
Abstract
The effect of pure Zn2+ and combined Zn2+ and Mn2+ supplements on the growth and ligninolytic activity of wood-rotting basidiomycetes was studied. Based on measurements of minimum inhibitory concentration and minimum decolourisation-inhibiting concentration available from cultures of Inocutis sp., Trametes versicolor and Gloeophyllum trabeum, and a comparison of such results with those obtained in previous work for P. chrysosporium and P. atropurpurascens, metal supplements were shown to inhibit fungal physiology at concentrations within the study range. Average minimum inhibitory concentration values were consistently lower for ZnSO4 than for ZnSO4/MnSO4 (2 mM Zn2+), demonstrating the antagonistic effect of the tested cations. Furthermore, because the ligninolytic system of P. atropurpurascens had shown a high tolerance to metals, its ligninolytic and germinative behaviour was compared with that of P. chrysosporium. Conidial germination was inhibited at 1 mM of Zn2+ in the two strains, whereas extracellular activity was inhibited at concentrations above 2 mM.
Introduction
Copper has traditionally been used as an active ingredient in wood preservative formulations (Jellison et al. 1997; Bagley and Richter 2002; Pohleven et al. 2002), and some of these can also contain chromium and arsenic (i.e. CCA). Copper, arsenic and chromium are broadly toxic heavy metals that have come under increasing environmental pressure because of their potential effect on non-target organisms. Identifying alternative systems will become critical for retaining our ability to use wood in adverse environments (Humar et al. 2006).
A hydro-metallurgical process for battery recycling that generates no effluents or emissions (Rabinovich and Ibáñez 2004) produces an acidic solution of ZnSO4 and MnSO4 (at a Zn2+/Mn2+ molar ratio of 2∶1), also containing trace levels of other heavy metals, as the main process product of the recycling process. The antifungal activity of this system against white-rot fungi (WRF) and brown-rot fungi (BRF) was demonstrated in previous work (Ibáñez et al. 2008, 2011). The main component, Zn, is an essential element that is largely abundant in nature, thus available at low cost. Zinc has appropriate fungicidal properties and has low mammalian toxicity. Zinc does not bioaccumulate in the trophic chain and is not concentrated by vegetable species above levels found in soil (de Azevedo and da Matta 2003). The fungicidal mechanism of zinc is similar to that of copper.
Heavy metals affect the physiology of wood-rotting fungi (Yonni et al. 2002; Jarosz-Wilkolazka and Gadd 2003; Baldrian et al. 2005), including their metabolism, spore germination, growth and differentiation, and enzyme activity at both gene expression and translation levels. Whereas some bivalent heavy metals are essential for fungal metabolism, the biological role of others is still unknown. Whether essential or not, heavy metal ions are toxic to fungi at higher concentrations. Fungal strain tolerance to heavy metals varies according to growth stage and vegetative or reproductive form (Gadd 1993).
The inhibitory effect of Zn2+ on the growth and decolourisation activity of two WRF (P. chrysosporium and P. atropurpurascens) was studied in previous work (Ibáñez et al. 2011). At low concentrations, Mn2+ has been reported to induce several physiological activities of fungal metabolism; however, high concentrations ultimately lead to growth inhibition, as also reported for other metal species (Singh and Chen 2008; Gadd 2010). The concentration at which inhibition occurs is dependent on the particular species. Fungi like P. chrysosporium are capable of growing even at 20 mM of Mn2+ in a nutrient medium, but are completely inhibited at 50 mM of Mn2+. In contrast, less resistant species like Inocutis sp. are completely inhibited at Mn2+ concentrations as low as 5 mM (unpublished data).
The effect of metal supplements in the substrate was further studied for three additional strains of WRF (Inocutis sp., G. trabeum and T. versicolor) with widely varying metal tolerances and Mn2+ requirements (Ibáñez et al. 2011). These results were compared with those reported previously for P. chrysosporium and P. atropurpurascens to determine possible synergistic or antagonistic effects of the two cations on certain physiological traits.
Finally, the effects of Zn2+ and Zn2+/Mn2+ on enzyme activity and spore germination were studied in species with widely different ligninolytic systems (P. chrysosporium and P. atropurpurascens), to assess how the fungicidal properties of the study solution varied with growth stage.
