
Abstract
The clinical success of many orthopaedic implants relies on good integration between the implant and adjacent bone. As stabilising bone grows not only to the implant, but from it, the quick adhesion of bone forming cells called osteoblasts, their appropriate differentiation and ability to form mineralised bone are vital to achieve a good clinical outcome. Surface free energy can be thought of as a measure of the ‘unsatisfied bond energy’ resulting from ‘dangling bonds’ exposed at a material's surface. This unsatisfied bond energy affects protein adsorption and cell attachment, and thus controls the early stages of cell–biomaterial interactions and ultimately implant fixation. When water, proteins, or cells approach a surface, their surface domains align to minimise the overall surface free energy of the interface. Determining these interactions, however, is not simple. While contact angle measurements on flat surfaces can predict some surface free energy-related interactions, this is not the case when surface topography is modified. Here, the authors review how surface free energy can be altered on self-assembled monolayers, polymers, metals and ceramics and clarify the differences between measurements of surface free energy and wettability. The authors also review how surface free energy affects protein interactions and osteoblast behaviour. The result is a clearer understanding of the effect of surface free energy on cell behaviour and an unambiguous need for further studies that isolate such effects.
Introduction
Successful orthopaedic implant osseointegration relies on the quick and efficient formation of bone tissue at an implant surface. When biological fluids come in contact with an artificial material, water interactions, protein adsorption, and cell attachment are governed by the surface free energy of the material. These early interactions with the surface play a fundamental role in determining cell adhesion, differentiation and ultimately tissue formation at the interface. Understanding how surface free energy affects the interactions of a surface with the biological milieu may allow for the rational design of biomaterials. Rational design, or creating biomaterials prospectively with surface properties that promote particular cell responses, would be far more efficient than testing all possible materials retrospectively. In short, developing a set of rules that describe how various properties of materials' surfaces govern protein interactions and thus the resulting cell response, may allow for the design of surfaces that promote favourable interactions with proteins, cell adhesion and tissue-appropriate differentiation. Here, the authors review how surface free energy influences biological interactions with biomaterials and discuss how the field can move forward to design surfaces that promote favourable biological responses, particularly those that will promote implant osseointegration.
Biomaterials in joint replacement
Damage to the articulating surfaces of the joint, particularly resulting from osteo- and rheumatoid arthritis, is painful and debilitating. Joint pain is one of the most common reasons people report for visiting their general practitioner, and the US Center for Disease Control estimates that direct and indirect costs related to arthritis in the US are more than US $128bn annually. 1 Surgical interventions such as microfracture and autologous chondrocyte implantation are often employed to try to mitigate joint degradation and the emerging fields of tissue engineering and regenerative medicine aim to create cell-based therapies to prevent or reverse joint disease. 2,3 Nevertheless, the gold standard treatment applied to many patients for painful joint degradation in the knee, hip and finger is to replace the bearing surfaces with a metallic/polyethylene implant in an arthroplasty procedure. The American Academy of Orthopaedic Surgeons reports that over 400 000 knee and hip replacements procedures are performed annually in the US and this number is expected to climb as the population ages.
Total joint arthroplasties have historically relied on fixing metallic implants to the underlying bone with a poly(methylmethacrylate) cement. Despite the success of these treatments, it is widely accepted that the cement provides a weak interface and contributes to loosening and eventual failure of implants in the long term. 4 Subsequent revision surgeries of failed cemented implants are complicated by the need to remove residual cement and carry the risk that there is insufficient remaining bone to stabilise a new implant. 5 As the population ages and increasingly expects to remain active late into life, cementless (or uncemented) implants, which are stabilised by bone bonding directly to the implant surface, are increasingly preferred, and particularly so in younger patients. Despite their advantages, however, retrospective studies have found that cementless implants generally have not performed better than their cemented counterparts, and some studies even report worse patient outcomes. 6–8
Cementless joint arthroplasties are reported to fail for a number of reasons, but one of the most common is as a result of aseptic loosening, a process by which micromotion between the implant and the bone eventually leads to instability of the implant and the need for revision (Fig. 1). Some 4·2% of cementless total knee implants fail because of this mechanism. 7 When a strong bone–implant bond does not form soon after surgical placement, micromotion prevents the growth and mineralisation of bone at the implant surface, which further contributes to micromotion, and thus further prevents bone formation. In short, poor bony fixation results from micromotion and leads to the ingrowth of fibrous tissue instead of bone at the implant/bone interface. 9–11 This process can be exacerbated by the presence of wear particles from the polyethylene bearing surface, which can cause inflammation and interfere in the cell-mediated process of bone fixation. The other common failure mode for orthopaedic implants is infection, which affects between 0·8 and 4·0% of total knee arthroplasties. 7 When an implant is first placed in the joint, there is a virtual ‘race to the surface’ between microorganisms and cells. If microorganisms arrive at the surface first, the biofilm that they create can preclude later cell adhesion and the device may have to be removed. 12
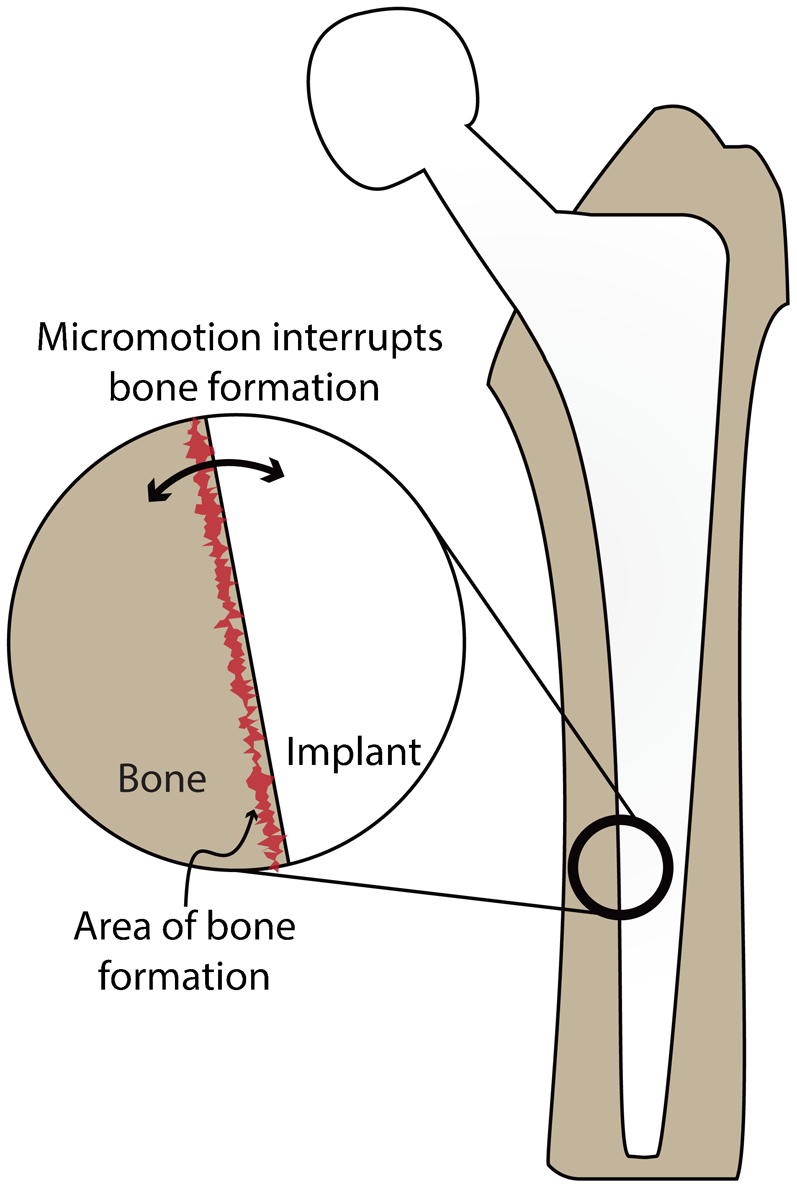
Schematic of a typical cementless implant used in hip replacement surgery. Although total joint replacements can fail for a number of reasons, including infection and poor surgical placement, among others, aseptic loosening remains one of the most common. The inset shows how micromotion at the bone–implant interface can prevent the formation of robust mineralised bone leading to aseptic loosening and eventually the need for surgical revision
To avoid such complications, the early formation of a robust bond between the implant surface and the surrounding bone is essential for long term success. In short, one of the most important factors in the success of a cementless total joint arthroplasty is initial implant stability, 12 which directly relies upon early and robust bone growth at the implant–tissue interface. Bone growth rates from the implant surface towards bone are up to 30% faster than those from the bone towards the implant. 13 As a result, one of the most promising paths for improved fixation is to accelerate the onset and rate of early cell adhesion and bone growth at the implant surface. 14,15 To accomplish this, a more complete understanding of how material surfaces' properties affect protein adsorption and cell response is integral.
