
Abstract
Objective:
Sonic hedgehog (SHH) signaling is a pivotal regulator of epithelial–mesenchymal interactions and oncogenesis, yet its regulatory dynamics in salivary gland tumors remain poorly defined. This study quantified the mRNA expression of key SHH pathway components—SHH, glioma-associated oncogene (GLI1), suppressor of fused (SUFU), hedgehog-interacting protein (HHIP), and signal transducer and activator of transcription 3 (STAT3)—to differentiate the molecular profiles of benign pleomorphic adenoma (PA) and malignant low-grade mucoepidermoid carcinoma (LG-MEC).
Methods:
In this cross-sectional molecular analysis, quantitative polymerase chain reaction (qPCR) was utilized to analyze 27 tissue samples categorized into: Normal salivary glands (NSG, n = 7), PA (n = 9), and LG-MEC (n = 11). Relative gene expression was calculated using the 2-ΔΔCt (cycle threshold [Ct]) method. Differences between groups were analyzed using the Kruskal–Wallis test with pairwise post-hoc comparisons.
Results:
A significant variation in the expression of SUFU, a key negative regulator of the SHH pathway, was observed across the groups (p = .040). Pairwise analysis revealed a significant downregulation/alteration of SUFU in PA compared to NSG (p = .035), suggesting a loss of pathway inhibition in benign tumorigenesis. Although GLI1 and STAT3 exhibited higher median fold changes in the LG-MEC group, these did not reach statistical significance (p = .151 and p = .158, respectively), likely due to the limited sample size.
Conclusion:
Our findings identify SUFU deregulation as a potential molecular event in the pathogenesis of PA. The lack of significant SHH and STAT3 differentiation between benign and malignant cohorts suggests that canonical pathway activation may be supplemented by complex regulatory feedbacks. These results provide a molecular foundation for larger-scale proteomic validation of the SHH pathway as a diagnostic or therapeutic target in salivary gland oncology.
Abbreviations
SHH–Sonic Hedgehog
GLI–Glioma-associated oncogene
HHIP–Hedgehog-interacting protein
SUFU–Suppressor of fused
STAT–Signal transducer and activator of transcription
PA–Pleomorphic Adenoma
MEC–Mucoepidermoid carcinoma
NSG – Normal salivary glands
PCR–Polymerized chain reaction
PTCH–Patched
Introduction
Salivary gland tumors are a rare and highly diverse group of neoplasms, with an annual incidence of 1.5–4.0 cases per 100,000 people, 1 accounting for less than 0.5% of all tumors and approximately 5% of all head and neck tumors. 2 Pleomorphic adenoma (PA) is the most common benign salivary gland tumor, while adenoid cystic carcinoma and mucoepidermoid carcinoma (MEC) are the most frequent malignancies. 3 The broad histological spectrum of these tumors suggests diverse differentiation pathways, underscoring the need for a thorough understanding of their molecular mechanisms. The sonic hedgehog (SHH) pathway is a potential key player in these mechanisms, given its established role in regulating differentiation and its possible significance in tumorigenesis.