Materials and methods
Fungal strains, nutrient media, culture conditions and metal supplements
Four WRF strains and one BRF strain were studied. The selection of basidiomycetes was consistent with the relative prevalence of WRF over BRF on Uruguayan hardwoods. Phanerochaete chrysosporium (1556, DZM), Punctularia atropurpurascens (H2126 CCMFQ), Inocutis sp. (HO214 CCMFQ), Trametes versicolor (H2140 CCMFQ) and Gloeophyllum trabeum (H2130 CCMFQ) strains were obtained from the fungal collection Cátedra de Microbiología, Facultad de Química of Uruguay's Universidad de la República.
Strains were maintained on 1·25% malt extract and 2% agar (MEA) (Oxoid Ltd, Basingstoke, UK). The medium was sterilised by autoclaving at 121°C for 15 min. Plastic Petri dishes (85 mm inner diameter) containing unsupplemented or supplemented MEA were inoculated with a mycelial plug (5 mm diameter) obtained from actively growing mycelia. Dishes were incubated at 28°C for 21 days.
Solutions of analytical grade ZnSO4 and MnSO4 (Sigma Chemical Co.) were prepared to supplement the MEA medium in order to determine the effect of Zn2+ and the combined effect of Zn2+ and Mn2+ on mycelial growth, dye-decolourizing ability, enzyme activity and conidium germination. Malt extract agar was amended with a ZnSO4 solution to obtain six different concentrations in the final medium (0·5, 1, 2, 4, 5 and 10 mM of Zn2+). A second series of tests was supplemented with a 2∶1 mixture of ZnSO4 and MnSO4 to obtain the same final Zn2+ concentrations in the medium as above (i.e. also containing 0·25, 0·5, 1, 2, 2·5 and 5 mM of Mn2+). Unsupplemented MEA medium was used as the control. Each treatment was replicated on three plates per test fungus.
Mycelial growth measurements
Mycelial growth was determined by measuring the diameter (mm) of colonies (Hatvani and Mécs 2003), on three plates per test fungus, for each of the studied strains growing on non-supplemented (control) and metal/s supplemented media. For each of the above experiments, the lowest concentration completely inhibiting growth [(minimum inhibitory concentration (MIC)] was determined based on the resulting plots (Borokhov and Rothenburger 2000). Initial growth rates (mm/day) were calculated based on the average slope of growth plots between days 1 and 9, and later compared with the corresponding value on day 21.
Wood decay test
Blocks of Pinus taeda L. and Eucalyptus grandis Hill ex Maiden sapwood (23×11×6 mm) free of defects and fungal colonization were used to test the effects of Zn2+ and Mn2+ on resistance to decay (Bravery 1978; Brown et al. 1991). Blocks were oven-dried at 103±2°C for 48 h and weighed (nearest 0·001 g). They were impregnated with solutions containing ZnSO4/MnSO4 (2∶1 molar ratio) at concentrations of 1·5/0·75% and 3/1·5% (six blocks were treated with each solution for each fungal strain). Impregnation was conducted using a Bethell process (modified to 0·5-h initial vacuum at −0·6 bar, 1-h pressure at 10 bar and 0·5-h final vacuum at −0·6 bar). The blocks were weighed again after treatment to determine the net metal retention. Dry, untreated wood blocks were used in control cultures.
Treated (and control) wood blocks were stored under non-drying conditions at 60±2°C for 36 h to allow for fixation to take place. They were later sterilised in an autoclave at 121°C for 15 min and then placed on mycelia of the tested basidiomycetes growing actively on MEA in plastic dishes. An untreated control and two identically treated blocks were placed in each dish, using three dishes for each test (i.e. equal supplement retention and strain). Dishes were incubated at 23±2°C and 60–70% relative humidity for 6 weeks. Following incubation, blocks were weighed, oven-dried at 100±2°C and weighed again (final dry substrate weight). Fungicidal effectiveness was assessed using the corrected weight loss values to take into account the non-fungal associated weight losses that may occur during incubation (initial dry treated substrate weight – final dry substrate weight/initial dry treated substrate weight). Average weight loss (%) (six blocks treated with each solution, for each fungus) was reported for each retention level.