Modifying biomaterial surfaces
During the earliest stages of osseointegration, proteins adsorb on the implant surface. This process is followed by osteoblast adhesion and maturation. 16 Physico-chemical signals from the surface resulting from its inherent chemistry and topography govern and drive these processes. Three methods are commonly employed to modify biomaterial surfaces: (1) physico-chemical changes, (2) changes in surface topography, and (3) biochemical changes. Changes in surface topography at the macro-, micro- and nano-scales can impact properties such as surface free energy and surface chemistry, and consequently will affect protein attachment, conformation and activity and thus how cells interact with a surface. The effects of topographical changes are complicated and reflect the variety of means by which the topography can be altered and the interested reader is referred to excellent reviews on this topic. 17–19 Biochemical changes rely on coating the surface with delicate molecules such as peptides or proteins. Osteogenic coatings may include alkaline phosphatase or bone morphogenetic proteins, which mediate mineralisation and encourage bone cell differentiation, respectively. Although these coatings show promise, 20,21 they can be delicate and may change the requirements for materials handling, FDA approval, and surgical techniques, and so their path to translation is more fraught.
Physico-chemical changes, which include changes in chemistry and charge at the surface, both of which are encompassed in the surface free energy of the material, will be the primary focus of this review. These changes modify the chemical nature of the surface and can affect the adsorption and attachment of proteins and cells, respectively. These changes also modify signals that are given to subsequent layers. Here, the authors will focus on the influence of surface free energy, independent of topography on cell–biomaterial interactions.
A materials science perspective on surface free energy and wettability
Surface free energy is the increase in energy associated with taking an atom from the bulk of a material and placing it at the surface. When that surface is created, the atoms at the newly exposed surface have fewer nearest neighbours than those same atoms would have had in the bulk, this is known as coordinative unsaturation of the bonds. Coordinatively unsaturated atoms at the surface have a higher energy state than atoms whose bonds are fully saturated in the bulk. Thus, surface free energy is a measure of the increase in energy created at the surface of the material by the type and number of dangling bonds present. The types of bonds can be loosely divided into primary- (ionic, covalent, and metallic) and secondary- (van der Waals) type bonds. If the dangling bonds are largely van der Waals in type, the surface free energy will have a predominantly non-polar nature. If the dangling bonds are of primarily ionic- and covalent-type, it will result in significant Lewis acid and base contributions to the total surface free energy. Most surfaces are composed of a combination of all three of these components, making their interactions with liquids and therefore the biological environment extremely complex.
Contact angle measurements
Contact angle measurements using a range of well-characterised liquids on solid surfaces are the most common method for measuring surface free energy in biomaterials research. Generally, contact angle measurements can be used to characterise these surfaces in two ways. On smooth surfaces, contact angle measurements can be used to determine the components of surface free energy. 22–26 It should be noted, however, that ‘smooth’ is defined by a combination of the accuracy with which contact angle can be measured by an instrument and the surface free energy of the system being measured. As a result, materials described as ‘smooth’ can have variable surface roughnesses. Conversely, on rough surfaces where the surface free energy is known, information about the topography of the surface can be extrapolated from contact angle measurements. Both of these pieces of information can be obtained because the equilibrium state for a liquid on a surface is dependent on both the thermodynamic equilibrium at interphase interfaces and the total length/area of the phases in contact. Together, these allow for the calculation of surface free energy from droplet geometry, or surface geometry from surface free energy. 27–29 This result highlights that both changes in topography and surface free energy can affect contact angle measurements and care must be taken to distinguish between them. While surface free energy is a materials' property that depends on surface structure and composition and is therefore limited in range for a given material, topography is not similarly restricted and can be modified dramatically by processing and fabrication techniques. As a result of the materials' constraints of surface free energy and the wide range of topographies that can be obtained using currently available fabrication techniques, surfaces fabricated from the same material have been observed to exhibit contact angles ranging from the superhydrophilic to superhydrophobic. 30,31
Surface roughness and wettability
Wenzel
28
and Cassie and Baxter
27
have described the role of surface roughness in wettability. Unlike surface free energy, which is determined by a material's inherent structure, wettability can be defined quite simply as a measure of the ability of a liquid to contact a surface. Wenzel described wetting of a rough surface when the liquid completely wets all the features underneath the droplet. In this fully wetting condition, contact angle was found to be dependent on the surface roughness as measured by the tortuosity, or increase in area, of the surface. The direct relationship between roughness and the change in wettability is described by equation (1). θw is the observed liquid contact angle, or wettability of the surface, θ is the contact angle for the flat surface, and R is defined as the tortuosity of the surface (real area divided by projected area)
28
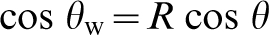
One of the most important features of this equation is that it predicts that surfaces which are intrinsically hydrophilic (contact angles less than 90°) will become more hydrophilic when roughened, and hydrophobic materials (contact angles greater than 90°) will become more hydrophobic with increasing surface roughness (Fig. 2). This type of wetting is most common when the features on the rough surface have relatively low aspect ratios (rolling hills as opposed to sharp mountain peaks).
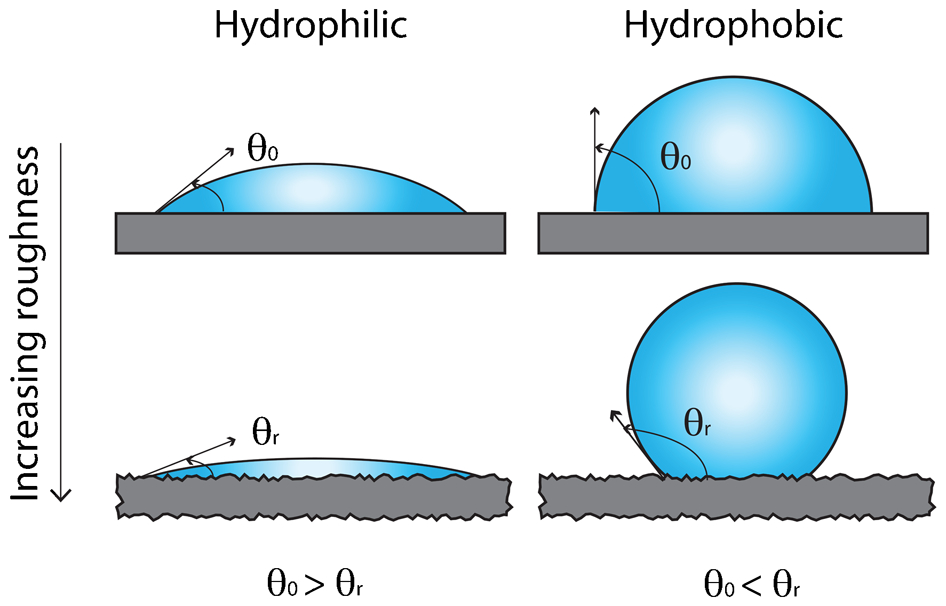
Diagram demonstrating the effect of roughness on contact angle when the liquid fully wets the surface. The right side shows the effect of roughening on a hydrophobic surface (liquid contact angle greater than 90° for the flat surface). In this case, roughening increases the observed contact angle. The left side shows the effect of roughening on a hydrophilic surface (liquid contact angle less than 90° for the flat surface). In this case roughening decreases the observed contact angle
In contrast to Wenzel, Cassie and Baxter described the effects of very high tortuosity roughness on surfaces where droplets do not fully wet the surface. In this case, the roughness of the surface is so great that instead of wetting the surface, the water sits on top of the features that create the surface roughness (Fig. 3). This, in effect, creates an air–solid composite surface below the wetting liquid. As the contact angle of water suspended in air is 180°, the resulting effect is superhydrophobicity, materials with contact angles in excess of 140°. 27,30–35 This is the wetting condition responsible for the hydrophobic behaviour of the lotus leaf and many other biological examples of superhydrophobicity. 33 This phenomenon is capable of inducing superhydrophobic behaviour in structures made of hydrophilic materials.