The SHH pathway is a key cell-signaling system that plays a crucial role in processes such as cell patterning, proliferation, fate specification, morphogenesis, and homeostasis. 4 SHH is a ligand-protein within this signaling pathway, which is secreted, and binds to the patched receptor upon secretion, thus negating its inhibitory effect on the smoothened (SMO) transmembrane protein. SMO, in turn, leads to the activation of the glioma-associated oncogene (GLI) family of transcription factors. GLI proteins regulate proliferation, differentiation, and interactions with the extracellular matrix. However, this pathway is controlled by two antagonists: Hedgehog-interacting protein (HHIP) and suppressor of fused (SUFU).4,5 HHIP is a membrane glycoprotein with a high binding affinity to ligands of the Hh pathway. SUFU, on the other hand, binds to GLI proteins, negatively controlling their activity through two mechanisms and sequestering GLI in the cytoplasm or inhibiting GLI transcriptional activity.6–8 Additionally, Signal Transducer and Activator of Transcription 3 (STAT3), a transcription factor, has been shown to have a significant association with the SHH pathway, suggesting its involvement in the regulation and functioning of this critical signaling pathway.9,10
The SHH pathway has been identified in various neoplasms, including hematologic tumors. 11 carcinomas of the breast, 12 stomach, 13 mouth, 14 pancreas, 15 prostate, 16 and thyroid. 17 Therefore, an in-depth understanding of the biology and pathology of the SHH pathway is crucial, and extensive studies have contributed to the development of treatment for these malignancies. Although predominantly dormant in adult tissues, inappropriate reactivation of the SHH pathway has been associated with tumorigenesis, tumor progression, and the maintenance of cancer stem-like cells in various epithelial cancers. 18
The SHH signaling pathway represents a well-conserved developmental pathway that plays a critical role in embryonic development, tissue organization, and organ formation, including salivary gland development. 19 A case involving a cerebellar medulloblastoma with positive SHH markers reveals the intra-axial mature ectopic salivary gland remnants. 20 Experiments on the radiotherapy-induced head and neck cancer survivors indicated that the associated hyposalivation can be restored through SHH pathway activation. 21 These molecular insights suggest the importance of the SHH pathway in salivary gland development. However, existing evidence regarding the roles of the SHH pathway and STAT3 genes in salivary gland tumors is limited, highlighting the need for further research. This preliminary study aims to investigate the expression of key SHH pathway molecules (SHH, GLI1, SUFU, and HHIP) and STAT3 to better understand their functional roles in salivary gland tumors.
Materials and Methods
Samples
The research protocol and ethics were approved by the Medical Ethics Committee, Faculty of Dentistry (FOD), Universiti Malaya (UM) [MEC: OM1715/0035] and the Research Management Center (RMC), MAHSA University, Selangor, Malaysia [RMC/AL12/18]. Formalin-fixed paraffin-embedded (FFPE) tissue sections from 20 salivary gland tumors and seven normal salivary gland (NSG) tissues were retrieved from the Malaysia Oral Cancer Database and Tissue Bank System, which is coordinated by the Oral Cancer Research and Coordinating Center, FOD, UM. These included Group 1: (NSG, n = 7) as healthy reference controls, Group 2: PA (n = 9), and Group 3: Low-grade mucoepidermoid carcinoma (LG-MEC) (n = 11), diagnosed at the Department of Oral and Maxillofacial Clinical Science, FOD, UM. All tissue sections were independently reviewed by two oral and maxillofacial pathologists (AR and DP). Clinical and histopathological data of the selected patients were obtained.
RNA Extraction
Total RNA was isolated from FFPE tissue blocks using the RNeasy FFPE Kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. According to the manufacturer’s protocol, cDNA conversion was performed using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Massachusetts, USA). The quantitation of targeted gene expression assay was performed on Applied Biosystem 7500 Fast Real-Time PCR System (Thermo Fisher Scientific, Massachusetts, USA) as triplicate reactions using specific designed Taqman→ probe assay for SHH (Taqman→ Assay ID: Hs00179843_m1), GLI1 (Taqman→ Assay ID: Hs01110776_g1), SUFU (Tagman→ Assay ID: Hs0096052_m1), HHIP (Tagman→ Assay ID: Hs00368450_m1), and STAT3 (Tagman→ Assay ID: Hs00374280_m1) together with housekeeping gene GAPDH (Tagman→ Assay ID: Hs00266705_m1) as the internal control.
RNA Isolation and Quantification
The extracted RNA samples were qualified and quantified using the NanoDrop 2000 spectrophotometer (Thermo Fisher, USA) and the Bioanalyzer system (Agilent, USA). RNA samples with insufficient RNA yield or RNA integrity number (RIN) of less than two were excluded from the subsequent experiment. Tissue sectioning and RNA extraction were repeated until the desired results were obtained.
Gene Expression Assays Using Quantitative PCR
The comparative quantification of gene expression was analyzed using ΔΔCt, which compares the cycle threshold (Ct) of genes of interest between cases and healthy reference controls, and the threshold Ct of GAPDH. With this method, Ct values for the SHH pathway genes in both cases and the reference control were adjusted using internal control gene Ct values from the same samples. The resulting ΔΔCt value was used to determine the fold change in gene expression. The quantitative polymerase chain reaction (qPCR) protocol was set to a maximum of 40 cycles. Samples that failed to reach the threshold within these 40 cycles were considered undetected and excluded from further fold change calculations.
