
Abstract
Osteoporotic and age-related fractures are a significant public health problem. The current standard of osteoporosis assessment via bone mineral density has been shown to be inadequate for fracture risk predictions highlighting the importance of material composition and structural design of bone in determining skeletal fragility. Bone is a hierarchical material that derives its fracture resistance from various mechanisms that act at length scales ranging from nano- to macroscale. Recent research efforts have focussed on the understanding of bone fracture based on this hierarchical structure to provide a more reliable assessment of fracture risk. Understanding the function, contribution and interaction of each length scale to bone toughness is a crucial step to develop new strategies for fracture risk assessment, fracture prevention, and development of therapeutic interventions for disease and age-related changes in bone. This review presents a hierarchical perspective of bone toughness ranging from nano- to macroscale and reports on the current state of knowledge in the areas of experimental and computational approaches to bone fracture.
Introduction
Osteoporotic and age-related fractures are a significant public health problem affecting the quality of life, morbidity and mortality, and health care costs. 1,2 The current standard of osteoporosis assessment via bone mineral density, measured using dual energy X-ray absorptiometry (DXA), has been shown to be inadequate for fracture risk predictions. 3–5 Material composition and structural design of bone, defined as bone quality, have a significant role in determining the propensity of bone to fracture and have the potential to improve fracture risk assessment. 6–8 Recently, there has been an increased emphasis on bone’s hierarchical structure and its influence on bone quality. One of the important measures of bone quality is bone toughness. At each length scale, different mechanisms contribute to bone toughness. Understanding the function, contribution and interaction of each length scale in determining bone toughness is a crucial step to develop new strategies for fracture risk assessment, fracture prevention, and development of therapeutic interventions for disease and age-related changes in bone.
With the goal of providing an in depth understanding of bone fracture and toughness, this review presents a hierarchical perspective of bone toughness ranging from nano- to macroscale and reports on the current state of knowledge in the areas of experimental and computational approaches to bone toughness evaluation. It presents a synthesis of various fracture mechanisms present at each length scale and summarises their influence on overall fracture toughness of bone.
Hierarchical structure of bone
In order to understand the influence of bone quality on fracture toughness of bone a thorough understanding of the hierarchical structure of bone is necessary. Complete descriptions of the hierarchical structure of bone have been previously reported. 9–12 In this section, the main characteristics of each hierarchical level are summarised. The hierarchical structure of bone can be divided into three broad levels (Fig. 1).
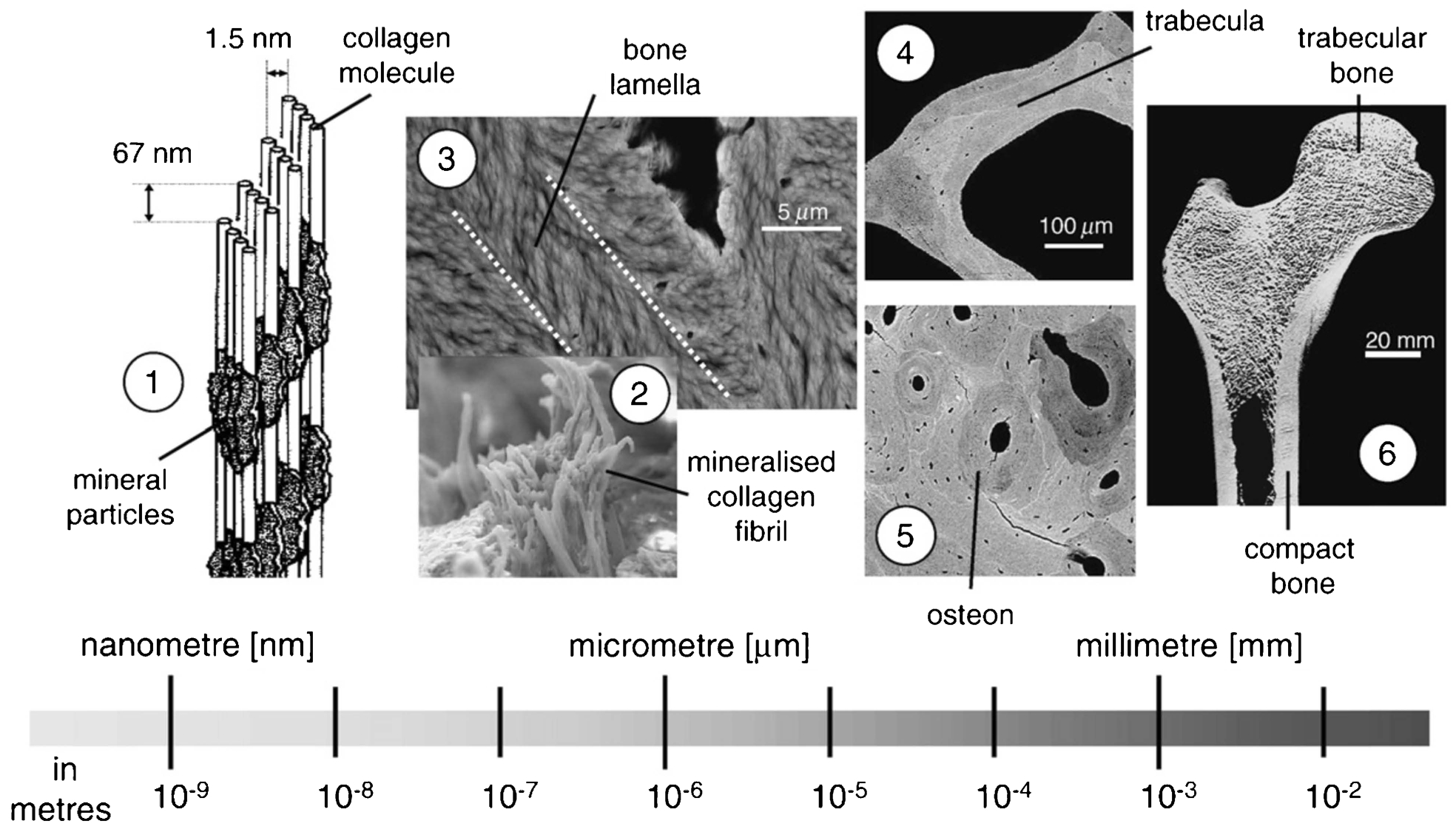
(1) collagen molecules and mineral particles, (2) mineralised collagen fibrils, (3) bone lamella, (4) trabeculae in cancellous bone, (5) osteons in cortical bone, (6) cancellous (trabecular) bone and cortical (compact) bone forming the whole bone (Reprinted from Weinkamer and Fratzl 195 with permission from Elsevier)
Macroscale
The largest scale associated with the bone is the whole bone level that defines the size and shape of the bone. At this level, two distinct bone types can be identified, which are cortical (compact) and cancellous (trabecular) bone (Fig. 1). Cortical bone is the dense bone tissue with low porosity (5–10%) that forms the shell surrounding the bones. Its thickness varies between several centimetres in the midshaft of long bones to several tens of a millimetre in vertebrae. 11 The interior part of the bone, which is enclosed by the cortical shell is filled with cancellous bone composed of trabecular struts that form a porous structure (75–95%) filled with marrow. The most important macroscale properties that influence bone quality are the geometric properties of cortical bone, which represent the size and distribution of bone tissue, trabecular architecture that measures the integrity of cancellous bone, and tissue-level properties of both cortical and cancellous bone.
Microscale
One level down is the cortical bone microstructure and individual trabecula that form the cancellous bone (Fig. 1). The microstructure of cortical bone is composed of osteons with an average size of 200–250 μm 9 that run approximately parallel to the axial direction of long bones. Osteons are formed by concentric layers of lamellae with a typical thickness of 3–7 μm. 9 Lamellae surround a central canal called Haversian canal that includes blood vessels with an average size of 50 μm. 13 Osteons are surrounded by a thin layer called the cement line, which is 1–5 μm thick. Cement lines are formed during the reversal from bone resorption to bone formation. 14 At the microscale, cancellous bone is composed of individual trabeculae of about 200 μm in size. 13 Trabeculae have a lamellar structure that usually does not contain osteons. 12
Nanoscale
At the nanoscale, bone is composed of organic matrix and minerals (Fig. 1). The organic matrix is primarily made up of type I collagen (∼90%) with smaller amounts of other types of collagen and non-collagenous proteins (∼10%) such as osteopontin (OPN) and osteocalcin (OC). The mineral phase of bone is predominantly composed of hydroxyapatite. Type I collagen is the most abundant organic component of bone, which is made up of tropocollagen protein molecules. Tropocollagen molecules form a staggered arrangement with 67 nm periodicity and 40 nm gaps between molecules held together by intermolecular cross-links, which make up collagen fibrils. Mineralisation of the collagen fibrils occurs through the deposition of hydroxyapatite crystals in the gaps between the tropocollagen molecules. 15 The mineralised collagen fibrils further form fibril arrays that are connected by a protein phase. These fibril arrays bundle together in different orientations to form a lamella, which is the building block of osteons at the microscale.