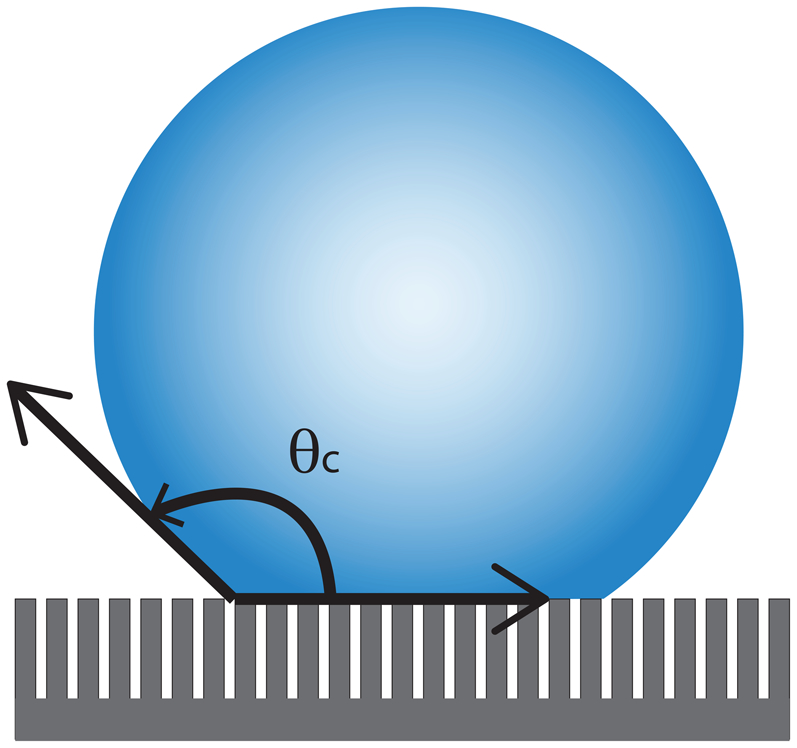
Diagram detailing the role of extreme roughness on observed contact angle (θ c). In this case, it is possible for the liquid droplet to sit on top of the surface features leaving an air–solid composite beneath the droplet instead of wetting the tortuous surface. As the air forms a 180° contact angle with water, the composite air/solid surface below the droplet can produce contact angles that approach 180° as the amount of solid in contact with the droplet is reduced
When it comes to understanding the wetting phenomena of liquids and biological materials with solid surfaces, care must be taken. Although surface free energy and topography are independent of one another, their measurements by contact angle are intimately linked. In order to make measurements of surface free energy using contact angle measurements the surface topography must be known. Once a smooth rigid surface of a material is obtained, the surface free energy can, and should, be determined by the measurement of contact angles using several liquids. The resulting equilibrium droplet shape is defined by Young's equation (equation (2)
28,29,36
), which describes a force and equivalent energy balance between the interacting surfaces (liquid, solid, and vapour)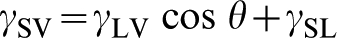
Here, γSV is the solid–vapour surface free energy, γLV is the liquid–vapour surface tension, and γSL is the solid–liquid surface free energy. This equation assumes that the solid surface is flat and rigid, and that the system is in equilibrium. This equation can be extended to include the non-polar and acid/base components of surface free energy of a material (non-polar γLW, Lewis acid γ+, and Lewis base γ−), which can also be determined using a series of contact angle measurements.
22,26
These measurements should be made on a flat, rigid surface with a minimum of three liquids whose non-polar, acid, and base surface free energies are known. The choice of base liquids has been well studied by Della Volpe and van Oss and the reader is encouraged to refer to their work when selecting base liquids.
23–25,37
The measured values of contact angle and the surface free energy values from each of the three liquids can then be entered into the following equation
22
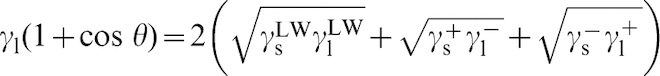
In this equation, γl is the total surface tension of the liquid (this is the same as γLV in Young's equation). The total surface free energy of each phase is then made up of γLW (non-polar), γ+ (Lewis acid), and γ− (Lewis base) components of surface free energy. In each case, the subscripts s and l denote the solid and liquid phases in contact with the gas, respectively. By solving this equation simultaneously for all three liquids, the three unknown surface energy values for the solid can be determined. Monte Carlo methods can then be used to determine the error in the surface free energy measurements. 38
High and low surface energy surfaces
As can be seen from the previous section, the surface free energy of a material is independent of surface topography. De Gennes devised a method to divide materials' surfaces into two general categories: high energy and low energy. 39 ‘High-energy’ surfaces were described as being composed of materials that are metallically, covalently, or ionically bonded, while ‘low-energy’ surfaces are defined as being largely composed of van der Waals-type bonding and often include molecular crystals and polymers. High energy surfaces include metals and oxides and have surface energies ranging from approximately 500 to 5000 mN m−1, while low energy surfaces, including molecular crystals and plastics, have surface energies in the range of 5–50 mN m−1. Both high and low energy surfaces have surface free energies containing non-polar, Lewis acid, and Lewis base components. 22,25
High-energy surfaces such as metals and metal oxides are most commonly used in biological implants and will be discussed here. The chemisorption on these surfaces is different from that on low energy surfaces primarily because of the presence of considerable amounts of Lewis acid and base character. The large amount of acid and base surface free energy is the result of ionic character present in the dangling material bonds at the surface. The ionic nature of the bonds results in the formation of cation and anion sites on the surface leading to strong acid/base components of surface free energy that are not present in ‘low-energy’ surfaces. At these charged surface sites, it is possible for the ions to interact with molecules at the surface through ion–dipole attractions and electronic orbital overlap. The strong acid/base surface energy of these surfaces can result in interactions with adsorbates on the surface. The most common example is water molecules, which are susceptible to the dissociative reaction where the adsorbate is deprotonated to form surface hydroxyl groups. These hydroxyls are known to be present on highly ionically bonded surfaces such as oxides. 40 The presence of these hydroxyl groups has also been shown to change the surface free energy of the material, effectively reducing its acid and base components and increasing the observed contact angle for polar liquids such as water. 38 Indeed, Gentleman and Ruud observed an increase in water contact angle from ∼0° to ∼35° as hydroxyls were added to a single crystal alumina surface.
Crystallographic structure in surface free energy
With the exception of platinum group metals, which are rarely used outside of dental applications, metals used as biomaterial implants develop an oxide scale on their surfaces that dominates the surface properties of the implanted material. For high-energy materials, there are a limited number of ways that the surface free energy can be modified. The most obvious is through the modification of the number of Lewis acid and base sites that are available as well as their strength. This can be accomplished by changing the crystallographic plane that is exposed at the surface.