Statistical Analysis
The Kruskal–Wallis nonparametric test was used to compare median fold changes in gene expression across the three groups and to determine significance levels. The significance level for this analysis was set at p < .05, indicating that any observed differences with a p value below this threshold would be considered statistically significant. Post hoc pairwise comparisons were performed using Dunn’s test to identify specific group differences.
Results
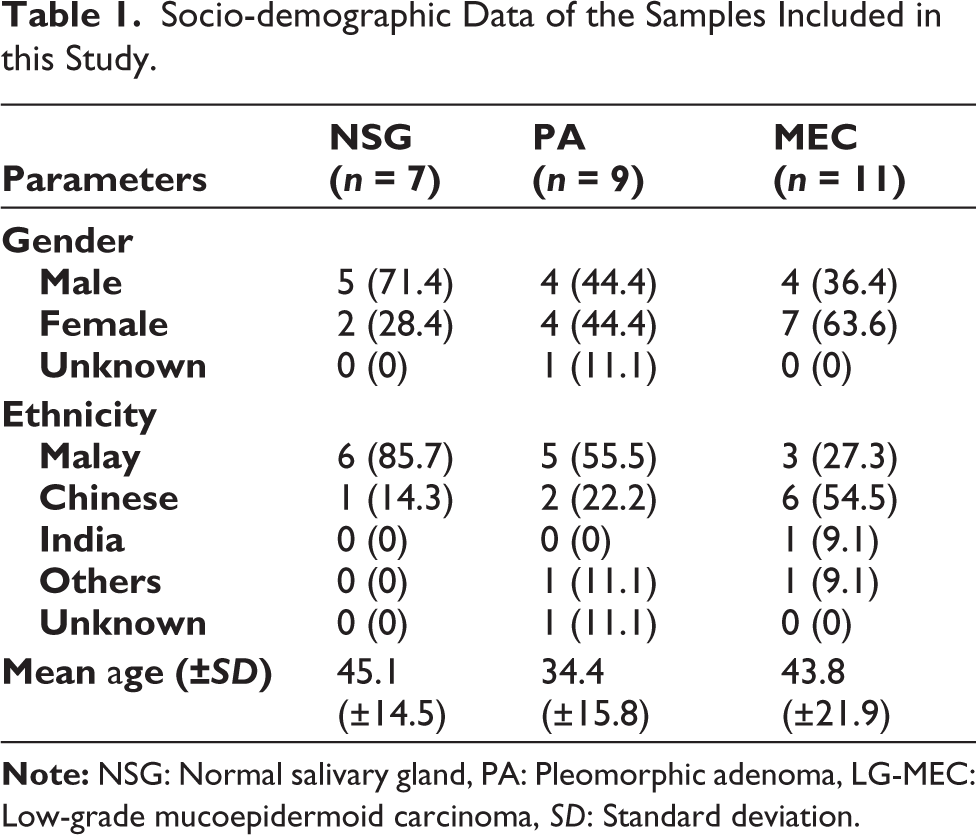
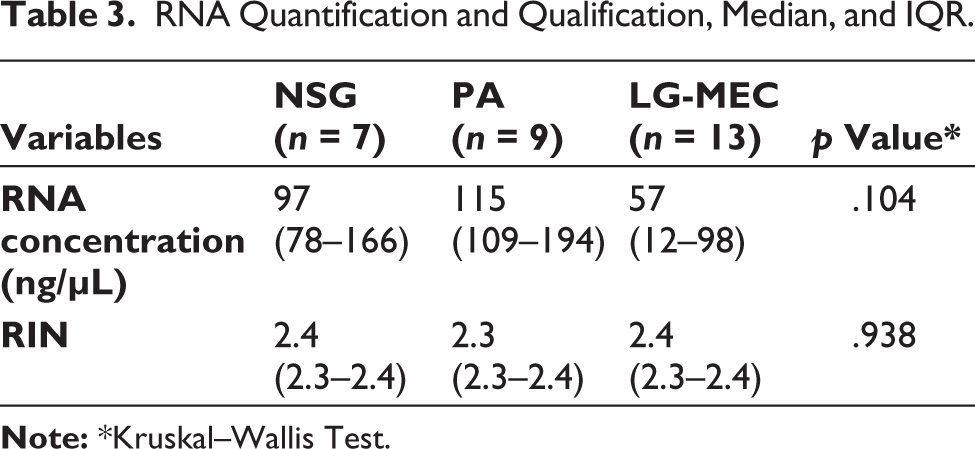
Tables 1 and 2 summarize the socio-demographic and clinicopathological data for the PA and LG-MEC samples, excluding NSG. Notably, most cases in the NSG and PA tissues were among Malay individuals and males. In contrast, the LG-MEC group represented more Chinese and female individuals. The RNA quantification and qualification were summarized in Table 3; there were no significant differences in RNA yield and RIN across tissues. The RNA concentration and RIN for each sample analyzed in this study are detailed in Supplementary Table S1.