The hierarchical structure of bone outlined above results in multiscale fracture mechanisms that are directly influenced by the changes in bone quality. In the following sections, the influence of each material scale on fracture toughness of bone and their relationship with bone quality measures will be reviewed with a particular emphasis on cortical bone.
Nanoscale bone toughness
Contribution of nanoscale components to fracture behaviour of bone
As highlighted above, bone is composed of organic matrix and minerals at the nanoscale. The organic matrix and mineral amount, distribution and characteristics change with age and disease and may lead to changes in the mechanical properties of bone including its fracture resistance. 16
The organic matrix of bone undergoes age-related changes that reduce the collagen amount and result in post-translational changes that affect the collagen cross-links. 17,18 One of the biochemical alterations that occur in bone with age is non-enzymatic glycation (NEG). Non-enzymatic glycation leads to the formation of advanced glycation end products (AGEs) and modifies the cross-linking profile of collagen fibrils. 18–21 Accumulation of AGEs with age or disease negatively affects the mechanical properties of bone. Negative correlations have been found between AGEs and post-yield energy dissipation in both cancellous and cortical bone. 22–24 Reduction in the fracture toughness of bone has been correlated with increasing AGEs. 25,26 In addition, the effectiveness of the load transfer from tissue-level to mineralised collagen fibrils through fibrillar sliding has been shown to decrease with higher AGE content. 27 These observations highlight the possible adverse effects of increased non-enzymatic cross-links on the energy dissipation mechanisms at the nanoscale. A further evidence of these adverse effects has been shown in a computational study that demonstrated a reduction in dissipated energy with increasing number of non-enzymatic cross-links. 28 Contrary to the increase in NEG, enzymatic cross-linking has been shown to decrease with age. 20 A recent study has demonstrated that NEG led to the deterioration of mechanical properties in trabecular bone whereas enzymatic cross-links were beneficial. 29 These observations are further supported by a reduction in enzymatic cross-links and increase in NEG in femoral neck fracture cases compared to controls. 30
Although, there is a large body of evidence that underlines the adverse effects of non-enzymatic cross-links on the fracture behaviour of bone, other accompanying changes in the collagen such as collagen connectivity may also contribute to the mechanical response of bone. Willett et al. 31 have shown that the ultimate strain and toughness of bone increased with increasing pentosidine and total cross-linking in ribosylated bone. In addition, a positive correlation between collagen connectivity and bone toughness has been found. 32 The results indicate the probable role that the collagen integrity may play in determining the mechanical properties of bone. These findings raise the question about the compensating effect of non-enzymatic cross-links in retaining the collagen connectivity when enzymatic cross-links are reduced due to age or disease. Although this is a plausible mechanism, the impairment of energy dissipation mechanisms such as fibrillar sliding due to AGEs may counteract the mechanical benefits gained from collagen connectivity due to non-enzymatic cross-links.
Other modifications in collagen that may influence the mechanical properties of bone are the collagen fibril structure and orientation. In osteoporotic bone, collagen fibril size is reduced and collagen fibrils form an irregular arrangement compared to a regular structure in healthy bone. 33 Changes in the collagen fibril arrangement may have implications for the fracture behaviour of bone as experimental observations show that the fracture toughness of bone is influenced by the angle between the crack path and the mean collagen orientation. 34
There is also increasing evidence that non-collagenous proteins including OPN and OC significantly influence the fracture behaviour of bone. 16 Osteopontin and osteocalcin both regulate bone formation and resorption, provide a scaffold for mineral deposition, and control hydroxyapatite crystallisation. 16 These proteins were shown to have a significant contribution to the nanoscale energy dissipation mechanisms in bone including dilatational band formation, and stretching and rupture of sacrificial bonds described below. 35,36 Studies showed that the fracture toughness and work to fracture decrease in OPN deficient mice independent of other factors, 37,38 which shows the important contribution of OPN to fracture toughness of bone. Evidence of increased mineral acquisition and impaired microdamage formation with the lack of OC in mouse bone has been shown in a previous study. 39 In addition, propagation fracture toughness was shown to decrease in OC, OPN, and both OC and OPN deficient mice bone accompanied by lack of formation of diffuse damage and dilatational bands. 36 Furthermore, the reduction of OPN in younger osteonal bone compared to older interstitial bone 40 may explain the age-related reduction in fracture toughness of bone based on protein deficiency.
Similar to the organic content of bone, the amount and composition of mineral phase also undergoes changes with age and disease and influence the mechanical response of bone. Previous studies have shown that there is an inverse highly non-linear relationship between bone toughness and its mineral content. 41–43 The increase in mineral content with age 43 results in more brittle bone, which is expected to enhance damage accumulation and reduce the resistance of bone to fracture. In addition to the amount of mineralisation, its distribution may also significantly change with age or disease. Heterogeneity in mineral-to-matrix ratio and carbonate-to-phosphate ratio was lower whereas heterogeneity of crystallinity was higher in the femoral neck fracture cases compared to the controls. 44 In addition, mineral-to-matrix ratio was lower in fracture cases. 44 These studies highlight that the changes in the bone mineral content, crystal size, and structure may also adversely affect the energy dissipation and fracture resistance of bone.
Nanoscale fracture mechanisms
At the nanoscale, several energy dissipation and fracture toughening mechanisms have been identified including molecular uncoiling, dilatational band formation, and fibrillar sliding of mineralised collagen fibrils. 36,45
Molecular uncoiling involves energy dissipation due to stretching and unwinding of individual collagen molecules involving hydrogen-bond breakage. 46 The energy dissipation based on this mechanism is dependent on the intermolecular cross-links. Molecular analysis performed on collagen fibrils demonstrated that the energy dissipating deformation mechanism via molecular uncoiling was adversely affected by an increase in the number of intermolecular cross-links within the fibril leading to brittle fracture behaviour. 46 These results provide evidence of one of the possible reasons for the change in the fracture properties of bone that occurs as a result of accumulation of AGEs in bone. 24,25
Dilatational bands (Fig. 2a ) are associated with the formation of ellipsoidal voids between fused mineral aggregates that occur as a result of the unfolding of the non-collagenous OC–OPN protein complex embedded in the mineralised bone matrix. 36 The energy dissipation due to extension and separation of the OPN–OC complex is followed by the deformation and rupture of collagen fibrils. The growth of these bands in size and number leads to the formation of diffuse damage at the next hierarchical level. 36

Molecular slip between tropocollagen molecules and slip along hydroxyapatite and tropocollagen interfaces also provide an energy dissipation mechanism. 47 Due to the adhesion between the mineral platelets and the collagen, mineral carries a part of the tissue strain and transfers it to the mineralised collagen fibril. This increases the strain level that causes intermolecular slip and it initiates higher order energy dissipation in the form of microcracks. 47
In particular, the mineralised collagen fibrils are bonded by a glue-like material composed of a non-fibrillar organic matrix that resists the separation of the fibrils (Fig. 2b ). 35,48 Intermolecular slip and fibrillar sliding of the collagen fibrils provide toughening through the sacrificial bonds in the glue material that dissipate energy as they stretch and break. 35 Although the exact nature of these sacrificial bonds is not known, OPN has been suggested as a candidate protein because it is known to create networks stabilised with sacrificial bonds. 49
The importance of nanoscale mechanisms on the overall mechanical behaviour of bone is highlighted in the findings that demonstrate a tissue strain level which is six times the mineral level strain and more than twice the collagen fibril strain. 50 The tissue strain, transferred to the mineralised collagen fibrils, decreases with age and is accompanied by an increase in AGEs. 27 Such a reduction may highlight the influence of NEG on the nanoscale mechanical behaviour of bone. Additionally, previous studies have shown that the heterogeneous distribution of properties measured by nanoindentation contribute to the energy dissipation mechanisms in bone compared to a homogeneous distribution. 51 All these observations are indicative of the contribution of nanoscale composition, structure and distribution on the fracture behaviour of bone.