Bulk alpha-alumina, for example, has the corundum crystal structure where aluminium atoms sit in two-thirds of the octahedral positions formed by the close-packed oxygen planes and the other third are left vacant. These vacancies are arranged on the (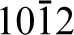
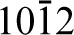
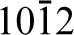
The (0001) surface, unlike the (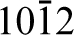
This effect is also relevant in other biomaterials. The rutile form of titanium dioxide has also been studied (TiO2). This is the stable native oxide present in most titanium-containing implant materials. The (001) plane of this oxide, which can be purchased in single crystal form, sees the cation coordination decrease from six in the bulk to four at the (001) surface. 40 Owing to the low coordination at the surface, TiO2 tends to be prone to reconstruction, and (011) and (104) facets are often observed after even moderate thermal exposures. This may lead to a decrease in surface free energy and, in very extreme cases, increases the surface roughness of the material.
These results teach several things. First, water contact angle cannot discriminate between all the components of the surface free energy that will likely control biological interactions at a surface. Water, for example, has significant non-polar, acid and base components of surface free energy. Therefore, many combinations of these three components can result in the same contact angle measurements. Second, if there is a strong acid–base interaction between proteins and surfaces, a better understanding of how those interactions take place and the limits of those interactions must be explored.
Other methods for modifying surface free energy
In addition to changing the crystallographic termination of the surface, surface free energy can also be modified through the non-equilibrium addition of ions. This can be accomplished in several ways, all of which bombard the surface with high energy radiation of some sort followed by rapid quenching to trap in the metastable state. One example of this is the use of CO2 laser treatment. As the equilibrium concentration of oxygen in many oxide scales is dependent on temperature, laser surface treatments can modify oxygen concentration at the surface very effectively. 44 Likewise, plasma treatments, which can be used to sterilise surfaces, also modify surface energy, often because they remove weakly bound organics from the surface. This removal of organic surface contamination is important because most hydrocarbon ‘dirt’ on surfaces has very low surface free energy with predominantly non-polar nature. Since the adhering or wetting liquid only ‘sees’ the top monolayers of the surface, cleaning off these non-polar contaminants of the surfaces can dramatically alter surface free energy and water contact angle. Additionally, extra surface chemistry can be added to the surface using the plasma. Oxygen plasmas, for example, can oxidise or hydroxylate the surface, increasing or decreasing the surface free energy. 38 Carbon tetrafluoride (CF4) plasma treatments, on the other hand, often result in very low surface energy fluorite-containing groups covering the surface.
The biological response to surface free energy
As we have seen, surface free energy, which differs from wettability, is a fundamental property of a material associated with chemical bonds at its surface. These bonds interact with species that it comes in contact with. In the biological environment of the body, such species include water, ions, sugars, proteins and cells. How does the surface free energy of a material affect these interactions and thus the overall biological response?
Protein adsorption
When a biomaterial surface first comes in contact with a biological fluid, the oriented adsorption of molecules creates a conditioned surface which will later govern cell–surface interactions. In short, a cell never encounters a completely clean surface. Materials are instead covered by the components of the fluid in which it is immersed, which includes water, ions, sugars and proteins. In vivo this fluid can be blood, saliva or interstitial fluid, and in vitro, it is often cell culture medium and serum constituents. Human plasma may contain as many as 1175 distinct proteins, any of which may interact with a surface. 45 For cells, which will move more slowly than proteins, it is this conditioned surface that they first encounter and it governs how the cells attach, their resulting morphology and behaviour.
Proteins are often regarded as the primary and most important constituent of biological fluids that condition a surface (although others argue that water itself plays a significant role 46 ). Proteins are ubiquitous biological molecules responsible for everything from forming tissues to mediating most biological reactions, and from a materials point of view, are highly surface active. Indeed, it has been estimated that the surface-associated concentration of protein is some 1000 times that in solution. 47 Once adsorbed, the charged domains on proteins' surfaces make it such that even significant dilution of the protein solution will fail to remove them from the surface. Which proteins adsorb to a surface from a biological fluid, their resulting bioactivity and their time-dependent concentrations on the surface, is the subject of a great deal of research and remains controversial. Although beyond the scope of this review, the reader is referred to excellent reviews 48,49 and a thorough opinion piece by Vogler, 50 which nicely describes many fundamental issues surrounding the interactions at the protein–surface interface.
Despite such controversies, however, a few general observations are widely accepted and will be discussed here. When a biomaterial comes in contact with a biological fluid, the constituents of the adsorbed protein layer will be governed by a combination of the material's characteristics, the concentration of the proteins in solution and their affinity for the surface. Proteins in high abundance tend to adsorb quickly, but are often later replaced by proteins with higher affinities for the surface, but often lower concentrations in the solution, in a phenomenon termed the ‘Vroman Effect’. Over time, the adsorbed protein layer therefore changes from one dominated by proteins that are abundant to one that contains proteins with high affinities and/or great resistance to displacement. For example, on various polymers, vitronectin adsorption is highly enriched compared to its concentration in the serum. 51 These observations suggest that vitronectin overcompetes proteins that are more abundant in the solution for binding sites on the material surface and/or exhibits greater resistance to displacement once adsorbed.
Many general trends for protein associations with biomaterials are described in the literature terms of materials' ‘hydrophobicity’ and ‘hydrophilicity’. As discussed above, descriptions of surface wettability lack the precision to define true surface reactions because the relative ‘phobicities’ of several surfaces can be identical while their surface chemistries can remain quite different. For this reason, descriptions of hydrophobic/hydrophilic surface interactions should always be considered in this context. Indeed, much controversy in the literature regarding protein–surface interactions may be explained by the inappropriate attribution of these characteristics to particular surfaces. Nevertheless, as much of the literature relies on this terminology, reported trends are mentioned here and are described using these terms.
One general observation is that protein adsorption is greater on hydrophobic surfaces compared to hydrophilic. Larger adhesion forces for a number of blood proteins have been consistently observed on more hydrophobic surfaces compared to hydrophilic surfaces, and these adhesive forces increase with surface–protein contact time. Hydrophobic surfaces seem to irreversibly bind albumin, an ubiquitous and abundant protein in serum. Alternatively, hydrophilic surfaces bind proteins such as fibronectin, a common component of the extracellular matrix (ECM), which promotes cell adhesion. Counter intuitively, the inverse is often observed with cell attachment: cells generally attach better on hydrophilic surfaces as compared to hydrophobic. However, which proteins adsorb to a surface also appears to be essential in determining the resulting cell behaviour. For example, in studies in which proteins were selectively removed from serum, it was found that vitronectin, but not fibronectin, was essential for early cell adhesion. 52 Many studies have also demonstrated that protein adsorption to metallic and ceramic surfaces mediate bone cell attachment and spreading. Indeed, fibronectin is often reported to be essential for osteoblast attachment and differentiation. Its enhanced adsorption (along with that of vitronectin) on hydroxyapatite as compared to stainless steel and titanium is thought to be responsible for significant increases observed in osteoblast attachment. 53
Proteins, however, also undergo structural rearrangements when they encounter a surface as a result of charge interactions. In this way, not only is the concentration of protein and its charge important in its interaction with the surface, but also its potential for conformational change once adsorbed to the surface. Protein activity resulting from conformational changes upon adsorption appears to play an important role in subsequent cell attachment. 54,55 For example, while it appears that adsorbed fibronectin maintains its functionality on hydrophilic surfaces, it displays markedly reduced functionality on hydrophobic surfaces. 56 In contrast, vitronectin's activity does not appear to vary with surface wettability. 57 In short, during these initial interactions, surface free energy controls which species initially adsorb, their orientation, conformation and thus bioactivity.