Socio-demographic Data of the Samples Included in this Study.
Clinico-pathological Features of the Tumor Samples Included in this Study.

RNA Quantification and Qualification, Median, and IQR.
Comparative Gene Expression
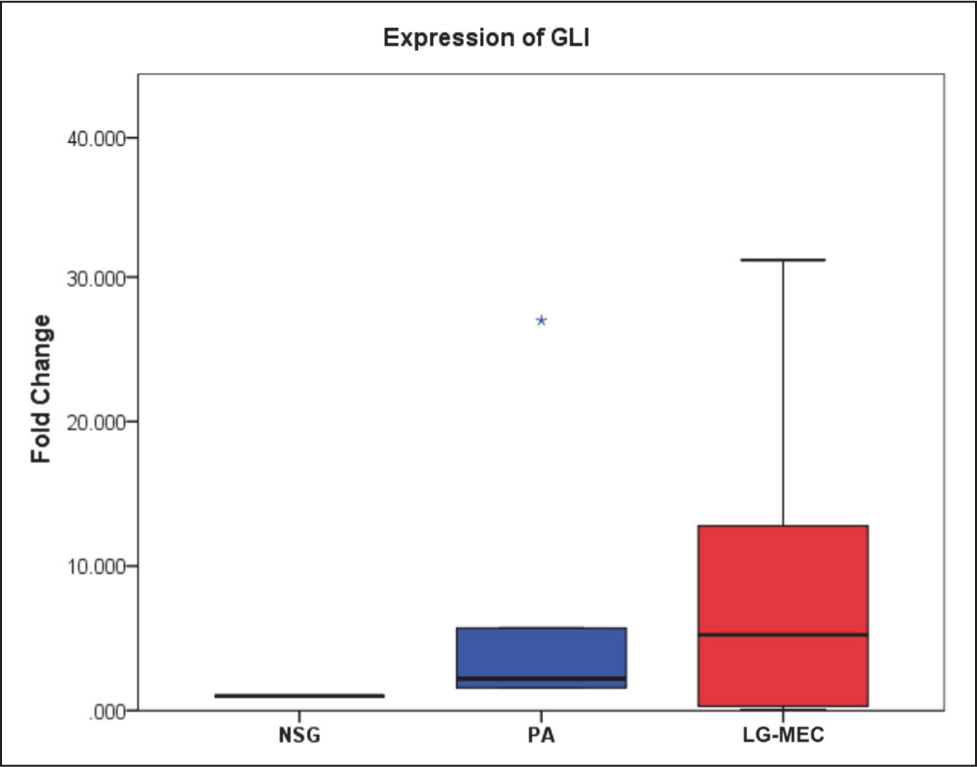
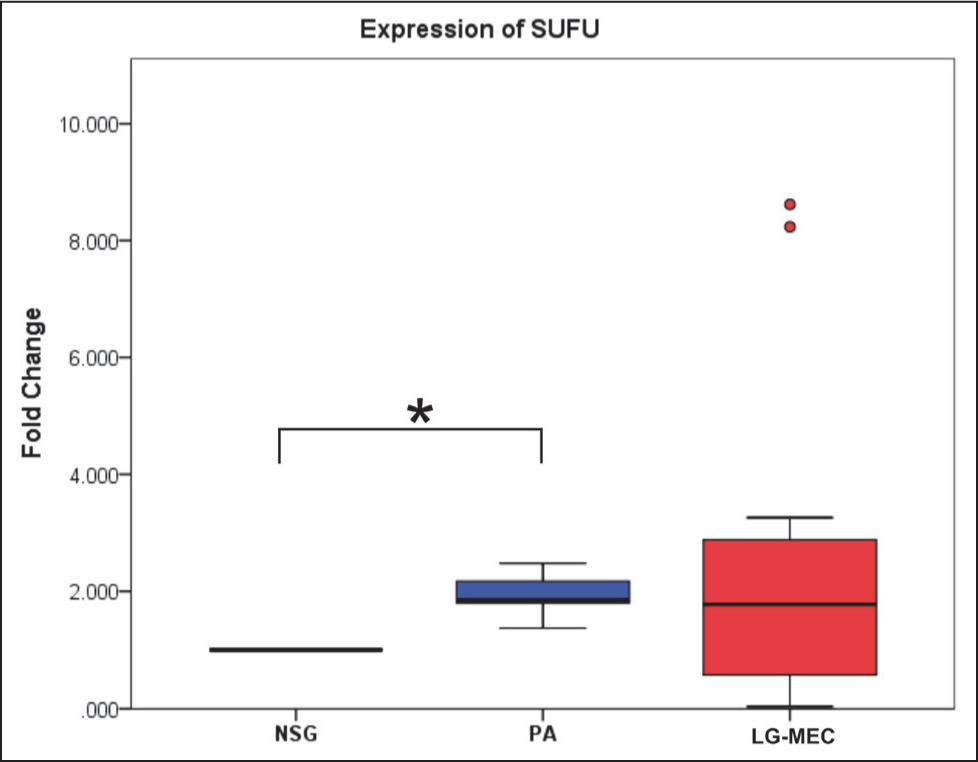
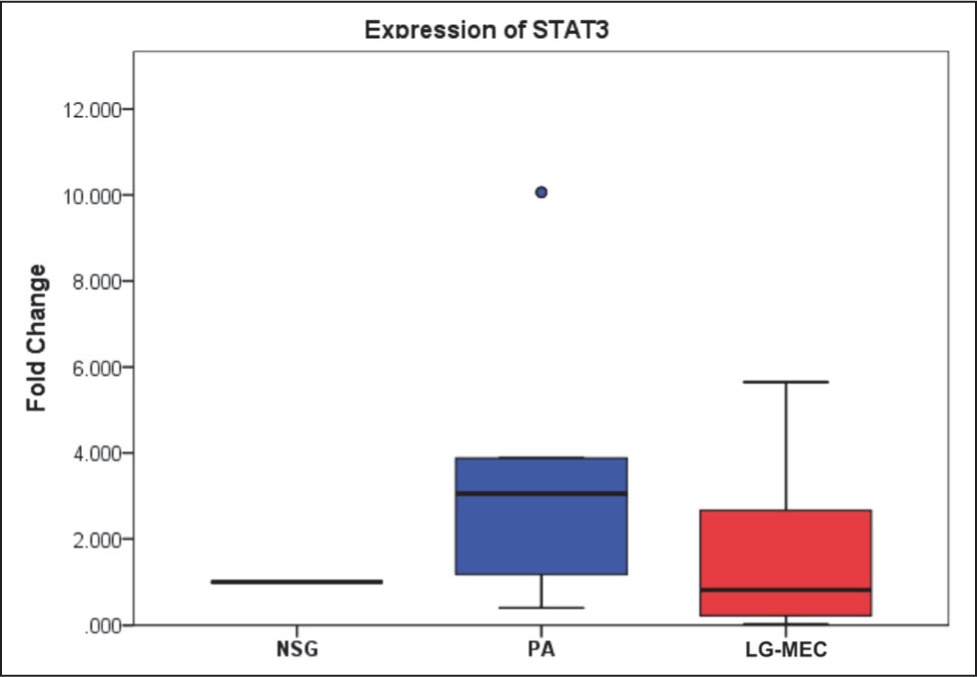
This study analyzed five genes involved in the SHH pathway using qPCR. Among these, Ct values for SHH and HHIP were undetermined after 40 cycles of amplification in all samples; the expression of these genes was below the detection limit of the qPCR assay in the present study. The relative expression levels of GLI1, SUFU, and STAT3 were then compared across different tissue types. The median expression levels of GLI1 showed no statistically significant differences across the NSG, PA, and LG-MEC groups (p = .151) (Table 4, Figure 1). PA and LG-MEC, compared to NSG, showed elevated median fold change of 2.21 (IQR = 9.45) and 5.23 (IQR = 2.505), respectively. In the case of STAT3, the median fold change varied numerically across the groups but did not reach statistical significance (p = .158). Specifically, while PA showed an elevated median fold change of 3.05 (IQR = 2.82) relative to NSG tissue (1.0), LG-MEC tissues exhibited a reduced median fold change of 0.61 (IQR = 2.42). Notably, only SUFU showed a statistically significant difference in expression between the groups (p value = .040). Further pairwise analysis revealed that SUFU expression was significantly elevated in PA tissues compared to NSG (p value = .035) (Table 3, Figure 2), and the expression of GLI1 and STAT3 showed that the pairwise median fold change observed in PA and LG-MEC relative to NSG was not statistically significant (Table 4, Figures 1 and 3).