Microscale bone toughness
Contribution of microstructure to fracture behaviour of bone
As outlined above, the most prominent microstructural features that influence bone strength and fracture toughness are osteons, cement lines and pores such as Haversian canals and resorption spaces.
The microstructure of bone is constantly altered as a result of remodelling. Total porosity, average pore size, 52–55 and osteon density 55–57 increase with age. Total osteonal bone area and average osteon size are found to both increase 53,54 and decrease 52,56,58 with age. Age-related changes in the microstructure may result in alterations in the fracture response of bone. Reductions in both initiation and propagation fracture toughness values were reported with an increase in porosity and osteon density, and a decrease in osteon size. 53 Finite element simulations performed on micro-computed tomography-based models also demonstrated that both initiation and propagation fracture toughness decreased with increasing cortical porosity. 59
Bone microstructure has been shown to be a significant contributor to bone strength and fracture toughness. 53,54,60–62 Microstructural features including osteons and cement lines play an important role in determining the crack growth trajectory in cortical bone 14,63–70 (Fig. 3a ). Cracks that penetrate osteons may lead to complete fracture. On the other hand, cracks that are deflected into cement lines slow down the crack propagation and increase the fracture resistance of the bone. Experimental observations showed that microcracks are predominantly located in the interstitial bone. 67,71 Longer cracks are more likely to penetrate osteons than to deflect into cement lines, whereas shorter cracks are arrested at the cement lines. 66 It has also been shown that the age of the osteons influences the crack growth trajectory. Longer cracks stops at the newly formed osteons more frequently compared to older osteons. 65 The composition of the cement line has been evaluated to gain more insight into its contribution to crack growth. Burr 72 proposed that cement lines promote crack deflection due to their low mineralisation compared to osteonal and interstitial bone. On the other hand, analyses by Skedros et al. 73 suggested that the cement lines may be more mineralised and contribute to crack initiation as a hypermineralized interface. 74 In addition to cement lines, lamellar interfaces also act as crack deflection zones because they are deficient in collagen and rich in non-collagenous protein. Furthermore, collagen orientation changes between lamellae 75 and these changes in composition and orientation promote lamellar sliding.
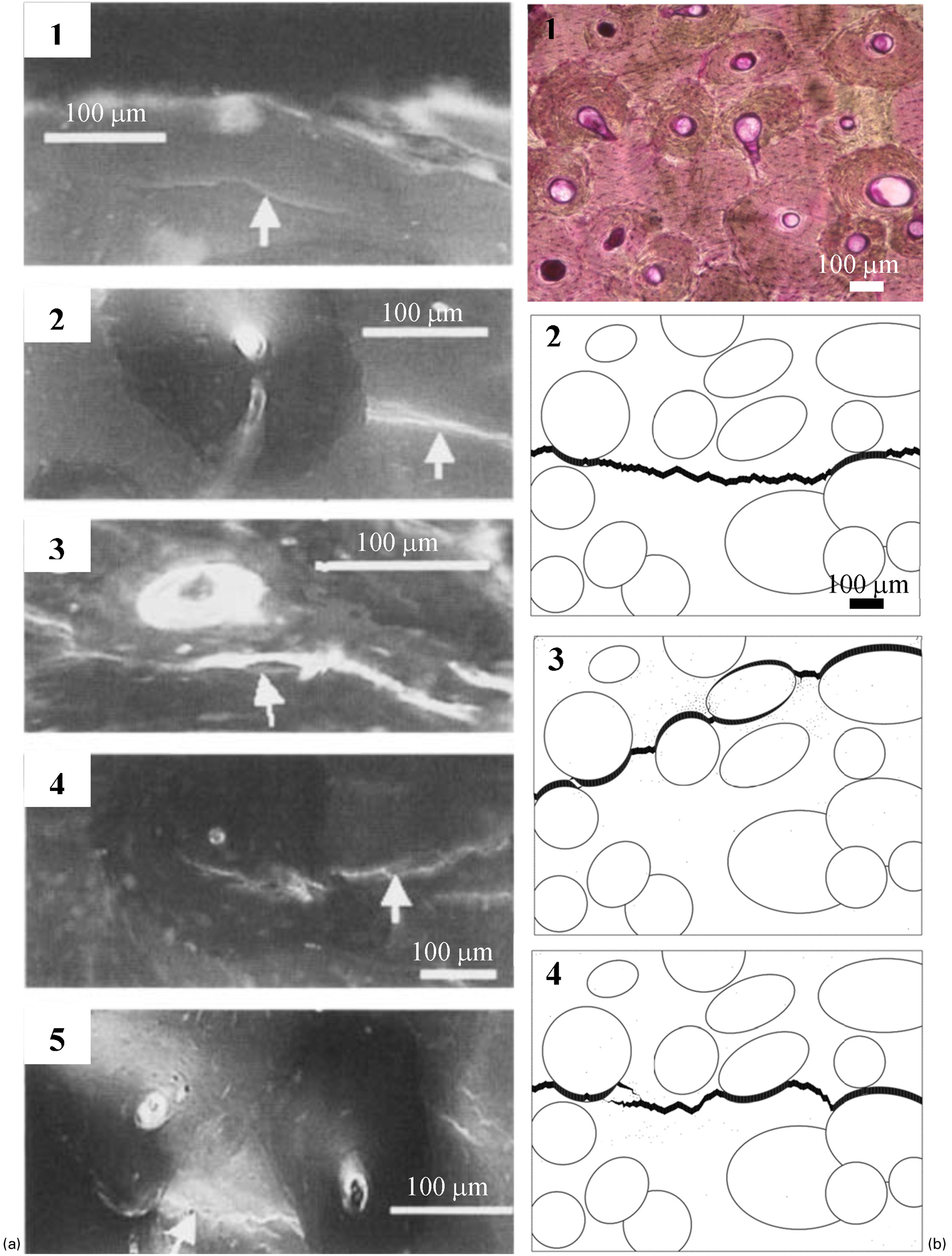
a Epifluorescence microscopy images of interaction of microcracks with osteons including (1) microcracks in interstitial bone, (2) microcrack arrest by a single osteon, (3) microcrack deflection at the cement line, (4) microcrack propagation into the osteon, and (5) microcrack arrest by two osteons crack growth (Reprinted from O’Brien et al. 66 with permission from John Wiley and Sons). Note that bone samples were subjected to a series of dyes 196 to label the microcracks formed during testing. b Finite element simulation of crack growth in a 70-year-old human bone, (1) actual bright-field microscopy image of the bone microstructure, which has undergone en bloc staining in 1% basic fuchsin. Predicted crack growth in finite element models (2) with equal properties for osteons, interstitial bone and cement lines, (3) with cement line strength reduced to half of the strength of osteonal and interstitial bone, (4) with cement line fracture toughness reduced to half of the toughness of osteonal and interstitial bone (adapted from Mischinski and Ural 79 ). Note the change in the crack growth pattern with the change in the cement line properties. The crack deflection into cement lines is most prominent in the case where the cement lines have lower strength compared to the surrounding bone
Analytical studies also evaluated the influence of cement lines on the fracture toughness of cortical bone. Such calculations predict that the interfacial toughness needs to be less than a quarter of the bone tissue toughness for cement line debonding to occur. 76 Bone tissue to cement line fracture toughness ratio increases with increasing age 77 and may adversely affect the crack deflection mechanism. This increasing elastic mismatch between the osteon and interstitial bone promotes the crack growth towards osteons and improves toughening. 78
Computational studies focussing on the effects of bone microstructure provide additional insight into some of the mechanisms that cannot be directly measured by experiments. Two recent computational studies employed fracture mechanics-based cohesive finite element modelling and explicit representation of the cement line as an interface. 79,80 These results showed that the ratio of the cement line strength to the surrounding bone played a significant role in determining the crack growth trajectory where the low cement line strength facilitated crack deflection (Fig. 3b ). Additional factors that influenced the resistance to crack growth were microcrack orientation, fracture toughness of the cement line, and microcrack length. 80 Such results highlight the importance of cement line properties in improving the fracture toughness of bone. Recent computational studies that utilise the linear elastic fracture mechanics (LEFM) 81,82 and extended finite element (XFEM) method 83–85 also support the role of osteons and cement lines in determining the crack propagation path and fracture toughness of bone.