More recent studies utilising self-assembled monolayers (SAMs) which control the presentation of functional groups (see ‘Self-assembled monolayers’ section) have also reported that the key adsorbed protein that controls cell attachment is vitronectin. 58 Studies on these surfaces demonstrate that wettability alone does not play the dominant role in determining subsequent cell behaviour. Hydrophilic surfaces with OH and PEG functional groups which failed to adsorb vitronectin did not promote cell attachment. This observation suggests that the characteristics of the surface that allow for protein absorption, rather than the wettability of the surface itself, play the most important role in subsequent cell attachment and behaviour. 58
Integrin-mediated cell–surface interactions
Cells are not perfect spheres that simply adhere to a surface. Instead, the cell membrane is covered in a pericellular matrix, a mostly hyaluronan-based coating that extends out from the surface of the cell, 59 and thus plays a role in mediating cell–surface interactions. Cells do not adhere uniformly to a surface either. Although non-specific adhesion mediated by ionic and van der Waal's forces between the negatively charged cell membrane and a surface likely acts as a complementary mechanism, cells are thought to attach via discrete points, often referred to as focal adhesions, created by protein clusters and mediated by integrins (Fig. 4). Integrins are a major family of cell surface receptors that mediate cell adhesion to ECM proteins (Fig. 5). The integrin family is composed of 22 transmembrane heterodimers consisting of two types of sub-units: α and β. Osteoblasts have been shown to be able to express a variety of integrin subunits including α1, α2, α3, α4, α5, α6, and αV, and β1 and β3. 60,61 Different types of cells express a particular complement of integrins and different integrins recognise different ECM proteins, with some being highly specific and others able to bind a variety of sequences.
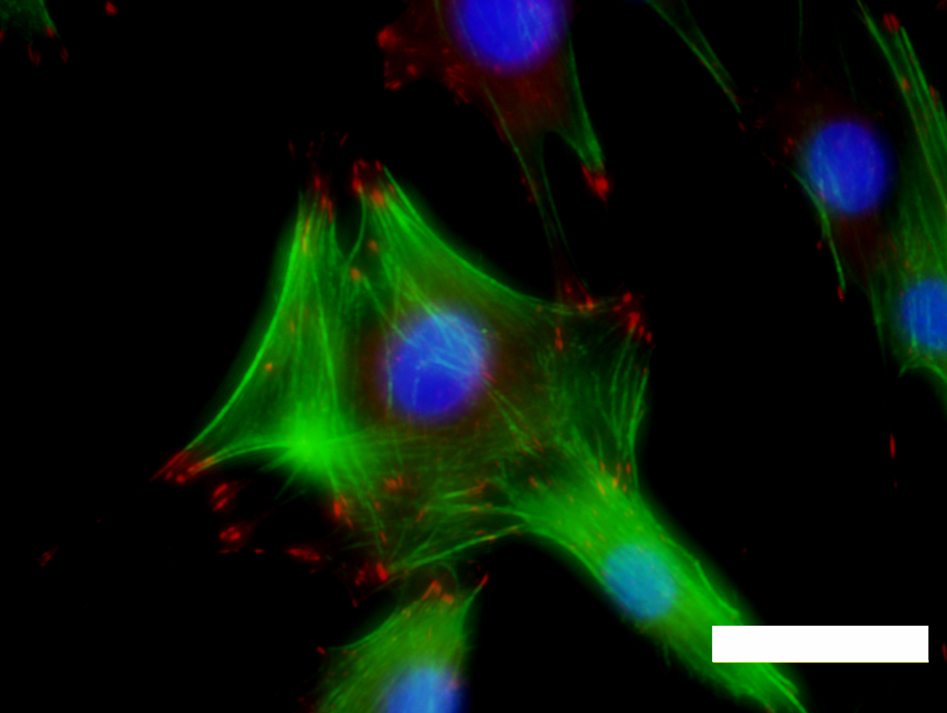
Fluorescence micrograph of MC3T3-E1 mouse preosteoblast cells cultured on a titanium surface. The actin cytoskeleton appears green. Focal adhesions appear red and were detected by staining for vinculin. Cell nuclei appear blue. Scale bar = 50 μm
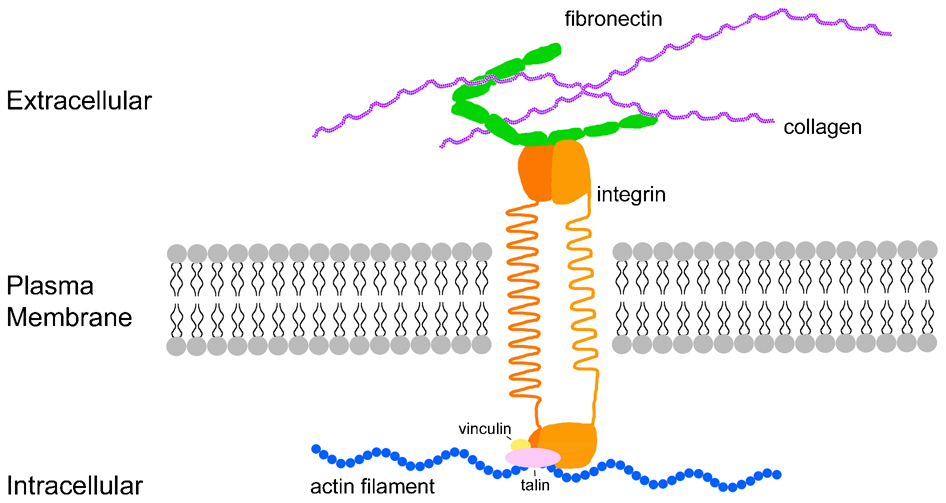
Diagram showing an integrin receptor in a mammalian cell. Integrins are transmembrane proteins that mediate linkages between ECM proteins such as fibronectin and the intracellular cytoskeleton, depicted here as an actin filament (blue). The focal adhesion complex on the intracellular side is composed of a number of proteins, including vinculin and talin, which are highlighted here in yellow and pink, respectively
Although conflicting data are rife in the literature, α5β1 integrin and αVβ3 integrin, among others, appear to play important roles in osteoblast adhesion and differentiation, 62,63 and so the adsorption of proteins that are recognised by these integrin sequences may be important in mediating their adhesion. The β1 integrin (as part of the α5β1 integrin subunit), in particular, appears to play an important role in osteoblastic differentiation of precursor cells. 63–65 Moursi et al. demonstrated that blocking α5β1 integrin inhibited osteoblast differentiation (as determined by gene expression analyses) and mineralised nodule formation when MG63 (human osteosarcoma) cells were cultured on gelatin-coated tissue culture plastic. 63 However, this effect may not be specific as they reported that other integrins were regularly expressed by osteoblasts both in vitro and in vivo. Ligand binding by the β1 subunit may also be necessary for matrix mineralisation 64,66 as blocking its function with an antibody inhibited osteoblasts' ability to form mineralised nodules. Since α5β1 is the only integrin that binds fibronectin exclusively, the presence of this protein adsorbed to a biomaterial surface may be necessary to allow for osteoblastic differentiation.
However, which particular integrins are expressed when a cell comes in contact with a surface may also be material specific. Although other factors which affect adhesion such as topography confound general observations, on cobalt–chromium alloys for example, osteoblasts do not always express α3, α6 and β3 subunits. 60 Similarly, integrin expression varied when cells were cultured on grit-blasted versus calcium phosphate-coated titanium, 67 or indeed on a variety of standard biomaterial surfaces: α5 and α6 were not expressed when cells were cultured on titanium or cobalt–chromium alloys, and α3 was not expressed when they were cultured on cobalt–chromium alloys or if the titanium surface was roughened. 60 Olivares-Navarette et al. similarly examined the role of integrins in osteoblast behaviour on titanium surfaces; 68 they demonstrated that while α5β1 mediated cell attachment and proliferation, it inhibited differentiation. Instead, α2β1 was required for osteoblastic differentiation.