Expression of GLI1 in Pleomorphic Adenoma (PA) and Low-grade Mucoepidermoid Carcinoma (LG-MEC) After Normalization With Normal Salivary Gland (NSG) Tissue. No Significant Difference in Median Fold Change Was Observed Among the Groups (p = .151).
Pairwise Comparison of the Median of Fold Change Associated with GLI1, SUFU, and STAT3 Genes.

Expression of SUFU in Pleomorphic Adenoma (PA) and Low-Grade Mucoepidermoid Carcinoma (LG-MEC) After Normalization with Normal Salivary Gland (NSG) Tissue. The Kruskal–Wallis Test Showed a Significant Difference in Median Fold Change Among the Groups (p = .040). Pairwise Analysis Revealed a Significant Difference Between NSG and PA (*p = 0.035), but Not Between the Other Group Comparisons.
Expression of STAT3 in Pleomorphic Adenoma (PA) and Mucoepidermoid Carcinoma (MEC) After Normalization With Normal Salivary Gland (NSG) Tissue. No Significant Difference in Median Fold Change Was Observed Among the Groups (p = .158).
Discussion
The salivary gland tumors are believed to comprise a morphologically diverse group. Variations in clinical features and unpredictable prognostic indicators are analogous to the cytomorphologic variations observed in salivary gland tumors. 22 The SHH pathway genes are believed to be essential for morphological development and tissue regeneration, and any aberrations in this pathway have been linked to tumourigenesis.23,24 Understanding the intricacies of this pathway is vital for elucidating the molecular mechanisms underlying morphologically diverse salivary gland tumors.
The prevalence of PA was higher among the Malay population, whereas LG-MEC was more commonly associated with the Chinese population. The clinical manifestations observed in the subjects of the current investigation align with the demographic features of PA and MEC documented within the Malaysian population.25,26
This study has demonstrated a statistically significant upregulation of the SUFU gene within the SHH signaling pathway in PA compared to NSG tissue. SUFU is postulated to serve an adverse regulatory effect on the SHH signaling pathway by modulating the transcriptional activity of GLI, 27 thereby acting as a tumor suppressor gene. The present study reveals that SUFU gene expression is significantly upregulated in PA. Several plausible explanations may account for this apparent upregulation. This may reflect a feedback response or a compensatory cellular response to counterbalance the proliferative signals driven by GLI-mediated transcription in the SHH pathway.28,29 Furthermore, this may suggest that SUFU functions as a barrier to the oncogene-induced senescence mechanism in benign neoplasms and to malignant transformation. 30 It may also indicate maintenance of epigenetic integrity, with SUFU remaining hypomethylated and functionally active in benign tumors. 31 This protective response helps restrain progression and explains why PAs, despite proliferative potential, usually remain benign and non-invasive.
