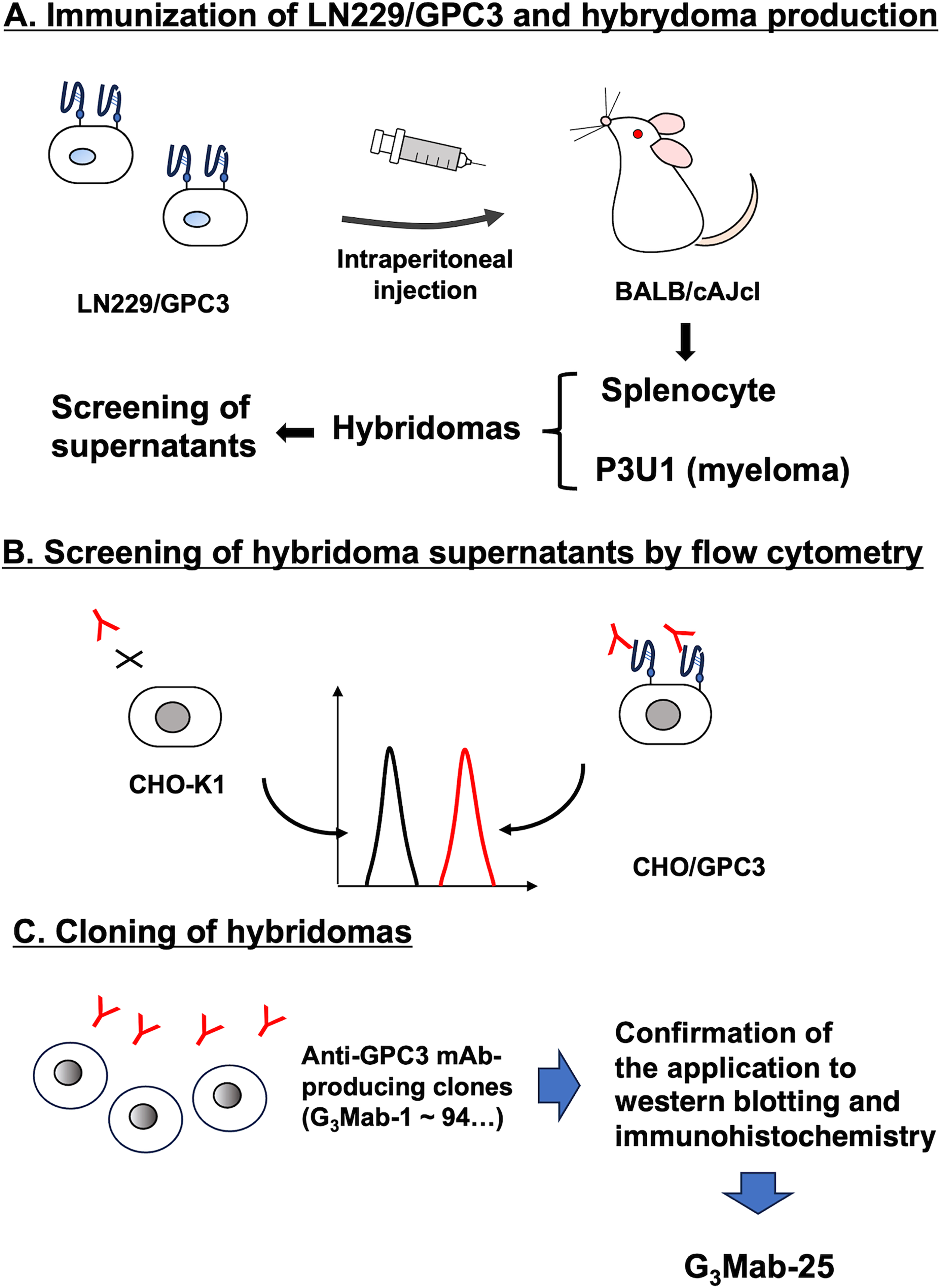
Schematic representation of anti-GPC3 mAbs production.
Flow cytometry using anti-GPC3 mAbs
We purified G3Mab-25 and performed flow cytometry using G3Mab-25 and a commercially available anti-GPC3 mAb (clone SP86) on CHO/GPC3 and CHO-K1 cells. The G3Mab-25 and SP86 recognized CHO/GPC3 in dose-dependent manner from 10 to 0.01 μg/mL (Fig. 2A). Still, they did not recognize CHO-K1 even at 10 μg/mL (Fig. 2B). SP86 showed the similar reactivity at 10 and 1 μg/mL but relatively lower reactivity at 0.1 and 0.01 μg/mL compared to G3Mab-25 (Fig. 2A). We next investigated the reactivity of G3Mab-25 against an endogenous GPC3-expressing HCC cell lines, HepG2, HuH-7, and JHH-5. G3Mab-25 and SP86 exhibited dose-dependent reactivity of HepG2, HuH-7, and JHH-5 (Fig. 3). These results indicate that G3Mab-25 recognizes GPC3 in flow cytometry.