Taken together, these results suggest that osteoblast adhesion to different substrates is mediated by differential expression of integrins. These differences are likely due to the differential adsorption of various proteins on the different materials, which is again related to their surface characteristics. On hydrophobic surfaces, human foetal osteoblasts express significantly lower levels of the α5 and β3 integrin subunits compared to cells cultured on hydrophilic surfaces. 69 Surprisingly, however, findings of enhanced integrin expression and increased cell spreading do not appear to correlate with osteoblast-specific gene expression, which is necessary for appropriate differentiation and bone tissue formation. For example, the expression of osteopontin, a protein in bone and often used as a marker for osteoblastic differentiation, is higher in human foetal osteoblasts cultured on relatively hydrophobic silane-treated quartz compared to more hydrophilic surfaces. 69 Similarly, Lim et al. report that although osteoblast adhesion and spreading are inversely correlated with contact angle on materials with a range of hydrophilicities, they could not detect similar trends when measuring alkaline phosphatase activity, an enzyme necessary for mineralisation, in human foetal osteoblast cells. 70 Indeed, while appropriate protein adsorption to a biomaterial surface is important for subsequent cell adhesion, more cell adhesion is not necessarily better and promoting particular protein adsorption to encourage specific integrin interactions may be the more promising means to promote osteoblast differentiation and bone tissue formation on a surface. Just as biomaterial surfaces used to induce bone formation can be described as either class A (osteogenic/osteoproductive, promotes bone formation) or class B (osteoconductive, allows bone to migrate across the surface), surfaces that allow not only for cell attachment, but also encourage appropriate differentiation and tissue formation will better promote osseointegration.
Cell adhesion and differentiation
Most cells, including osteoblasts, are anchorage dependent and will not survive in suspension. That is, the cells must be adherent to a surface to remain viable. It has long been recognised that a cells' ability to adhere onto a surface plays a significant role in osteoblast differentiation. And indeed, morphology is of critical importance in maintaining or inducing a particular cell to maintain its phenotype, or tissue-specific identity. For example, when mesenchymal stem cells (MSC), which have the ability to differentiate into a variety of musculoskeletal cell types including osteoblasts, are cultured on large adhesive islands that permit cell spreading, they tend to adopt an osteogenic phenotype, while those whose spreading is limited become fat cells called adipocytes. 71
In general, adherent cells that spread on a surface will proliferate, while those that do not and remain rounded will divide at a much lower rate. 72 Cell shape is strongly correlated with surface properties, and generally increases in size with increases in hydrophilicity. 69 On hydrophilic surfaces, cells also show strong focal adhesions formation and stress fibre bundles within 3 h of plating. Conversely, on hydrophobic surfaces, staining for actin, an important cytoskeletal protein, is far more diffuse and vinculin staining for focal adhesions is lacking. 73 However, while cell attachment to a biomaterial surface is clearly important for good implant integration, the trend for ‘improved’ cell behaviour with increasing adhesion is not perfect. Indeed, excessive adhesion may actually be detrimental. One report of high levels of MSC attachment on positively charged surfaces concomitantly showed reduced cell spreading and differentiation. 74
Despite trends in cell behaviour on hydrophobic versus hydrophilic surfaces, morphological differences between cells on various surfaces tend to disappear after 48 h. This observation is thought to be attributable to cells' ability to compatibilise the surface with secreted proteins. On surfaces that do not support cell attachment (likely because of the lack of appropriate adsorbed proteins) cells secrete adhesive proteins such that they can then adhere and spread. Such observations are confirmed by studies using serum and serum-free medium, the former of which is rich in proteins which readily adsorb to many surfaces. Healy et al. showed that materials that did not allow for cell attachment in the absence of serum, but were adhesive to primary osteoblasts in the presence of serum proteins, could be made adhesive if left in the presence of cells for approximately 24 h. 75 The authors concluded that endogenous protein secretion rendered the surfaces adhesive.
However, cells do not just secrete proteins to compatibilise surfaces, they also appear to reorganise those that are already adsorbed. When cultured on hydrophilic clean glass and hydrophobic octadecyl glass, human fibroblasts display typical behaviour, they poorly adhere to the hydrophobic material but display better attachment to the hydrophilic. 76 Attachment can be rescued by pre-coating the surfaces with fibronectin, however, cells still fail to proliferate on the hydrophobic surfaces. Nevertheless, when the authors conjugated the fibronectin to a fluorophore they found that on the hydrophilic surfaces, cells were re-organising the fibronectin, and this phenomenon seemed to contribute to their ability to proliferate. Alternatively, cells were unable to reorganise the fluorescent fibronectin on the hydrophobic surfaces.
Surface free energy and cell behaviour
In general terms, researchers have often reported that high surface free energies or wettability promote cell adhesion, while surfaces with low surface free energies are not supportive of cell attachment and spreading. 49,70,73,77–79 However, because wettability and surface free energy are not necessarily directly correlated and because the surface interactions are complex, this is not strictly reported as true across the board. A number of researchers have shown that instead of there being a direct correlation between contact angle and cell attachment, there rather seems to be an ideal contact angle that best directs cell proliferation and behaviour and this occurs around 60°–70°. 78–80 As such, 65°contact angle is often reported as a ‘magic’ number for biomaterials to achieve ideal levels of cell attachment and spreading.
However, there is clear data from many authors indicating that this does not necessarily hold true either and contact angle is not a good predictor of cell attachment and behaviour. Howlett et al. report that contact angle measurements on titanium, alumina, stainless steel and polyethyleneterephthalate (PET) between 37° and 83° failed to show a predictive relationship with cell attachment. 52 Groth and Altankov have similarly shown that while fibroblasts spread more and create more stress fibres and stronger focal adhesions on hydrophilic surfaces compared to hydrophobic, proliferation did not follow the same trend and was higher on the hydrophobic materials (octadecylsilane and silicone) compared to hydrophilic (glass and aminopropylsilane). 78 Human foetal osteoblasts also appear to show a strong preference for hydrophilic quartz (SiO2) over similarly hydrophilic plasma-treated glass (SiOx). 70 The reason for the difference in cell behaviour due to surface chemistry, quite independent of surface wettability, is unclear, but again likely relates to protein adsorption and reiterates the fact that contact angle alone should not be used as a predictor of cell–surface interactions.
Moreover, a number of studies have shown that the general trend for increasing cell attachment and spreading on hydrophilic surfaces over hydrophobic does not always hold true either. Indeed, Padial-Molina et al. studied the effects on MG63 cells by adding methyl groups to oxidised silicon surfaces, a system that allowed them to change the surface energy without concomitant changes in roughness. Increasing water contact angle (hydrophobicity), which correlated with increased methylation, lead to increases in cell attachment and spreading. 81 Similarly, Kennedy et al. examined osteoblast attachment, spreading and proliferation on SAM surfaces (see ‘Self-assembled monolayers’ section) whose hydrophilicity had been altered by UV oxidation. 82 They report linear increases in cell proliferation with increases in hydrophobicity and the lowest cell spread area on the most hydrophilic surfaces. How can such results be explained? In addition to the previously discussed issues with defining such materials by their wettability, other factors may also play a role. One reason may be that many surfaces that are used for cell culture or to assess the effects of substrate chemistry or wettability on cell behaviour are heterogeneous at the molecular level. Indeed, plastic and glass tissue culture surfaces, for example, lack uniform surface organisation. There is a significant need, therefore, to assess the effects of these variables on surfaces that are precisely defined.
Some have carried out more ‘defined’ experiments by examining cell behaviour on crystals, which are highly ordered and have a repetitive, defined chemical nature at the atomic scale (see ‘Crystallographic structure in surface free energy’ section). Hanein et al., for example, examined A6 Xenopus epithelial cell behaviour on calcite and calcium (R,R)-tartrate tetrahydrate crystals, on which the orientation of the carboxylate and hydroxyl groups of the tartaric acid will differ resulting in differences in surface free energy. 83 When cells were grown on the two surfaces, the (011) face was found to be highly adhesive, while the (101) face fostered significantly slower cell attachment. The authors attributed these effects to differences in the surface distribution of lattice water molecules and charges. It is interesting to note that fibronectin and other serum protein adsorption to the different crystal surfaces was comparable, suggesting that the conformation or orientation of the adsorbed proteins, rather than their quantity, may have played a role in the resulting cell behaviour. 84
Similarly, Faghihi et al. examined the effect of titania crystal orientation on MC3T3-E1 mouse osteoblast and rat fibroblast cell adhesion.