Contribution of microdamage to fracture behaviour of bone
Microdamage forms in bone during daily activities and is repaired through the bone remodelling process. Microdamage formation plays a significant role in the fracture resistance of bone. 69,86–88 Two distinct forms of in vivo microdamage, linear microcracks and diffuse damage, have been observed in the human bone. 67,70,71,89 Linear microcracks range between tens and hundred microns and appear as sharply defined lines under bright-field microscopy (Fig. 4a ), whereas diffuse damage is a patch of submicroscopic cracks less than 1 μm in length (Fig. 4b ). 70,89–91
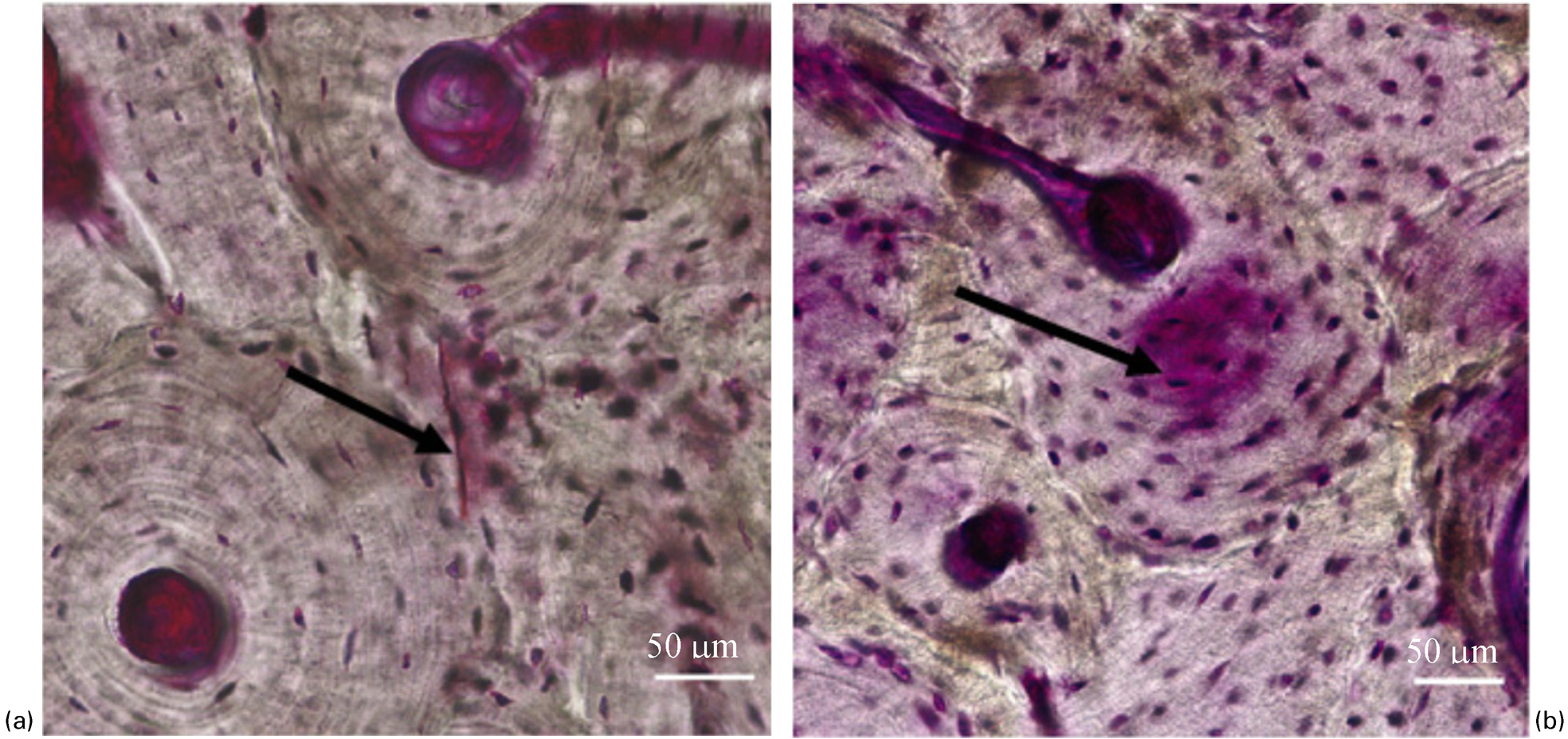
a linear microcrack and b diffuse damage (Reprinted from Diab and Vashishth 93 with permission from Elsevier). Note that the bone samples have undergone en bloc staining in 1% basic fuchsin
Linear microcracks and diffuse damage predominantly form under compressive and tensile loading, respectively. Previous studies demonstrated that diffuse damage is more effective in energy dissipation compared to linear microcracks. 92 Linear microcracks are more frequently associated with old bone whereas diffuse damage preferentially forms in young bone. 93 The reduction in the formation of diffuse damage in old bone may explain the age-related deterioration in fracture properties of bone. Linear microcracks preferentially form in the interstitial bone 67,70,93,94 and diffuse damage is predominantly located in the secondary osteonal bone. 93 Therefore, a reduction in osteon size with age may adversely affect bone fragility by promoting linear microcrack formation over diffuse damage. In addition, computational studies showed that diffuse damage formation unlocks higher order fracture toughening mechanisms such as uncracked ligament bridging and collagen fibril bridging. 36
With ageing, changes in the bone remodelling process such as a reduction in targetted remodelling, imbalance in resorption and formation, and excessive accumulation of microdamage may occur in the bone. Several studies showed that in vivo microdamage increases with age 95 and is accompanied by a reduction in the fracture toughness. 95–97 Although, these studies showed that initiation fracture toughness is lower when microcrack density is higher, another aspect that needs to be taken into account is the formation of microcracks during crack growth. Microdamage that forms in the frontal process zone and in the wake of a crack during crack propagation contributes significantly to crack growth resistance of bone. 87 Formation of microcracks is regarded as a precursor to other fracture toughening mechanisms in bone and is outlined in the next section.
Tissue-level bone toughness
Experimental and computational fracture toughness evaluation at the tissue level
Tissue-level fracture resistance of bone can be quantified using fracture toughness evaluation procedures developed for engineering materials. The most common test specimens that are utilised in bone fracture toughness measurement include the compact tension test specimen and the single-edge notched bend test specimen. 87,98–103 The fracture toughness measurements are performed on bone specimens in different orientations including longitudinal (crack growth parallel to the bone axis), transverse (crack growth perpendicular to the bone axis), and anti-plane longitudinal (crack growth in the radial direction) orientation to capture anisotropy in bone.
Initially, studies in this area focussed on a single parameter characterisation using stress intensity factor (K c) and critical energy release rate (G c) at crack initiation. Fracture toughness can be measured in three different modes including opening (Mode I), in-plane shear (Mode II) and anti-plane shear (Mode III) loading. The early critical stress intensity measurements carried out on bone focussed on the measurement of K c in the opening mode. The measurements on human femur and tibia (27–90 years-old) yielded values from 1 to 5 MPa m1/2 in the longitudinal direction, 26,87,99,100,102,104–107 0·18 to 2·21 MPa m1/2 in the anti-plane longitudinal direction 104,108 and 1 to 6·7 MPa m1/2 in the transverse direction 69,98,99,105,109 (Table 1). In addition, average values of 400 and 520 N m−1 were obtained for Mode I G c in the human tibia and femur. 110 Taken together, these results show that the initiation toughness varies with anatomical site and orientation and it is highest in the transverse direction, which is the clinically relevant fracture direction.
Summary of initiation and propagation fracture toughness measurements on human cortical bone. Note that CT = compact tension specimen, SENB = single-edge notched bend specimen, CS = compact shear specimen, SCS = single layer compact sandwich specimen, AFPB = asymmetric four-point bending specimen
*Crack growth<100 μm.