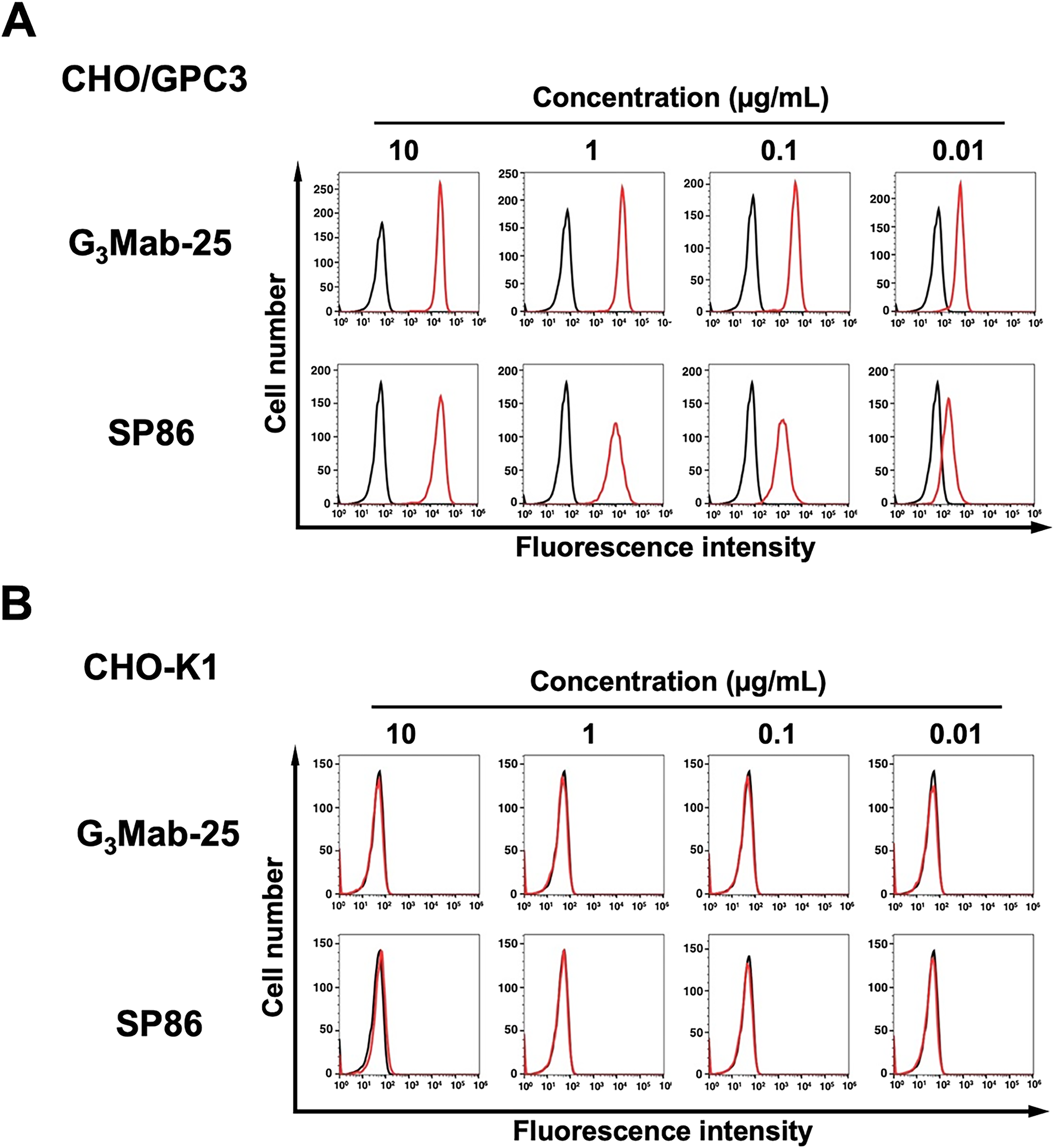
Flow cytometry analysis of G3Mab-25 and SP86 against CHO/GPC3 and CHO-K1. CHO/GPC3
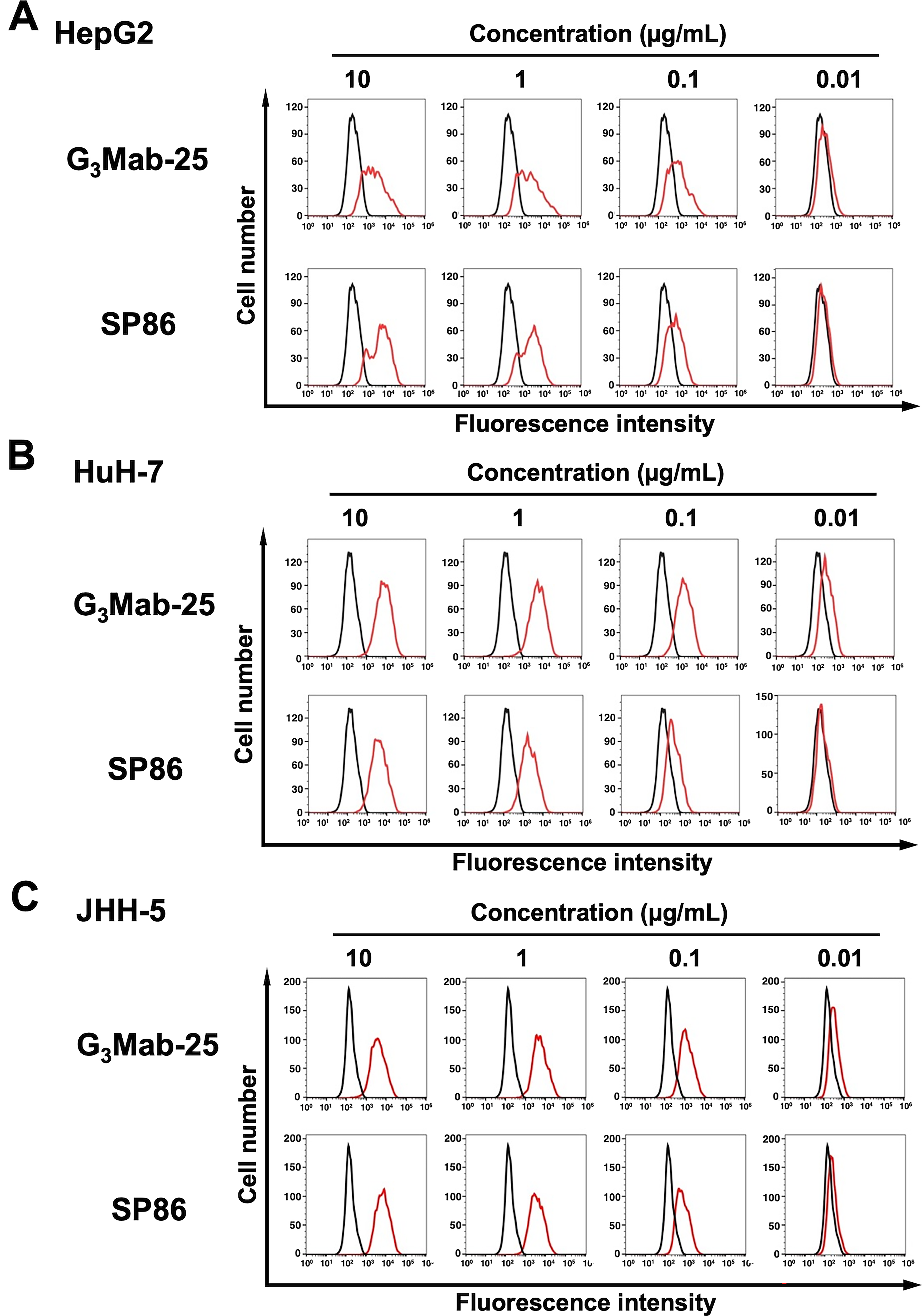
Flow cytometry analysis of G3Mab-25 and SP86 against HCC cell lines. HepG2
Specificity of G3Mab-25 against GPC family members
Using GPC1 ∼ GPC6-overexpressed CHO-K1, the specificity of G3Mab-25 was investigated. As shown in Figure 4, G3Mab-25 recognized CHO/GPC3 and did not react with other GPC-overexpressed CHO-K1. These results indicate that G3Mab-25 is a specific mAb against GPC3 within the GPC family.
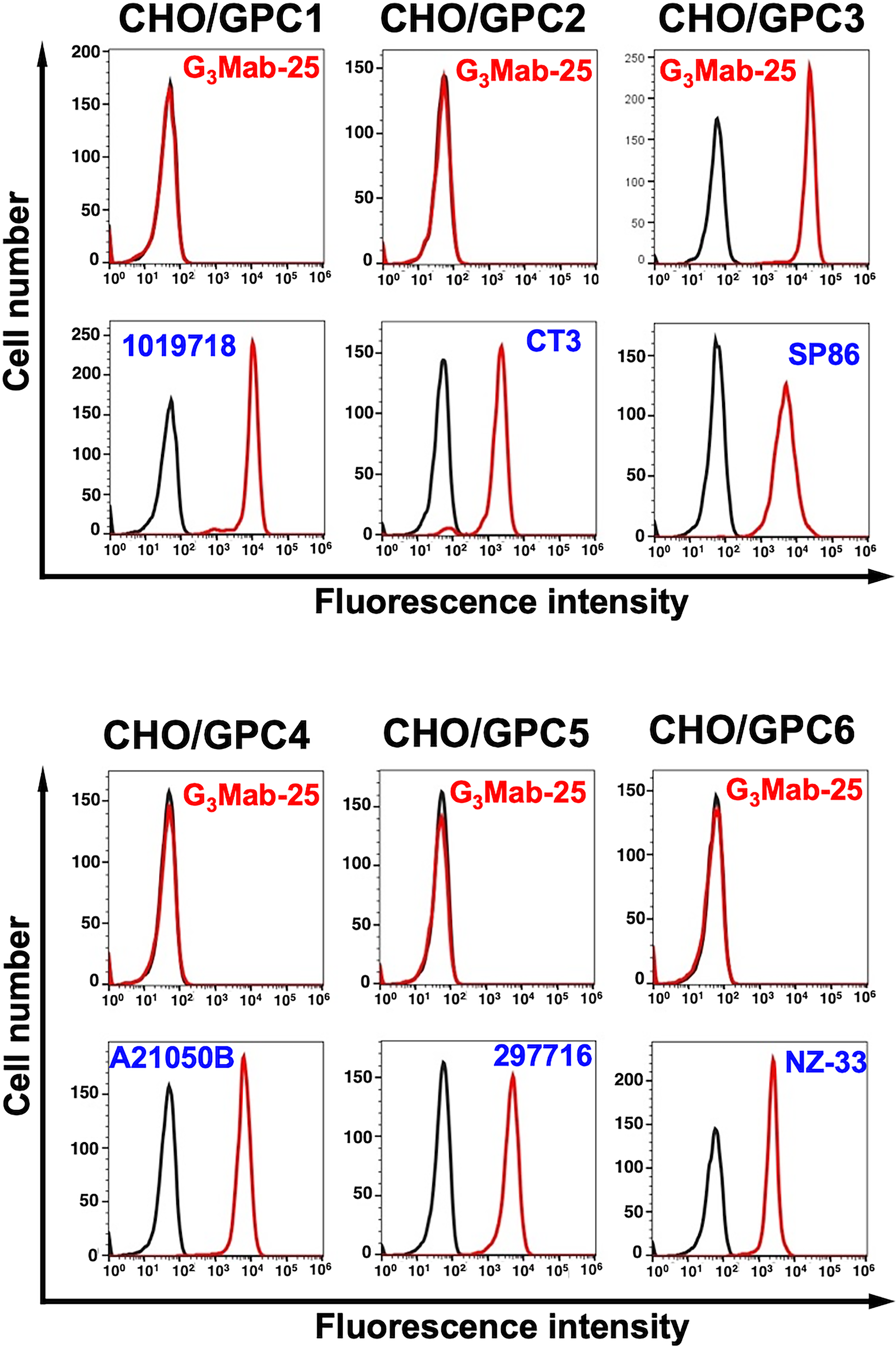
Flow cytometry analysis of G3Mab-25 in GPC family members-overexpressed CHO-K1. GPC-overexpressing CHO-K1 cells (GPC1, GPC2, GPC3, GPC4, GPC5, and PA16-GPC6) were treated with 1 µg/mL of G3Mab-25 (red) or control blocking buffer (black). Expression of each GPC was confirmed using 1 µg/mL of an anti-GPC3 mAb (clone 1019718), 1 µg/mL of an