85
They prepared (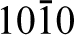
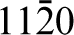
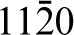
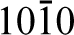
Similar studies have also been carried out on hydroxyapatite, the primary mineral constituent of bone. Hydroxyapatite (Ca10(PO4)6(OH)2) has the space group P63/m and generally facets to display two sets of growth planes: the a,b-type and the c-type. The a,b-type is defined as (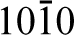
Surface free energy on polymer, metal, ceramic and self-assembled monolayer surfaces
As noted in ‘High and low surface energy surfaces’ section different materials are dominated by different types of bonding. As we have seen, the ‘high energy’ and ‘low energy’ surfaces resulting from different types of bonds will affect protein adsorption and subsequent cell behaviour. Here, the authors review some of the general trends that are observed in cell behaviour on particular materials as a result of these interactions.
Polymers
Polymers encompass a wide-ranging group of large molecules with repeating units and are used in a variety of biomaterials applications. For example, polyethylene forms one of the bearing surfaces of many orthopaedic implants and polymers such as PTFE and PET are regularly used in large blood vessel repairs. Polymers are often considered to have low-energy surfaces because their covalent and van der Waals bonding often lend the surfaces non-polar, and thus a hydrophobic nature. However, as polymers are a diverse class of molecules, this is not always true. As discussed above, cells generally adhere poorly to hydrophobic materials, and so for many biomaterial applications in which cell adhesion is required, polymer surface modification is often necessary.
Plasma treatment, which was discussed in ‘Other methods for modifying surface free energy’ section, has often been used to increase the wettability and enhance cell adhesion on polymers. 87,88 Poulsson et al., for example, examined the behaviour of human osteoblasts on ultra high molecular weight polyethylene that had been modified by UV/ozone treatment to incorporate surface oxygen, and thus increase hydrophilicity. 89 Cells showed general trends for increased attachment, proliferation and mineralisation on the more hydrophilic surfaces. Similarly, Wei et al. examined plasma-treated hexamethyldisiloxane surfaces. They showed that fibronectin preferentially bound to hydrophilic surfaces while albumin to hydrophobic, and that hydrophilic surfaces strongly supported osteoblast attachment. 90
However, it is clear that plasma treatment often does not simply alter surface chemistry; it can also affect topography and so plasma treatment should not be strictly considered as a means to alter surface chemistry alone. Yang et al. showed that plasma treatment of polydimethylsiloxane actually smoothed the surface with short treatments and roughened it with longer treatments due to the formation of surface cracks resulting from tensile stresses. 88 Similarly, Xu and Siedlecki examined protein adhesion on low density polyethylene that had been plasma treated. 91 Plasma treatment decreased water contact angle, however, it also simultaneously affected surface roughness as determined by atomic force microscopy. In short, ‘low energy’ polymers can often be made more cell adhesive, but care should be taken in interpreting such results.
Metals
Metals and metal alloys including titanium, titanium alloys (Ti–6Al–4V), and cobalt chromium alloys (Co–Cr–Mo) are frequently used in orthopaedic and dental applications because of their favourable mechanical properties, chemical stability and biocompatibility. Probably because of their ‘high energy’, the surfaces of many metals and metal alloys are suitable for cell attachment. However, as noted above, metal and metal alloys exposed to air develop a surface oxide scale. Particularly in the case of titanium and its alloys, biocompatibility has been attributed to their ability to form this ‘passivating’ oxide layer (titania TiO2). This fundamental property should be taken into consideration when examining behaviour on these surfaces, as proteins and cells will often interact with the oxide rather than titanium metal.
A number of authors have explored the effects of varying the surface free energy of metals and metal alloys on osteoblast behaviour. In many cases, however, surface treatments affected both surface chemistry and topography, making the contribution of each difficult to discern. Feng et al., for example, examined the behaviour of rabbit osteoblasts on titanium surfaces that had been heat treated in various oxidation atmospheres to alter the surface hydroxyl group presentation. 92 Here, higher levels of oxidation correlated with increased roughness, increased surface energy and more hydroxyl groups. These parameters fostered greater numbers of adhered cells and higher alkaline phosphatase activity. Similarly, MacDonald et al. showed that the adsorption of fibronectin and MG63 human osteoblast attachment onto titanium alloys modified by heat and peroxide treatment was highly correlated with changes in surface chemistry. 93 That is, treatments, which increased the content of Al or V at the surface, enhanced fibronectin-promoted cell adhesion. They attributed this result to increased bioactivity of the adsorbed fibronectin, however, AFM measurements demonstrated that treatments significantly affected surface topography, which could have similarly affected cell behaviour. Others have used a technique called micro-arc oxidation to alter the surface chemistry of titanium implants. 94 While this technique added oxides to the surface layer, it simultaneously created a porous surface with enhanced roughness, so again the contribution of changes in oxide presentation is difficult to determine.
Tsukimura et al. attempted to separate the effects of surface topography from surface chemistry by creating titania and titanium surfaces with the same roughness (as determined by atomic force microscopy). 95 They report enhanced rat marrow stromal cell proliferation on the titania surfaces, however, expression levels of osteogenic genes were not significantly different. Interestingly, they also found that mineralised nodule area and alkaline phosphatase staining were enhanced in the titania samples compared to the titanium. They attribute these results to the greater wettability of the titania samples and the enhanced oxygen content in their surfaces. Nevertheless, the true nature of these surfaces was uncertain and overall there are very few studies that effectively separate the effects of surface free energy from topography on cell behaviour, so it is quite difficult to demonstrate clear trends.
Ceramics
Bone is a nanocomposite material composed of organic, predominantly collagen fibrils, interspersed with platelike carbonate substituted hydroxyapatite crystals. At particular ectopic (non-bone) sites such as muscle, some groups have noted that the surgical placement of specific forms of calcium phosphate can stimulate bone formation. 96,97 To encourage bone formation on biomaterial surfaces, many have similarly hypothesised that placing cells in contact with a hydroxyapatite-like material would be preferable to other foreign surfaces such as metals. As such, numerous metallic implants for joint arthroplasty procedures are regularly coated with calcium phosphates, and some orthopaedic implants, particularly parts of hip replacements, are made from ceramics. Like metals, the ‘high energy’ surfaces of many ceramics tend to be conducive for cell attachment, however, as it is difficult to fundamentally alter their surface free energy, studies showing clear effects of changes in surface free energy of ceramics on cell behaviour are not widely available.
In one study, Redey et al. examined the adhesion and proliferation of human osteoblasts on stoichiometric and type A carbonate apatite, which is similar to bone mineral. 98 They report that cell attachment was significantly lower and cells spread far less on the carbonate apatite when compared to standard hydroxyapatite. After 6 weeks in culture, collagen synthesis was also significantly lower on the carbonate apatite when compared to the pure hydroxyapatite, but osteocalcin, a bone-specific protein, production was unaffected. In another study, surface charge was examined for its role in cell behaviour on ceramics. Tarafder et al. electrically polarised β-tricalcium phosphate/hydroxyapatite composites, generating surfaces with positive and negative charge. 99 Negatively charged surfaces promoted enhanced osteoblast adhesion, proliferation and ECM formation, regardless of the composite composition when compared to positively charged or uncharged surfaces. However, again it is difficult to discern the true contribution of surface free energy to cell behaviour on these surfaces because surface free energy is often not the only surface property that has been altered.
Self-assembled monolayers
As we have seen, it can be quite difficult to uncouple the effects of surface free energy from other effects to determine its role in protein adsorption and cell interactions. Many biomaterial surfaces, which have been explored as substrates for cell attachment and clinical use, also often possess a large degree of surface heterogeneity. That is, the type, distribution and presentation of functional groups can vary from one region to the next. Moreover, many surface treatments intended to alter the presentation of reactive groups simultaneously alter topography, making the differences attributable to each difficult to discern. These differences may account for much of the controversy and lack of agreement among different studies. Therefore, over the past two decades, researchers have turned to SAM, which have flat, chemically well-defined surfaces created by controlling the presentation of functional groups. 100 These have been used to lend insight into the role of surface chemistry and surface free energy in protein adsorption, cell attachment, osteoblast differentiation and mineralised matrix formation.