# K c (MPa m1/2), G c (N m−1).
The protocol described above determines the fracture resistance at the point of crack initiation. However, depending on the properties of a material fracture resistance may change as the crack length increases. In order to evaluate the variation of fracture resistance of bone with increasing crack length, continuous measurement of the stress intensity factor and crack length were undertaken to determine the resistance curve (R-curve) of bone. 26,87 The experimental data showed that toughness of bone increases with crack growth in all orientations. 26,87,99,104,108,109 Experimental studies demonstrated a linear relationship between the crack length and fracture toughness (Fig. 5a ). The slope of the R-curve defines the effectiveness of the toughening mechanism where a larger slope corresponds to a higher propagation fracture toughness.
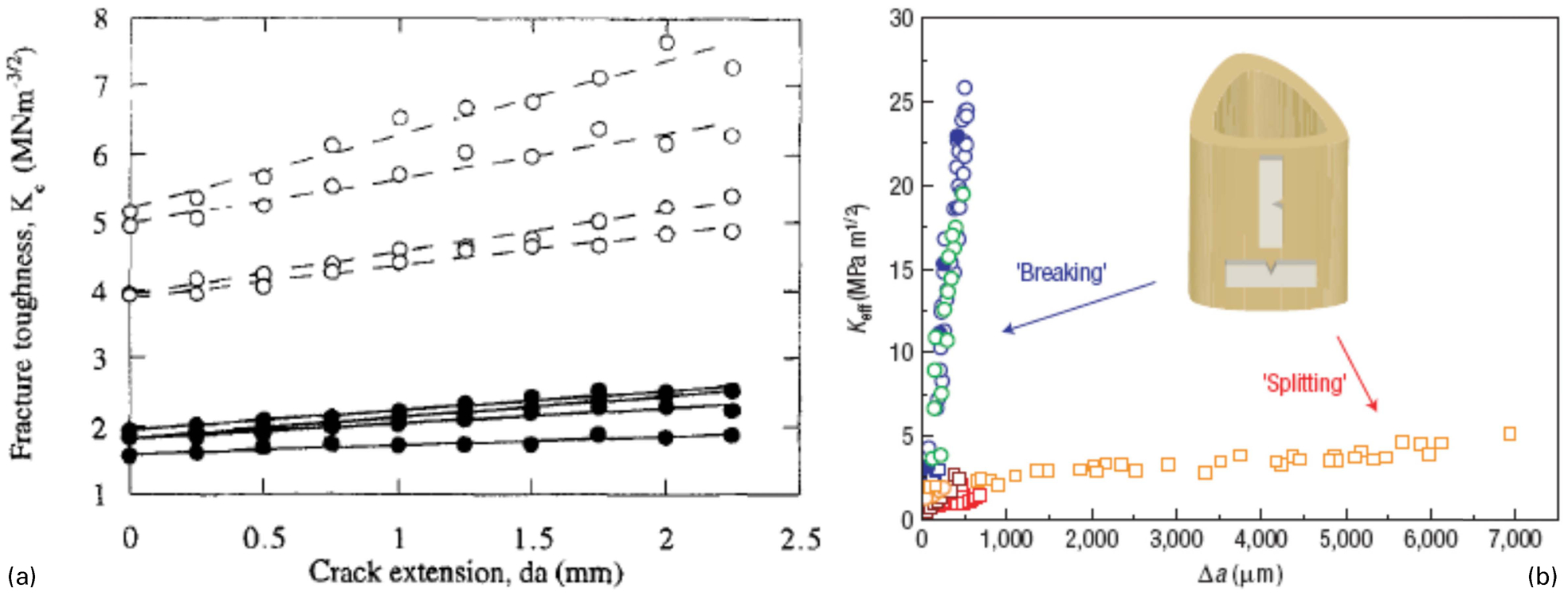
a Increasing fracture toughness with increasing crack length defining the fracture resistance curves (R-curve) in human (solid circles) and bovine (hollow circles) cortical bone (Reprinted from Vashishth et al. 87 with permission from Elsevier) b R-curves for human cortical bone in transverse (breaking) and longitudinal (splitting) orientation (Reprinted from Koester et al. 99 with permission from Macmillan Publishers Ltd: Nature Materials)
Initiation and propagation toughness represent different fracture processes in bone. 111 Physiological loading leads to the creation of in vivo microcracks in bone, which may be repaired by targetted bone remodelling. 112 Therefore, propagation of these existing microcracks to form complete fracture is the critical measure of toughness that truly reflects the fracture resistance of bone. The experimental measurements showed that slope of the R-curve ranged between 0·01 and 1·05 MPa m1/2 mm−1 in the longitudinal direction, 26,87,99,104 0 and 2·38 MPa m1/2 mm−1 in the anti-plane longitudinal direction 108 and 24 and 47 MPa m1/2 mm−1 in the transverse direction in human cortical bone (Table 1). 99,108,109 These results show that the most effective toughening occurs when the crack is growing in the transverse direction resulting in a fracture toughness of 25 MPa m1/2 within 500 μm. In a recent study, the transverse and longitudinal fracture toughening mechanisms at short crack lengths were measured showing that the stress intensity factor started from the same value, around 1 MPa m1/2, in both transverse and longitudinal directions but with crack propagation fracture toughness increased to 25 MPa m1/2 for the transverse direction whereas, in the longitudinal direction, fracture toughness values remained less than 5 MPa m1/2 (Fig. 5b ). This indicates that the energy associated with driving the crack is five times higher in the transverse direction compared to the longitudinal direction. 99 These results show that orientation plays a significant role in the fracture toughness of bone. Both initiation and propagation fracture toughness were higher in the transverse direction. The measurements demonstrated a particularly drastic difference in the propagation fracture toughness, which is the critical parameter that defines the true fracture resistance of bone. This significant difference is attributed to crack deflection and twisting that occurs at the cement line interfaces. In the longitudinal orientation because the crack propagates parallel to the osteons it does not get deflected by the cement lines and the toughening mechanisms are only restricted to microcracking and uncracked ligament bridging.
Most of the studies on fracture toughness measurement in bone have focussed on the opening mode. However, bone does not only experience pure tensile loading but undergoes multiaxial loading. In order to understand the response of bone under different types of loading, Mode II fracture toughness was also investigated. 74,106,110,113 The experimental measurements showed that for longitudinal crack growth initiation fracture toughness is higher under shear than the opening mode. 106,110 On the other hand, pure shear led to lower initiation fracture toughness than tensile loading under transverse crack growth. 113 Furthermore, the measurements of propagation fracture toughness under mixed-mode loading showed that fracture resistance increases with increasing crack length but at a lower rate than the pure opening case for transverse specimens. 74 These results indicate that when the crack driving force and the preferred microstructural path are in the same direction, the toughening mechanisms of bone are not as effective. 74,113 Consequently, under multiaxial loading conditions which the bone experiences during daily activities, its fracture resistance may be lower than the transverse fracture toughness measured under pure tensile loading.