anti-GPC2 mAb (clone CT3), 1 µg/mL of an anti-GPC3 mAb (clone SP86), 1 µg/mL of an anti-GPC4 mAb (clone A21050B), 1 µg/mL of an anti-GPC5 mAb (clone 297716), and 1 µg/mL of an anti-PA16 mAb, NZ-1. The cells were then incubated with Alexa Fluor 488-conjugated anti-mouse IgG, anti-rabbit IgG (for SP86), or antirat IgG (for NZ-1). Fluorescence data were collected using the SA3800 Cell Analyzer and analyzed using FlowJo. Experiments were conducted at least twice. Representative histograms are shown.
Determination of KD values of G3Mab-25 by flow cytometry
The binding affinity of G3Mab-25 was determined using a flow cytometry-based approach. The KD values of G3Mab-25 for CHO/GPC3, HepG2, HuH-7, and JHH-5 were 1.8 × 10−8 M, 7.3 × 10−9 M, 3.9 × 10−9 M, and 1.4 × 10−9 M, respectively (Fig. 5). The KD values of SP86 for CHO/GPC3 could not be determined because the sigmoid curve did not reach a plateau (Supplementary Fig. S1). These results indicated that G3Mab-25 has a high affinity to GPC3-positive cells.
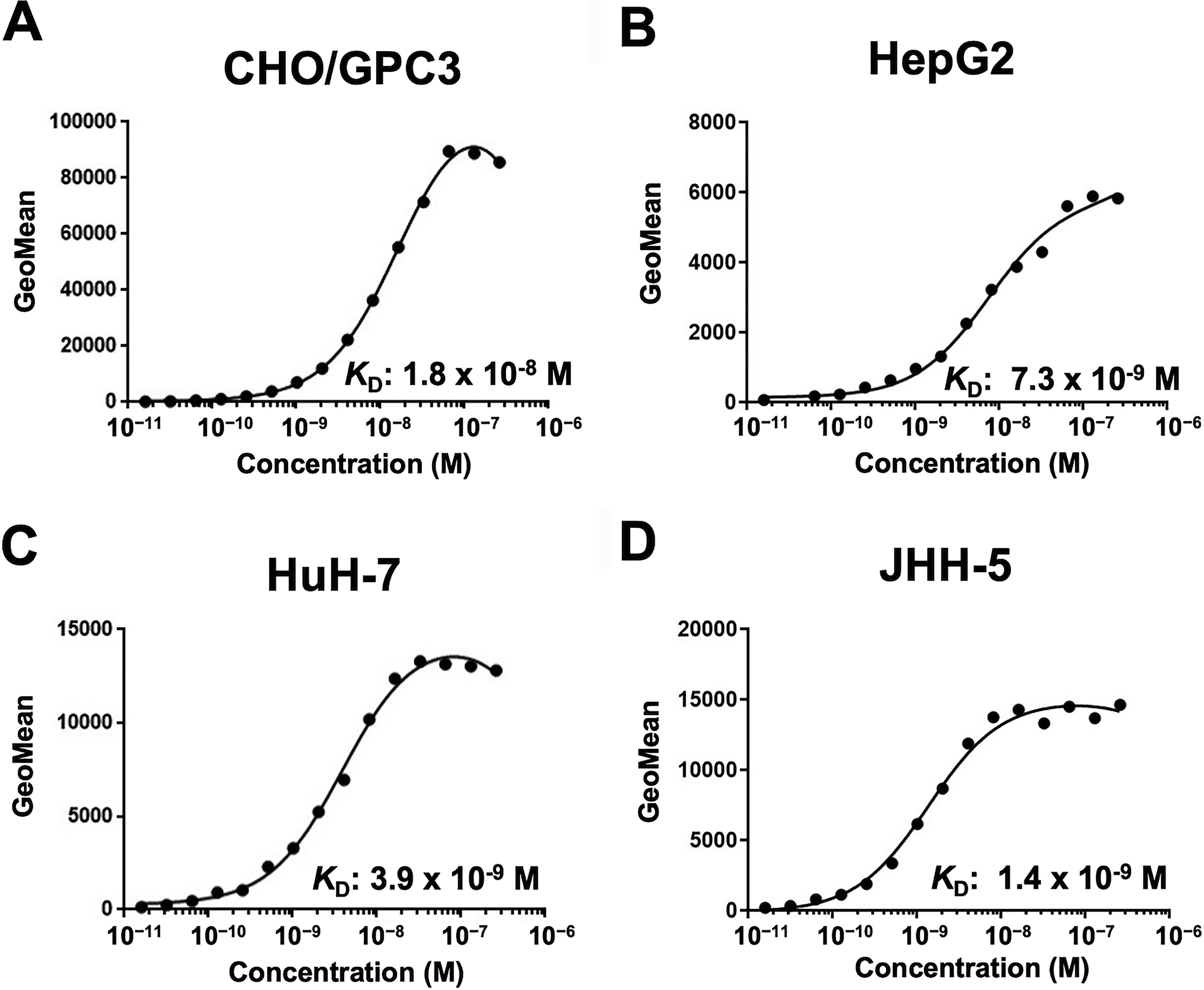
Measurement of the binding affinity of G3Mab-25 using flow cytometry. CHO/GPC3
Western blotting using anti-GPC3 mAbs
We next performed western blotting using G3Mab-25 and SP86. Whole-cell lysates of CHO-K1, CHO/GPC3, HepG2, HuH-7, and JHH-5 were used. G3Mab-25 detected 70-kDa and 40-kDa bands in CHO/GPC3, HepG2, HuH-7, and JHH-5, but not in CHO-K1 (Fig. 6A). SP86 detected 70-kDa and 17-kDa bands in CHO/GPC3 in CHO/GPC3 (Fig. 6B). Although SP86 detected 70-kDa band in HepG2, it was not detected in HuH-7 and JHH-5 (Fig. 6B). Furthermore, the 17-kDa band was weak in HepG2 and was not detected in HuH-7 and JHH-5 (Fig. 6B). IDH1 detected by RcMab-1 was used as an internal control (Fig. 6C). Figure 6D represents the structure of GPC3. These results indicated that G3Mab-25 detected the 70-kDa (uncleaved form) and N-terminal fragment (40-kDa) of GPC3. In contrast, SP86 detected the 70-kDa (uncleaved form) and C-terminal fragment (17-kDa) of GPC3.