In general, many of the typical patterns for protein adsorption and cell behaviour observed on other less defined materials are similarly observed on SAM systems. For example, on SAM of organosilanes, osteoblast proliferation has been observed on oxidised surfaces and NH2 surfaces, both of which are hydrophilic. 101 However, such patterns do not always hold true. CH3 and CF3 groups, both of which are hydrophobic, have been shown to enhance and inhibit cell proliferation, respectively. 101 Lopez et al. similarly showed that in the absence of serum, cells were able to attach to both hydrophobic and hydrophilic functional groups on SAM of alkanethiols on gold. 102 Using similar SAM systems, Faucheux et al. also demonstrated COOH- and NH2-functionalised surfaces promoted integrin-dependent cell spreading, while CH3, OH and PEG inhibited it. 58 Here, PEG- and OH-terminated surfaces were highly wettable, but did not promote cell attachment. Moreover, alkyl chains terminated with SiO2 or Br groups have been shown to support 3T3 cell attachment, but hydrophobic surfaces terminating in CH3 or C = C groups gave an intermediate response, while COOH, CN and Diol yielded the poorest results, 103 suggesting a lack of clear trend with wettability.
To try to explain some of these discrepancies, some studies have examined simple systems with a single protein such as fibronectin. Lee et al., for example, examined the role of the α5β1 integrin when fibronectin was adsorbed to an alkylsilane SAM, which presented CH3 (hydrophobic), NH2 (positively charged), COOH (negatively charged) and OH (neutral hydrophilic) groups. 104 Their analysis showed that less fibronectin adsorbed to hydrophilic groups compared to hydrophobic (CH3 and NH2). This trend confirms previous experiments on standard materials, which have similarly shown that protein adsorption is enhanced on hydrophobic surfaces compared to hydrophilic. Also, as had been predicted for hydrophobic surfaces, a model cell line that only expressed the α5β1 integrin bound more efficiently to fibronectin adsorbed to the hydrophilic and negatively charged groups (COOH and OH) as compared to the hydrophobic and positively charged groups. The authors attributed this to the disruption to the native structure of fibronectin on hydrophobic surfaces and the lack of specific integrin binding to these groups.
Others have tried to utilise SAM to explore the role of functional groups in cell behaviour, specifically osteoblast differentiation. Keselowsky et al. cultured MC3T3-E1 on SAM of alkanethiols on gold, again with CH3, NH2, COOH and OH groups. 105 They found that OH and NH2 surfaces upregulated osteoblast-specific gene expression, alkaline phosphatase activity and matrix mineralisation compared to that measured in cells grown on COOH and CH3 substrates. They attributed these differences to the binding of specific integrin receptors to adsorbed fibronectin. In another study, the same authors also reported that binding to the various chemistries was highly integrin dependent. 106 Interestingly, the authors also found that matrix mineralisation of MC3T3-E1 was surface chemistry dependent, with OH and NH2 chemistries promoting high levels of mineralisation while COOH and CH3 did not. The authors speculate that the most important factor driving these differences in cell behaviour is how the various chemistries on the surfaces affect the functional presentation of adsorbed fibronectin.
Conclusions
The cell response to a biomaterial surface is complex, but will be governed by the interactions of water, proteins and cells with the chemical and topographical nature of the surface. Here the authors defined surface free energy, as this fundamental property of the material surface will govern the first interactions with the biological environment. The authors have also explained how surface free energy differs from wettability and outlined how simple measurements of hydrophobicity versus hydrophilicity lack precision when attempting to explain cell–biomaterial interactions, particularly when topographical changes are also introduced. As many biomaterial surfaces have been defined in terms of wettability, in many cases the true contribution of surface free energy to protein adsorption and cell adhesion remains uncertain. Nevertheless, studies have shown that protein adsorption and its resulting conformational changes upon adsorption appear to play fundamental roles in dictating cell behaviour on biomaterial surfaces. More studies to understand these interactions are desperately needed, particularly to discern the role of material surface properties in controlling osteoblast attachment, differentiation and mineralised matrix formation. Indeed, to effectively design materials for successful clinical applications there remains a need for systematic experiments that decouple the effects of changes in surface free energy from topography to gain a clear picture of cell response. Experiments that control these factors, such as those with SAM, are beginning to elucidate the effects of chemical groups alone on cell attachment and differentiation. Similarly, more experiments that examine cell behaviour on single crystal materials that are chemically defined and atomically smooth should provide us with a better understanding of how the atomic arrangements of various crystal planes affect such interactions and provide fundamental insight into these complicated phenomena. The emerging field of nanotechnology, which promises more precise engineering of surfaces and their characterisation, may also allow for experiments that truly decouple the effects of topography from changes in surface free energy. Insights gained from these efforts have the potential to provide us with a set of rules that truly describe how surface properties of a material will affect its interactions with the biological milieu. Once achieved, rational design of biomaterials to elicit particular cell responses will no longer just be wishful thinking but reality, and the authors hope that researchers will continue to focus their efforts to achieve this.
Biographical sketches
Molly M. Gentleman is an Assistant Professor of the Department of Materials Science and Engineering at the State University of New York at Stony Brook. She received her PhD in Materials from the University of California at Santa Barbara where she developed optical characterisation techniques for ceramic coatings in harsh environments. Before joining SUNY Stonybrook, Dr M. Gentleman held positions as an Assistant Professor in the Mechanical Engineering Department at Texas A&M University, and as a staff Materials Scientist at GE Global Research Center. She currently holds inventor rights on 15 issued patents in the design and control of the wettability of ceramic surfaces for a range of applications, and is a lead inventor on 16 further pending applications. Dr M. Gentleman's current work at SUNY Stony Brook focuses on the modification of ceramic materials by conventional techniques followed by non-traditional characterisation of their surfaces and structures using Raman spectroscopy, surface topography, and surface free energy measurements. This work has lead to a broad range of new insight into these materials and hence new applications including turbine coatings, superhydrophobic surfaces, and biological coatings. Her work has been published in journals including Langmuir, Acta Materialia, European Cells and Materials, and the Journal of the American Ceramic Society. Dr M. Gentleman has received funding awards from the Defense Advanced Research Projects Agency (DARPA) and NASA and her work has been featured in CERAMIC TECH TODAY and Materials Views.
Eileen Gentleman is a Wellcome Trust Research Career Development Fellow in the Department of Craniofacial Development & Stem Cell Biology at King's College London. She joined Imperial College London in 2005 as a post-doctoral research associate after completing her PhD in Biomedical Engineering at Tulane University (USA), where she investigated collagen-based biomaterials for engineering ligament tissue. In 2011, she moved to King's where her research focuses on utilising biomaterial systems to direct stem cell differentiation to bone and cartilage and material characterisations of engineered tissues. She is particularly interested in the osteochondral interface, the transitional tissue that connects cartilage to bone, and the role it plays in normal joint function. Her multi-disciplinary research interests also include biomineralisation and the role of mechano-sensing in tissue development. She has also worked extensively with biomaterials, including bioactive glasses, and is interested in the biological effects of surface energy and ion release on stem cell differentiation. In 2009, her work on the characterisation of engineered bone formed from different stem cell sources was published in Nature Materials. Her work on biomaterial systems has also been published in journals such as Biomaterials, Acta Biomaterialia, Tissue Engineering Part A, and European Cells and Materials. She has also published on the fundamental mechanisms of biomineralisation in bone and cardiovascular tissues in prestigious journals such as Proceedings of the National Academy of Sciences USA and Nature Materials. Dr E. Gentleman has received funding awards from the Wellcome Trust, the Rosetrees Trust, the Royal Society and Orthopaedic Research UK. The Orthopaedic Research Society named her as a finalist for their New Investigator Recognition Award (2010) and in 2013 her work in tissue engineering and regenerative medicine was recognised with a prestigious Philip Leverhulme Prize.
Footnotes
Acknowledgement
Eileen Gentleman gratefully acknowledges a Research Career Development Fellowship from the Wellcome Trust and continued support from the Rosetrees Trust.