In addition to experimental studies, new computational modelling techniques have been developed to provide further insight into the fracture mechanisms that influence the overall fracture response of bone. The strength of computational studies comes from the additional information they provide that cannot be measured by experiments and from their ability to assess the individual influence of each factor without confounding effects. One of the recent computational techniques utilising such methods is cohesive finite element modelling. The toughening behaviour of bone reported in crack propagation tests is a result of the physical processes taking place in the vicinity of a propagating crack. Cohesive finite element modelling represents the effects of these physical processes using a traction–displacement relationship. 114 This approach has captured the toughening behaviour of human bone successfully 114 and demonstrated the influence of porosity, orientation, and loading rate on fracture toughness. 59,115,116 Additional studies that utilised cohesive finite element modelling predicted the load–displacement curve obtained from fracture tests and highlighted the importance of using non-linear models to capture the fracture mechanisms in bone. 117
In addition, XFEM was utilised to measure the fracture toughness of bone under tensile and mixed-mode loading to capture the orientation dependent crack growth observed in the experiments. 118 XFEM simulations were also successful in predicting the experimental fracture toughness measurements in different orientations 119 and in capturing the influence of porosity and material heterogeneity on fracture toughness of bone. 120
Fracture toughening mechanisms at the tissue level
Experimental studies have identified several fracture toughening mechanisms that influence bone toughness at the tissue level including microcracking, collagen fibril bridging, uncracked ligament bridging, and crack deflection/twist (Fig. 6). 45,87,98,99
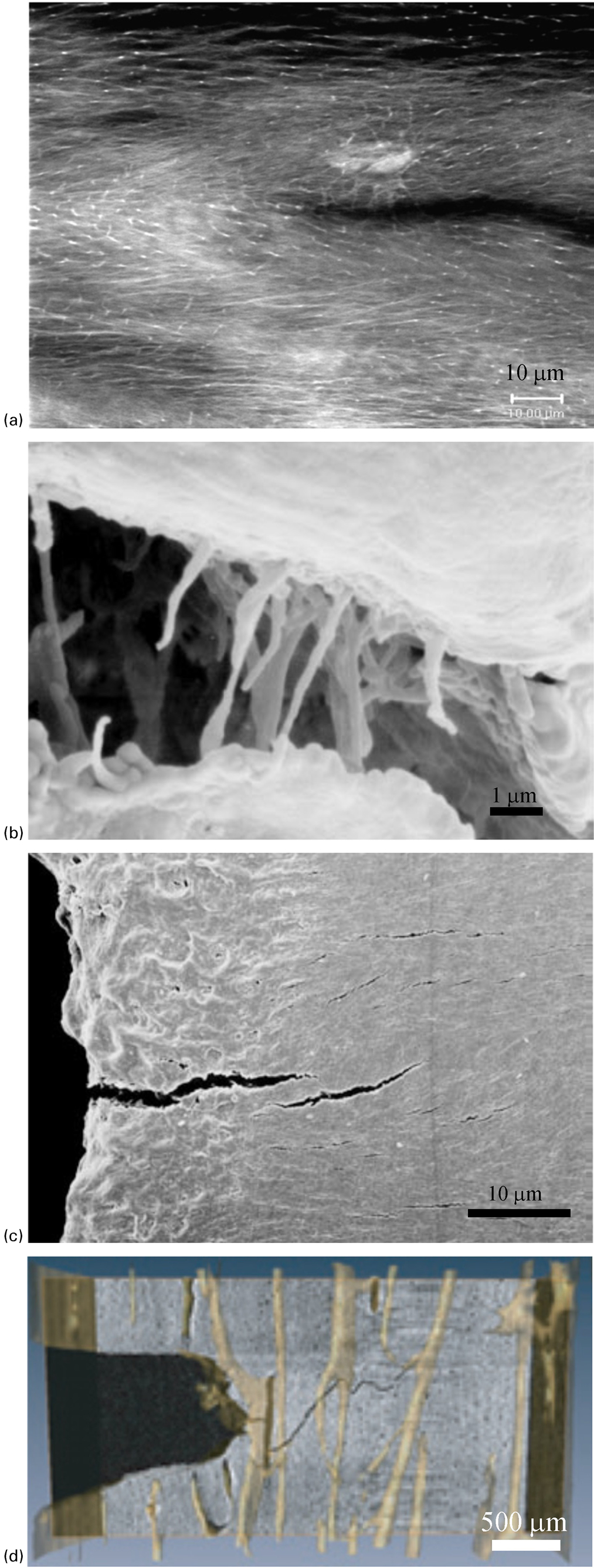
Tissue-level fracture toughening mechanisms in bone: a laser confocal image of microcracking (Reprinted from Vashishth 197 with permission from Elsevier), b scanning electron microscopy (SEM) image of collagen fibril bridging (Reprinted from Nalla et al. 98 with permission from Macmillan Publishers Ltd: Nature Materials), c SEM image of uncracked ligament bridging (Reprinted from Nalla et al. 98 with permission from Macmillan Publishers Ltd: Nature Materials), d synchrotron X-ray computed tomography image of crack deflection/twist (Reprinted from Koester et al. 99 with permission from Macmillan Publishers Ltd: Nature Materials)
Microcracking occurs in the frontal process zone ahead of the crack tip as well as in the process zone wake and contributes to the overall fracture toughness of bone through energy dissipation 87 (Fig. 6a ). In addition to its energy dissipating characteristics microcrack formation is critical for the development of other toughening mechanisms and is the precursor to the formation of crack bridging and crack deflection/twist. 45
Collagen fibrils span individual microcracks and provide stress transfer between the faces of cracks (Fig. 6b ). 98,121 The contribution of collagen fibril bridging to fracture toughness of bone has been identified by analytical models. 36,122 Although the contribution of collagen fibril bridging may not be large for a single microcrack, its effect in numerous microcracks may contribute significantly to fracture resistance of bone.
Uncracked ligament bridging is observed ahead of the crack tip particularly in longitudinal crack growth (Fig. 6c ). 98 This toughening mechanism arises as a result of microcracks that form parallel to the crack growth direction in the frontal process zone. During coalescence of microcracks uncracked sections that are formed ahead of the main crack increase the required load to propagate the crack and improve the fracture resistance of bone.
Crack deflection/twist is typically observed in transverse crack growth in which the crack growth direction is perpendicular to the direction of osteons (Fig. 6d ). 69,99 Cement lines act as boundaries to deflect and twist the crack path leading to tortuous and rough crack surfaces. Consequently, additional energy is required to cause fracture. 69,99
Influence of age-related tissue-level changes on fracture toughness of bone
Bone material properties and structure are altered with age and these lead to changes in bone’s fracture resistance. Experimental studies showed that initiation fracture toughness of various types of human cortical bone including humerus, tibia and femur decreased by 5–11% per decade in the longitudinal direction under opening mode (Fig. 7a ). 25,26,102,104,106,110 On the other hand, the propagation fracture toughness demonstrated a more substantial decrease of 12–19% per decade 26,104 and resulted in resistance curves with almost zero slope in old donors (Fig. 7b ). These results indicate a significant impairment in the toughening mechanisms in bone with age. In the transverse direction, initiation and propagation toughness decreased by 1–4%, and 3% per decade, respectively. 69,109 Both of these values are substantially less than the longitudinal direction. Similar trends in initiation and propagation fracture toughness were also observed in anti-plane longitudinal specimens. 108 Furthermore, there was a larger reduction in initiation fracture toughness under shear loading compared to the opening mode in the longitudinal direction, which ranged between 13 and 18% reduction per decade. 106,110
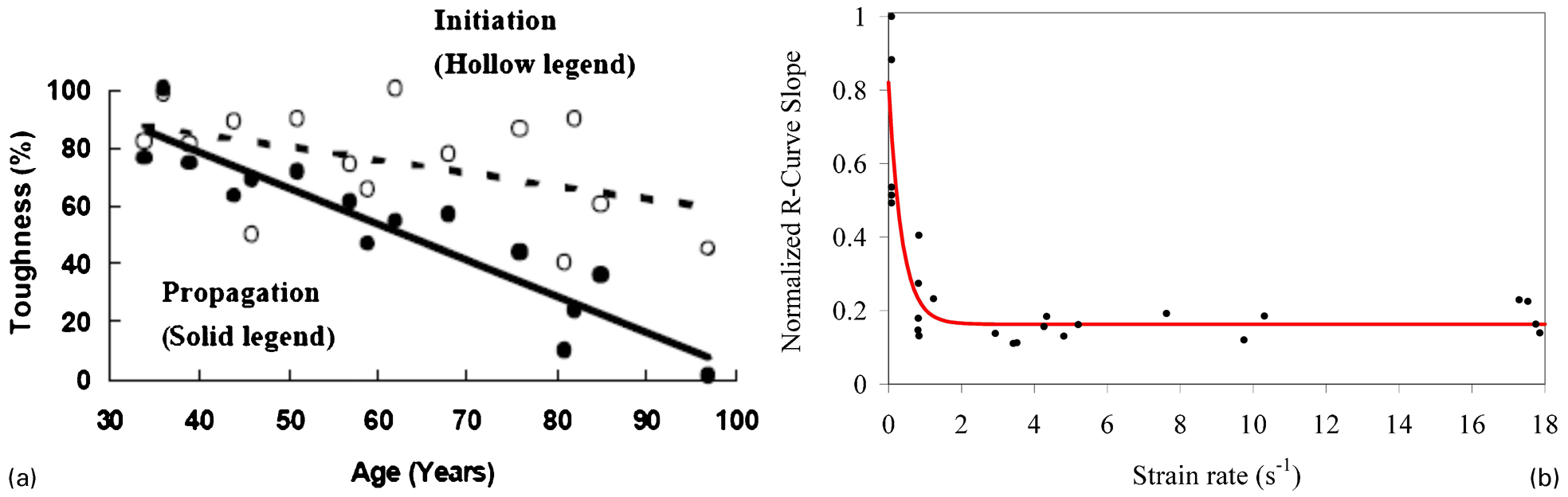
a Age-related changes in the initiation and propagation fracture toughness (R-curve slope) in human cortical bone (adapted from Vashishth et al. 26 ). b Finite element simulations on cortical bone showing the variation in the propagation fracture toughness with strain rate (Reprinted from Ural et al. 116 with permission from Elsevier)
The above observations indicate that there is a substantial reduction in the fracture resistance of bone due to the loss in the effectiveness of toughening mechanisms with age. 99,104,108 These tissue-level changes are a culmination of the changes in the lower length scales including the nano- and microscale.