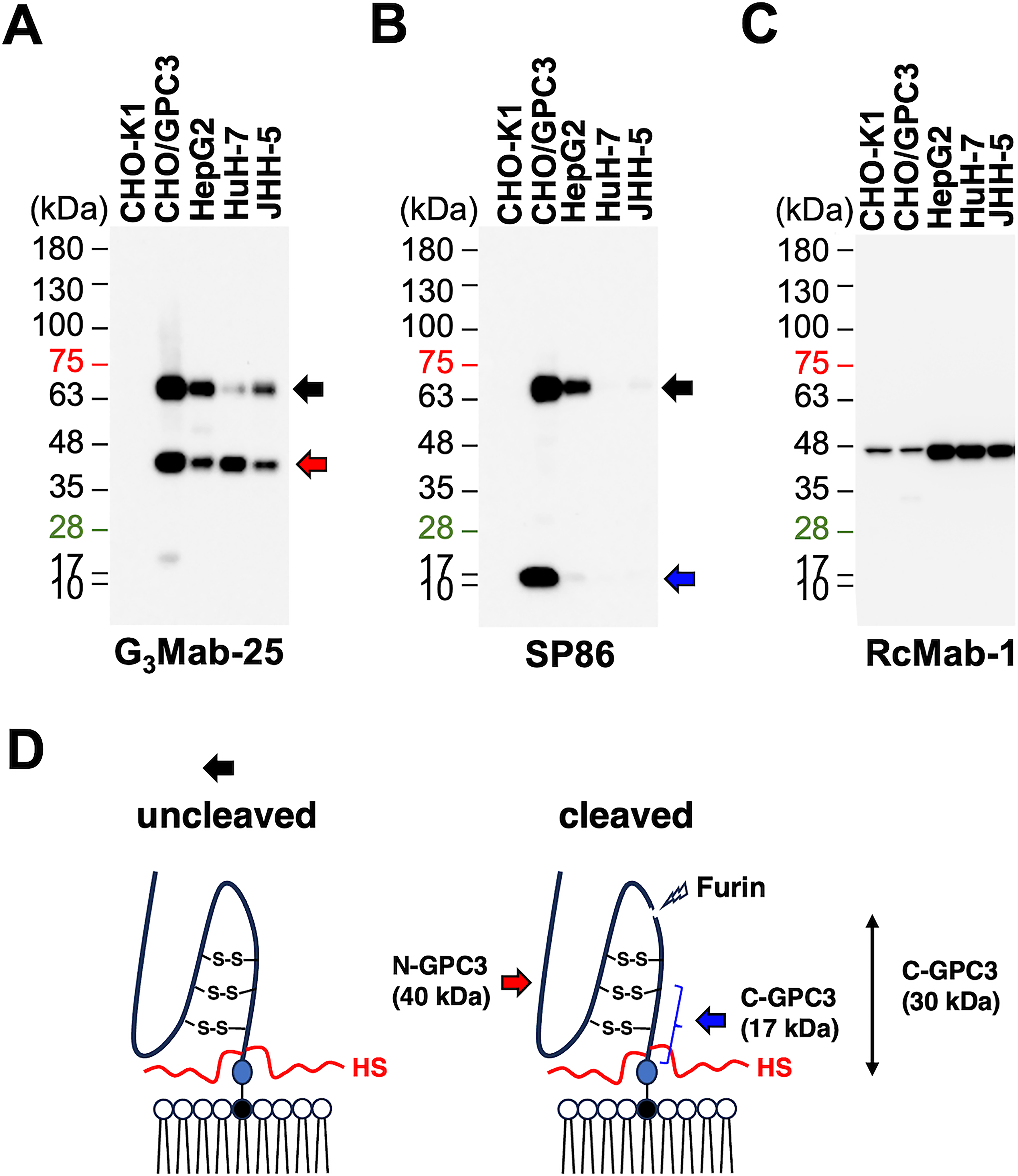
Western blotting using anti-GPC3 mAbs. Cell lysates from CHO-K1, CHO/GPC3, HepG2, HuH-7, and JHH-5 were electrophoresed and transferred to polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of G3Mab-25
IHC using cell blocks and tissue arrays
We next performed IHC in FFPE sections of CHO-K1 and CHO/GPC3 using G3Mab-25 and SP86 with BenchMark ULTRA PLUS, a fully automated slide staining system. Both membranous and cytoplasmic staining were detected in CHO/GPC3 but not in CHO-K1 by G3Mab-25 and SP86 (Fig. 7). These results indicated that G3Mab-25 and SP86 can detect GPC3 in FFPE cell sections.
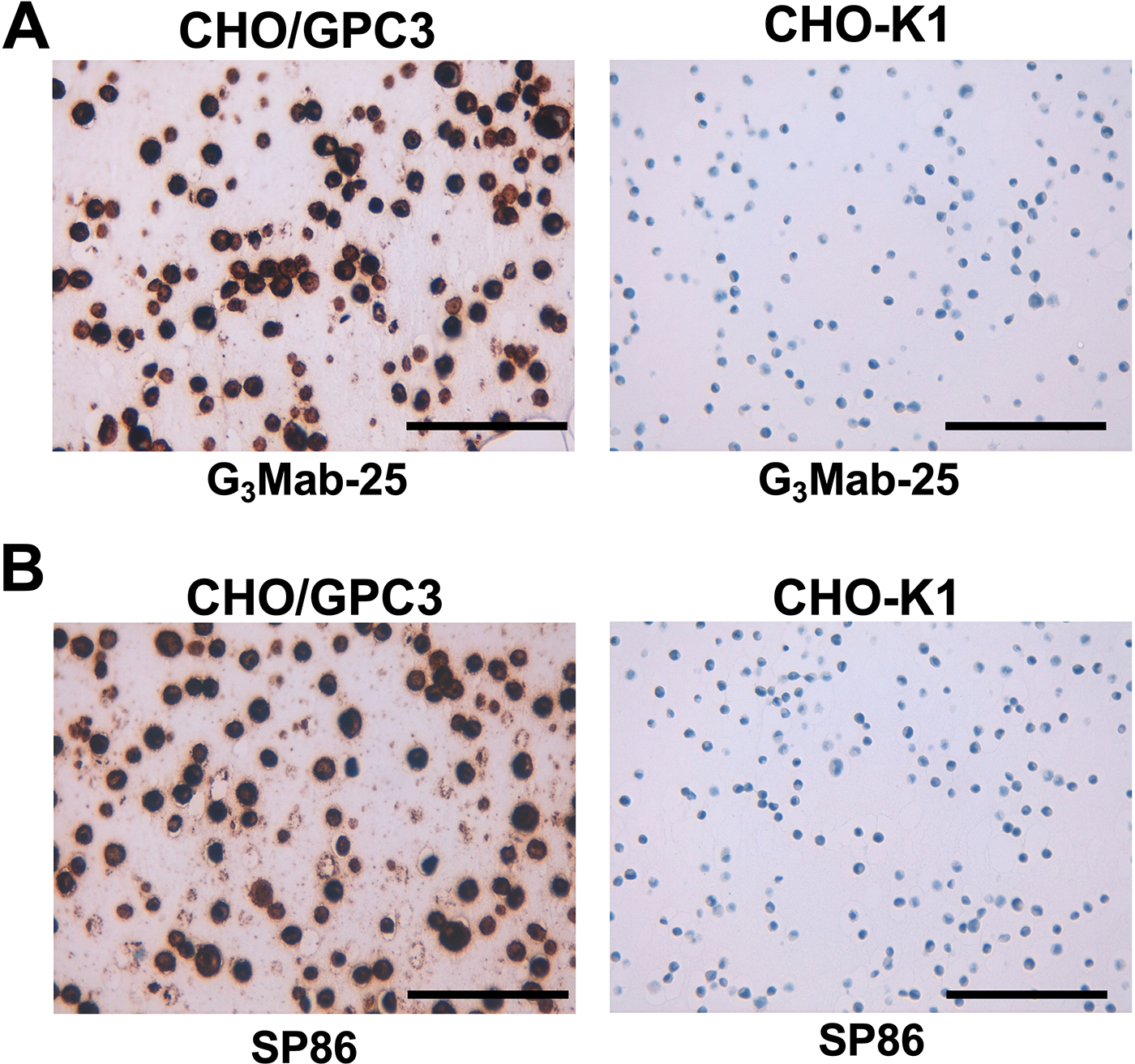
Immunohistochemistry using G3Mab-25 in formalin-fixed paraffin-embedded cell blocks. CHO/GPC3 and CHO-K1 sections were treated with 0.5 μg/mL of G3Mab-25
Since GPC3 is frequently overexpressed in HCC, we next investigated whether G3Mab-25 could detect GPC3 in a tissue array from a solid tumor screening panel. As summarized in Supplementary Figure S2 and Supplementary Table S1, G3Mab-25 showed the cytoplasmic granular staining (score 1) in HCC sections, but not in other tumor sections. We further stained the liver tumor tissue arrays in the same experimental setting. As shown in Figure 8, an intense diffuse cytoplasmic/membranous staining (score 7), cytoplasmic granular/moderate membranous staining (score 5), and weak membranous staining (score 3) were observed. Furthermore, the negative staining (score 0) was also observed in some cases of HCC and normal liver. SP86 showed staining patterns similar to those of G3Mab-25, but staining intensity differed in some cases (Supplementary Fig. S3). Supplementary Table S2 summarizes the results. These results indicated that G3Mab-25 exhibited the diverse staining pattern of GPC3 in HCC tissues.