The pairwise analysis of median fold changes revealed that SUFU showed significant differences between NSG and PA, but not between LG-MEC and NSG. This differential expression provides a basis for distinguishing between benign and malignant tumors, thereby assisting in diagnosis and subsequent treatment planning. 32 Notably, SUFU gene expression in LG-MEC displayed a broad range of fold change from 0.036 to 8.617, suggesting considerable variability in expression levels within the LG-MEC group. This wide range of expression suggests heterogeneity in LG-MEC and may indicate underlying biological diversity within this group. Therefore, further investigation is required to determine whether the observed low SUFU levels represent a genuine biological phenomenon or a technical artifact arising from long-term storage. Studies have reported that malignant tumors often harbor genetic or epigenetic alterations that turn off these feedback loops, which could also explain the study’s results. 33
Three LG-MEC samples in the present study revealed total downregulation of GLI1, SUFU, and STAT3 markers. However, it is critical to note that these specific samples were archival tissues stored for more than 10 years, suggesting that age-related RNA degradation may have compromised nucleic acid integrity. This implies the importance of accounting for tissue age and preservation status when selecting samples for gene expression studies.
The samples that remained undetected after 40 cycles of amplification for SHH and HHIP require careful interpretation, as the results likely reflect a combination of low biological expression and the technical limitations inherent to nucleic acid extraction from FFPE material. As indicated in Table 3, the RNA RIN values were consistently low across all tissues. Such reduced RIN values are expected in long-term-preserved FFPE specimens due to protein-DNA/RNA cross-linking, the specific fixative used, and nucleic acid degradation during storage. 34 If the endogenous SHH or HHIP mRNA fragments were shorter than the designed amplicon length, the primers and probes would fail to bind to and amplify the target sequence, potentially leading to false-negative results. 35
The present study did not find any statistically significant difference in GLI1 and STAT3 expression between PA, LG-MEC, and NSG. The correlation between primary cilia and GL1 expression in these tumors, and the association of the aggressive nature of the LG-MEC with GL1, could explain the variations of GL1 expression observed in the PA and LG-MEC.32,36,37 Studies have shown that STAT3 expression in LG-MEC is generally lower than in PA, which is attributed to differences in tumor biological behavior. 32
The present study is among the pioneering studies to provide valuable insights into the role of SHH pathway genes in salivary gland tumors by demonstrating RNA extraction from formalin-fixed tissue sections; however, it has several limitations. The limited sample size in the present study constitutes a significant constraint, as generalizing the findings becomes difficult. A larger sample size, including subtypes and different salivary gland tumors, will be needed to provide a clear understanding of tumourigenesis associated with SHH pathway genes. The mRNA extraction, along with the corresponding protein expression data, would have strengthened the findings of the present study.
Conclusion
The SUFU gene in the SHH pathway showed elevated expression in PA, suggesting a potential role in its pathogenesis. The preliminary study findings indicate that the SUFU gene may play a role in tumor maintenance in PA. However, the specific roles of these genes in LG-MEC remain unclear, highlighting the need for further investigations.
Footnotes
Acknowledgements
The authors would like to thank all the laboratory staff who helped with the laboratory work.
Authors’ Contribution
D.P. contributed to the conception, design, lab analysis, and interpretation, as well as drafted and critically revised the manuscript. W.G.R. contributed to the laboratory analysis, methodology development, and statistical analysis. L.F.F. contributed to the laboratory analysis and development of methodology, A.R. contributed to design, technical and material support, drafted and critically revised the manuscript, and R.B.Z. contributed to the design and guided the research project. All authors read and approved the final article.
Consent for Publication
Not applicable.
Data Availability Statement
The data analyzed during this study are available from the corresponding authors upon request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
All methods were carried out in accordance with the Declaration of Helsinki (2013). We also confirm that we complied with the Personal Data Protection Act 2010 to ensure the security of the participants’ personal data. All experimental protocols were approved by the Ethic Committees of the RMC, MAHSA University (Ethical approval no. RMC/AL12/18) and Medical Ethics Committee, FOD, UM [MEC: OM1715/0035].
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: RMC, MAHSA University, funded the study [RP150-07/18].
Informed Consent
Written informed consent was obtained from all study participants for the collection, long-term storage at the Oral Cancer Research & Coordinating Centre (OCRCC) biobank, and use of their tissue specimens for future research.