Whole bone toughness
Fracture mechanisms and properties of bone at the lower scales have a direct influence on the fracture behaviour at the whole bone level. In addition to the influence of nanoscale, microscale and tissue-level properties, macroscale cortical geometry and trabecular architecture also influence the amount of load that a bone can carry before failure.
Macroscale contributors to fracture risk have been investigated using various imaging techniques 123,124 as well as fracture testing of whole bones. 125–133 Experimental and imaging data provide valuable information on the correlation of the macroscale properties to fracture risk of bone. In particular, three-dimensional imaging techniques can non-invasively capture the detailed cortical and trabecular structure and identify the contributions of macrostructure on fracture load. One of the commonly used imaging techniques is quantitative computed tomography (QCT), which can be utilised in the assessment of proximal femur and spine. This imaging technique provides only a low resolution scan and does not capture the detailed structural information of the whole bone. Consequently, high resolution peripheral computed tomography (HR-pQCT) and magnetic resonance imaging (MRI) emerged as more suitable modalities for bone imaging that can provide high resolution information on the cortical and trabecular structure of bone. 123,124,134 HR-pQCT can only be utilised at peripheral sites such as distal radius and tibia (Fig. 8a ) whereas MRI can also be utilised at other sites including proximal femur but with lower resolution compared to distal radius and tibia.

a HR-pQCT image of distal radius and b finite element mesh generated based on the HR-pQCT image (Reprinted from Ural et al. 165 with permission from Elsevier)
Age-related changes in the cortical bone geometry 52,135–137 and trabecular bone architecture have been reported. 138 These changes in geometrical and architectural properties of bone influence the amount of loading a bone can carry. There is emerging evidence that the cortical bone plays an important role in determining fracture risk. The changes in the actual area of bone as well as its spatial distribution are important in determining the mechanical response. Bone area is a significant factor in determining the resistance against tensile or compressive axial loads whereas moment of inertia and polar moment of inertia determine the bending and torsional rigidity of the bone. The geometrical properties of the radius measured by computed tomography were found to correlate with the fracture load determined by whole bone testing of the radius. 126–128,130,139,140 The cortical bone content at the distal radius shows the highest correlation with the fracture load. 127,128 In addition, the cortical area at 20% of the distal radius has been reported to be a strong predictor of the fracture load of the whole bone. 127 These results suggest that cortical bone is an important determinant of strength in the distal radius. Studies performed on other sites such as the femoral neck have also demonstrated that cortical bone thinning and increased intracortical porosity influence the strength of the bone as well as skeletal fragility. 141,142 The biomechanical role of the cortical shell in vertebrae 143 as well as the femural neck 144 can be substantial. Previous studies found cortical thinning to be the most influential factor on bone strength at these sites. 145 It was also shown that the greater cortical thickness may enable the cortical compartment to carry a higher percentage of the load on bone and may lead to lower rates of fracture. 146 A recent study demonstrated that significant bone loss occurs in the cortical bone creating a porous structure with cortical remnants 147 and this significant increase in cortical bone porosity adversely affects the macroscale structure and fracture risk.
One of the strengths of imaging techniques such as QCT, HR-pQCT and MRI is the possibility of converting these images directly to finite element models to assess the mechanical competence of the whole bone (Fig. 8b ). The majority of the studies in the literature performed strength-based analysis under axial loading 148 and evaluated the structural and mechanical properties of the distal radius and tibia. 149,150 Assessment of the correlation of bone strength with cortical and trabecular structural parameters showed that individuals with a fracture history can be predicted based on the deterioration of both the cortical and trabecular bone structural parameters. 151–157 Evaluation of the load distribution between the cortical and trabecular bone identified that the influence of both compartments on the fracture load varies with anatomical site. 123,151,158 Additionally, the effects of various disease and therapeutic interventions on the mechanical response of bone were identified. 159,160 Cortical and trabecular parameters were also successful in predicting fracture load and bone strength in the femur and spine. 129,143,161–163
Recently instead of strength-based analysis, new fracture mechanics-based assessment methods that utilise cohesive finite element modelling has been used to determine macro level effects. 164,165 The use of a fracture mechanics-based approach in place of a strength-based approach provides a more robust failure criterion by explicitly modelling the crack formation process and by integrating fracture toughness, a parameter that directly influences the bone fracture behaviour. This modelling approach also provides more realistic fall loading conditions. Such studies demonstrated the significant contribution of the cortical geometry both in idealised and HR-pQCT based models. 164–166 The results of the simulations showed a positive and significant correlation between the cortical and whole bone fracture load as well as a significant and positive correlation between the cortical bone fracture load and the cortical to whole bone fracture load ratio. These findings indicate that a more robust cortical bone structure carries a larger percentage of the whole bone fracture load and highlight the importance of the cortical compartment in fracture risk. 165
Several studies have shown that the macrostructural properties of bone can be predictors of the material level properties of the bone. 167–170 For example, compared to a wider tibia a narrower tibia was shown to be at higher risk of stress fractures as a result of both higher tissue stresses caused by the smaller size of the bone as well as its higher susceptibility to damage accumulation. 167 Fracture toughness was also found to increase linearly with increasing thickness and cortical index (ratio of cortical thickness to its width) in the human femur. 170 Studies on mouse bone showed that the morphological and tissue-level properties demonstrate co-adaptation to resist the applied physiological loading. 171–173 Taken together, these findings demonstrate a close interaction between morphological and material level properties in order to determine the fracture properties of bone.
Influence of loading rate on fracture toughness of bone
The majority of the experimental fracture toughness measurements on bone are done under quasi-static loading. However, bone is subject to a wide range of strain rates varying from daily physical activities to walking and running (0·004–0·05 s−1). 174,175 In addition to daily activities traumatic events such as accidents or falls can increase the strain rate experienced by bone to as high as 25 s−1. 176
Experimental studies indicate that the energy absorption and initiation fracture toughness show an initial increase with increasing strain rate, which then change to a decreasing trend with a further increase in the strain rate. 177–188 Investigations of the propagation toughness also showed similar trends with lower propagation toughness under dynamic loading compared to quasi-static loading. 188 The reduction in propagation toughness was larger than that in initiation toughness under higher loading rates. 188
Computational studies were also performed to investigate the influence of strain rate on fracture toughness of bone. One of the studies which employed cohesive finite element modelling demonstrated a steep reduction in propagation toughness just above the strain rates that correspond to physiological range 116 (Fig. 7b ). This result indicates that the effectiveness of the toughening mechanisms that are active under physiological loading decreases significantly under traumatic loading. Other dynamic finite element analyses of bone have also shown that the critical energy release rate decreases with increasing strain rate. 185,189 The effect of strain rate on the fracture load in whole bone was also studied and it demonstrated a significant reduction in fracture load above the physiological loading threshold. 190
Conclusions and future directions
This review summarised the current state of knowledge in hierarchical fracture processes in bone ranging from nano- to macroscale and their contribution to overall skeletal fragility.
At the nanoscale, the primary fracture mechanisms that contribute to energy dissipation are identified as molecular uncoiling, dilatational band formation, and fibril matrix shearing. These fracture mechanisms are directly influenced by the amount and composition of nanoscale components of bone such as collagen, non-collagenous proteins, and mineral phase. The age-related changes in the nanoscale composition of bone including accumulation of AGEs, increasing mineralisation, and reduction in non-collagenous protein have negative effects on the nanoscale fracture mechanisms and deteriorate the overall fracture toughness of bone.