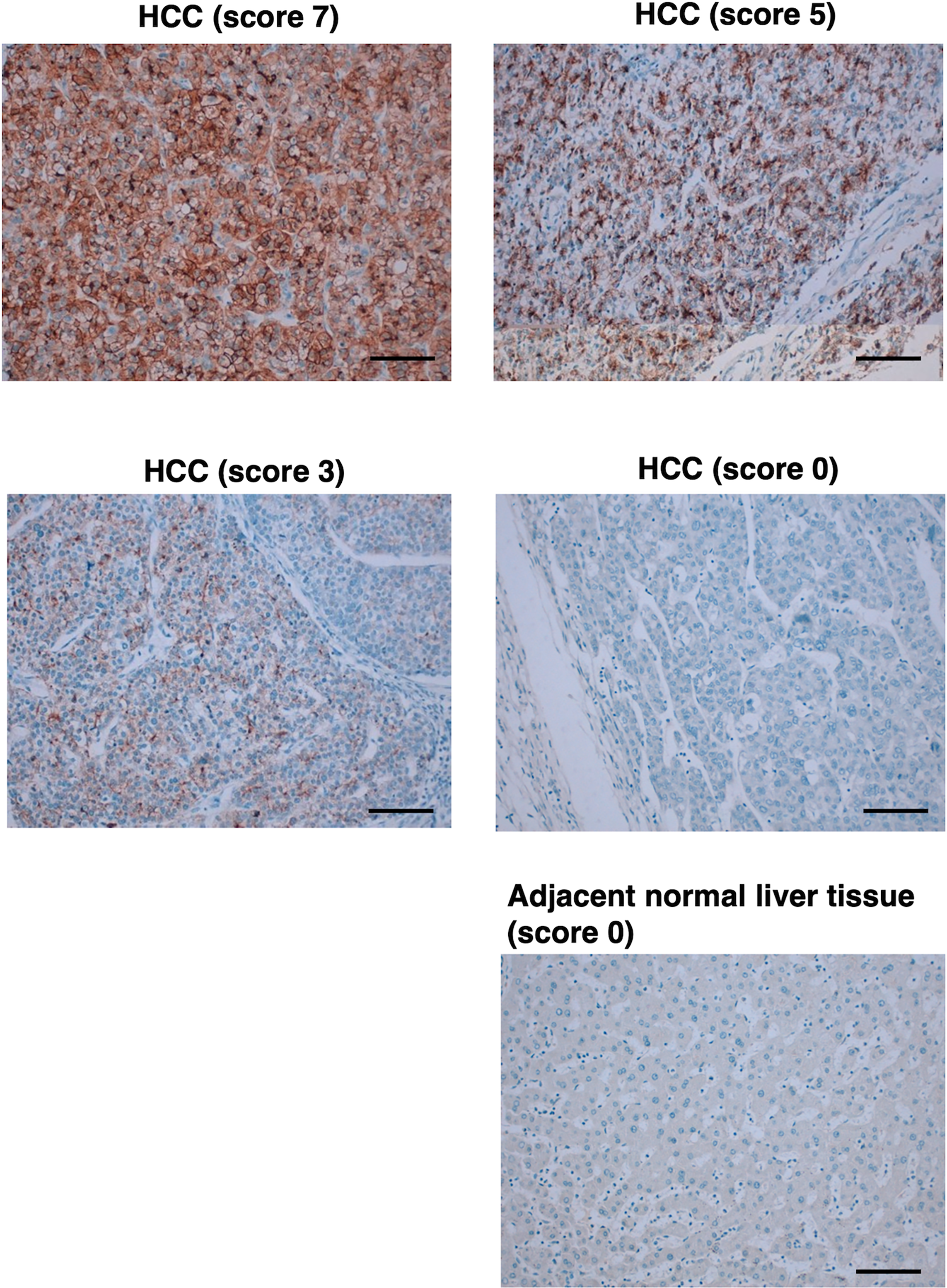
Immunohistochemistry using G3Mab-25 in liver tumor tissue arrays. Liver tumor tissue arrays were treated with 2 μg/mL of G3Mab-25. Staining was performed using the ultraView Universal DAB Detection Kit and BenchMark ULTRA PLUS. Staining intensity was graded as strong membranous (score 7), moderate membranous (score 5), weak membranous (score 3), and no staining (score 0). Scale bar = 100 μm.