At the microscale, osteons, cement lines and porosity have a significant influence on bone toughness as they affect the crack propagation path through crack deflection into cement lines and penetration of osteons. The amount and morphology of microdamage is another important contributor to energy dissipation at the microscale with diffuse damage being a more effective microdamage morphology for fracture toughness compared to linear microcracks. The age-related changes in the microstructure of bone including increased porosity, reduced osteon size, increased osteon density, and higher proportion of linear microcrack formation over diffuse damage adversely affect the microscale fracture mechanisms and result in a reduction in overall fracture toughness of bone.
The most prominent toughening mechanisms identified at the tissue level are crack bridging, crack deflection/twisting, microcracking, and uncracked ligament bridging. These mechanisms result in higher fracture resistance with increasing crack length. The effectiveness of all the tissue level toughening mechanisms deteriorates with age due to the changes in the lower scale levels. Loss of integrity in the bone macrostructure also adversely influences the fracture behaviour of bone. At the macroscale, morphological parameters including trabecular and cortical bone geometrical and architectural properties contribute to fracture risk and have been found to correlate with the fracture load at different bone sites including the distal radius, tibia and femur.
These observations at multiple scale levels show that the fracture occurrence in bone is a complex process that occurs as a result of the contribution of different processes from nano- to macroscale. Despite the wealth of knowledge of fracture mechanisms at different scale levels in bone, as outlined in this review paper, there are still many unanswered questions. Particularly, the processes at the nanoscale associated with collagen, mineral, and non-collagenous proteins are not fully understood. The causes for the compositional changes at the nanoscale, which leads to the impairment of fracture resisting mechanisms in bone needs to be investigated in more detail in order to obtain a deeper understanding of the processes occurring at this level. The individual influences of non-enzymatic and enzymatic cross-linking in collagen fibrils, the amount, connectivity and structure of the collagen and non-collagenous proteins, and the distribution, amount and structure of minerals on the nanoscale fracture processes need to be identified more clearly in order to develop strategies for preventing or reversing such changes. This can be achieved by new experimental techniques that can separate out the effects of each component as well as novel computational methods that can aid the evaluation process using parametric studies. Although, several important fracture mechanisms have already been identified at the nanoscale, further research is needed to unveil other unknown mechanisms that may contribute to fracture toughness. Another new direction that may improve the understanding of bone behaviour is the identification of subnanoscale fracture mechanisms, features, and constituents of bone. This will require the advancement of existing measurement technologies and development of new visualisation techniques with higher resolution. The new knowledge gained in nanoscale processes will be significant in providing further insight into their effects on fracture mechanisms at higher scale levels. The influence of changes in local material properties such as reduced heterogeneity and increased brittleness have been speculated to significantly affect the microscale fracture toughening mechanisms. Additional research in this area is needed to provide direct evidence of these mechanisms and to quantify the material composition required for effective microscale toughening in bone.
At the macroscale, one of the most important unanswered questions is the relative role of trabecular and cortical bone in determining fracture risk. Most of the studies have given emphasis to the contribution of trabecular bone to the fracture resistance of bone. On the other hand, recent studies have identified the possible importance of cortical bone in determining bone fracture load. There is already some progress in this area through the availability of advanced imaging techniques such as HR-pQCT and MRI that can measure macroscale properties including cortical bone amount and distribution and trabecular architecture with high accuracy. These imaging techniques are currently used only as research tools. Although, many of these studies have found correlation between the fracture load and cortical and trabecular structural properties of bone, there is no consensus on which parameters have the largest impact on fracture response of bone and should be included in fracture risk assessment. As a result, further research needs to be done to clearly identify the most important macroscale contributors to fracture risk and to translate these imaging techniques to the clinical setting.
The ultimate goal in gaining a thorough understanding of fracture mechanisms is to non-invasively identify individuals under fracture risk and develop new therapeutic approaches or other strategies to prevent future fractures. The review of the literature highlights the inadequacy of the current clinical fracture risk assessment and therapeutic intervention approaches that target only bone mineral density. Bone mineral density reflects only one aspect of the changes occurring in bone due to age or disease. Additional measures that represent overall bone quality other than bone mineral density need to be incorporated for reliable assessment of skeletal fragility and development of target-specific therapeutic approaches. There are currently two main issues that need to be addressed to include bone quality measures in clinical assessment of fracture risk. The first one is the lack of availability of non-invasive evaluation methods for all bone quality measures. Although patient-specific geometrical and architectural properties of bone can be assessed non-invasively using advanced imaging techniques, non-invasive evaluation of the mineral and organic matrix properties of bone is currently not possible. The only way to assess patient-specific material composition information is through bone biopsies which, in their current form are large and hence limited for routine application of this approach in a clinical setting. Therefore, an important future study direction is the identification of biomarkers that can non-invasively measure the nanoscale compositional state of the bone, which directly affects material level properties. The development of biomarkers that can evaluate the material quality of bone without invasive procedures will provide a significant improvement in fracture risk assessment. Combining the information obtained from advanced imaging and biomarkers will provide a more complete picture of bone quality including structural and material level properties.
The second outstanding issue in using bone quality measures in fracture risk assessment is the lack of clear identification of the most influential bone quality measures and their clinical thresholds for high fracture risk. The studies in the literature have identified association of various processes and constituents with fracture mechanisms at different scale levels but have not conclusively determined the factors that can be reliably used in fracture risk assessment. The application of the accumulated current knowledge in this area to minimally- or non-invasive assessment of fracture risk is a major new direction in bone research. The establishment of the missing link between the existing knowledge and prediction of fracture risk through new experimental and computational approaches will be invaluable in improving patient-specific fracture risk assessment.
Effective incorporation of computational methods such as finite element modelling in fracture risk assessment may provide unique information that may not be available through just experimental evaluation alone. Finite element modelling is a powerful tool that can measure the independent effect of different parameters one at a time and can eliminate the confounding effects that may exist in testing of biological materials that are not associated with the measured variable. A wider use of computational modelling to support experimental measurements is expected to be beneficial to advance the understanding of underlying material level changes and fracture mechanisms in bone. Development of accurate and detailed computational tools may also reduce the high cost associated with experimental research studies. Additional information gained from computational methods may also complement the information obtained from imaging and provide benefits in clinical fracture risk assessment. The challenges in the incorporation of these methods in clinical fracture risk assessment are the lack of mechanistic understanding of fracture by end users and the computational power that may be required for detailed simulations. Further research in this area can focus on the development of simplified modelling techniques that can be utilised in a clinical setting and can be performed on a single desktop computer.
As an alternative approach, fracture risk prediction tools such as WHO Fracture Risk Assessment Tool (FRAX) has been developed to combine measurable clinical risk factors including age, sex, body mass index, family history of fracture, and life style choices 191 with BMD to improve the prediction of osteoporotic fractures based on BMD alone. 192 The prediction tools such as FRAX provide a probability of 5–10-year fracture risk based on clinical risk factors with or without the knowledge of BMD 193,194 and are developed to serve as enhanced, cost-effective assessment and intervention guidelines for clinicians. Although, validation studies performed on FRAX and other tools based on clinical risk factors showed comparable and relatively successful fracture risk predictions, their effectiveness in selecting patients for treatment and preventing fracture still needs to be evaluated. 193
Another important research area is the assessment of the influence of current osteoporosis drugs, particularly bisphosphonates, on bone quality. Although positive effects of these drugs in increasing BMD and reducing fracture risk have been demonstrated in case studies, a thorough understanding of the material level changes that occur at different hierarchical levels of bone has not been established. Improving the understanding of these changes will elucidate the mechanical basis of the benefits of osteoporosis drugs and also will help understand conditions such as atypical femoral fracture that might be occurring due to prolonged use of bisphosphonates. Additional information that will be gained by detailed research in this area will provide important information that will help compare the effectiveness of different drugs and will provide insight into the outcomes in preclinical or clinical trials.
In addition to the development of better fracture risk assessment methods, the new knowledge gained on the hierarchical structure of bone should lead to the development of new drugs that target the nano- and microscale processes, composition and structure in bone instead of just bone mineral density. These new therapeutic developments may provide greater improvements in the prevention of bone fracture and reverse the effects of osteoporosis.
Footnotes
Acknowledgement
Research reported in this publication was supported by the National Institute on Aging of the National Institutes of Health under award number AG020618 and National Institute of Arthritis and Musculoskeletal and Skin Diseases under award number AR49635. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.