Discussion
We have established 88 clones of anti-GPC3 mAbs by the CBIS method and updated the information on our website “Antibody bank (
Wnt signaling is a central regulator of HCC progression. 44 In addition to the canonical receptor Frizzled (FZD), multiple coreceptors, including HS proteoglycans, participate in Wnt signaling activation. GPC3 serves as a Wnt coreceptor promoting HCC cell proliferation through recruitment of Wnt ligands to the C-terminal HS. 45 Furthermore, a structural model of GPC3-Wnt complex revealed that a Wnt-binding groove located in the putative FZD-like cysteine-rich domain at the N-terminal domain of GPC3. 46 The GPC3 groove around residue Phe41 interacts with the middle region of Wnt3a. Mutations within this groove markedly reduced Wnt3a binding, β-catenin activation, and inhibited HCC tumor growth in mice. 46 Significantly, blockade of this domain with a specific antibody against the N-terminal domain of GPC3 suppressed Wnt activation, 46 suggesting a potential therapeutic site on GPC3 for Wnt inhibition and HCC treatment. Since G3Mab-25 recognizes the N-terminal fragment of GPC3 in western blotting (Fig. 6), epitope mapping and the inhibitory effect on Wnt signaling should be investigated in the future.
G3Mab-25 clearly recognizes the endogenous N-terminal 40-kDa fragment of GPC3 (Fig. 6A). In contrast, SP86 recognizes the endogenous C-terminal 17-kDa fragment of GPC3, but its reactivity is lower than that of G3Mab-25 (Fig. 6B). This is probably due to differences in stability between the N-terminal 40-kDa fragment and the C-terminal 17-kDa fragment, as well as differences in binding affinity between G3Mab-25 and SP86 (Fig. 5 and Supplementary Fig. S1). More prolonged exposure of the western blot membrane by SP86 showed the endogenous C-terminal 17-kDa fragment in HCC cell lines (Supplementary Fig. S4A). Additionally, we obtained G3Mab-49 and G3Mab-52, which also recognize the C-terminal 17-kDa fragment (Supplementary Fig. S4B and C). A similar result was reported by another group. 47 After Furin cleavage of GPC3, the C-terminal fragment possesses about 200 amino acids, and the estimated molecular weight is 30-kDa.17,29 Since these mAbs can detect membrane-bound GPC3 in flow cytometry, they probably recognize a juxtamembrane C-terminal region of GPC3 (Fig. 6D). It should be determined whether the C-terminal 30-kDa fragment is further processed to a 17-kDa fragment or not.
IHC is an essential technique for tumor diagnosis. Clinically, BenchMark ULTRA PLUS is used for the diagnosis of HER2-positive tumors, which determines the application of anti-HER2 therapies. 48 For the use of trastuzumab in patients with breast cancer, HER2-overexpression should be determined by intense and complete membranous staining of >10% of cells in IHC 3+. 49 The GPC3 overexpression was evaluated in the different scoring in clinical trials.27,36,50 This study follows a scoring system as recently used in a clinical trial. 36 Therefore, standardization of IHC is essential for patient selection. Since G3Mab-25 works within the standard staining protocol of BenchMark ULTRA PLUS (Figs. 7 and 8). It would contribute to standardizing HCC diagnosis.
An anti-GPC3 mAb clone GC33 (mouse IgG2a, κ) was established by the immunization of bacterially-expressed C-terminal fragment (amino acids 524–563) of GPC3. 23 GC33 exhibited high binding affinity (KD value: 2.4 × 10−10 M in enzyme-linked immunosorbent assay 23 ) ADCC, and antitumor efficacy in a preclinical study. 25 However, the humanized mAb, codrituzumab, failed to improve survival in clinical studies. 29 Codrituzumab may exhibit limited efficacy due to glycosylation-related modifications that affect antibody binding. Glycosylation of antigens, including PD-1 and PD-L1, has been reported to mask binding epitopes and diminish therapeutic activity. 51 Similarly, glycosylation near the C-terminal region or HS modification of GPC3 may limit antibody accessibility. Our anti-GPC3 mAbs established by the CBIS method recognize a variety of epitopes, including conformational epitopes, linear epitopes, and glyco-epitopes, which enable the selection of antitumor efficacy in vivo. We have initiated cDNA cloning from anti-GPC3 mAb clones, including G3Mab-25, and will evaluate ADCC and antitumor efficacy using class-converted (mouse IgG2a or human IgG1) mAbs.
In the development of CAR-T, the property of single-chain variable fragment, including binding affinity and epitope, determines the persistence, tonic signaling, exhaustion, and therapeutic efficacy. 52 Although a variety of CAR against CD19 has been evaluated in the clinic, 53 the variation of anti-GPC3 CAR is still limited. G3Mabs will enable optimization of anti-GPC3 CAR-T cell therapy.
Authors’ Contributions
S.O., H.S., Y.O., and R.I. performed the experiments. M.K.K. and Y.K. designed the experiments. H.S. and Y.K. wrote the article. All authors have read and agreed to the published version of the article.
Footnotes
Author Disclosure Statement
The authors have no conflict of interest.
Funding Information
This research was supported in part by
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.