Tests of decolourisation capacity
The effect of added metals on ligninolytic activity was studied based on tests of Poly R-478 and Remazol Brilliant Blue R (RBBR) decolourisation (Borokhov and Rothenburger 2000; Eichlerová et al. 2005; Palmieri et al. 2005). Three dishes containing control and metal/s supplemented MEA at the different tested concentrations (as used in MIC tests) were added with 200 mg L−1 of Poly R-478 and with RBBR (Sigma Chemical Co.). Plates were incubated at 28°C for 21 days. Dye-decolourisation capacity was measured by estimations of the surface area (mm2) of decolourised zones (Hatvani and Mécs 2003). For each of the studied dyes, triplicate measurements were made for each metal/s concentration and strain. For each test, the lowest concentration that inhibited decolourisation [minimum decolourisation-inhibiting concentration (MDIC)] was determined (Borokhov and Rothenburger 2000; Hadacek and Greger 2000).
Tests of enzyme activity
P. chrysosporium was cultured in liquid carbon-limited medium (CLB), following the procedure used by Vanden Wymelenberg et al. (2009) except that trans-aconitic acid was replaced by 20 mM of acetate buffer (pH 4–5). P. atropurpurascens was cultured on 1·25% malt extract medium. Both cultures were incubated at 28°C for an incubation time previously determined to result in maximum manganese peroxidase (MnP) activity. For each strain, three aliquots were taken from the supernatant of the culture medium following filtration and MnP activity at 35° was immediately measured spectrophotometrically. Maximum activity was determined for each strain based on the resulting plots of enzymatic activity versus incubation time.
The effect of metals on MnP activity was determined for each of the two strains, using the same metal/s concentrations as for MIC determinations. Incubation conditions were the same as those used for the construction of the above curve and the enzyme determinations performed when the enzyme maximum activity had been detected.
Lignin peroxidase activity was determined according to Tien and Kirk (1988), measuring the extent of H2O2-dependent oxidation of veratryl alcohol to veratraldehyde. Whereas the alcohol exhibits no absorbance at 310 nm, the aldehyde absorbs strongly at this wavelength. Thus, 200 μL of 2 mM veratryl alcohol, 180 μL of sample (filtered supernatant of each fungal culture) and 120 μL of 2·5 mM H2O2 were added to 1380 μL of 0·1M sodium tartrate buffer (pH 3).
Manganese-dependent peroxidase activity was determined based on the oxidative coupling of 3-methyl-2-benzothiazolinone hydrazone and 3-(dimethylamino) benzoic acid (Castillo et al. 1994). Mn-dependent peroxidase catalyses the formation of a deep purple compound, presumably an azo-dye, in the presence of 1260 μl of 0·1M succinate-lactate buffer, 300 μL of 6·6M 3-(dimethylamino) benzoic acid, 100 μL of 1·4 mM 3-methyl-2-benzothiazolinone hydrazone, 30 μL of 20 mM MnSO4, 300 μL of sample and 10 μL of 10 mM H2O2, showing a broad absorption band between 575 and 600 nm, with a peak at 590 nm.
Laccase activity was determined in 3 mL of reaction medium containing 10 μL of syringaldazine (1·6 mg mL–1) in 2·690 μL of 100 mM phosphate buffer at pH 6 and 300 μL of sample. Quinone formation was monitored at 526 nm (Galliano et al. 1991).
All the above activities were expressed in mUnits, where one unit was defined as the amount of enzyme required to form 1 μmole of product in 1 min. All the assays were performed from triplicate cultures.
Conidial germination
Conidial germination of P. chrysosporium or P. atropurpurascens was assessed on a thin layer of MEA supplemented with ZnSO4: 0·08, 0·16, 0·25, 0·33, 0·66, 1, 1·5 or 2 mM of Zn2+; or MnSO4: 0·04, 0·12, 0·25, 0·5, 0·8, 1, 1·5 or 2 mM of Mn2+; or ZnSO4–MnSO4: 0·08–0·04, 0·16–0·12, 0·25–0·12, 0·33–0·25, 0·5–0·25, 0·66–0·5, 1–0·5, 1·5–0·7 or 2–1 mM of Zn2+ and Mn2+ respectively, placed on a sterile slide. Each slide was inoculated with one drop of conidial suspension (107 conidia/mL) obtained from the culture dish by extraction in 9% saline followed by filtration through cheese cloth for complete removal of mycelia (Guiraud and Galzy 1980). Four slides were prepared from two different cultures of each of the tested strains, supplements and metal concentrations. The inoculated slides were incubated in Petri dishes at 28°C and observed under a phase-contrast microscope (×40) until conidial germination was apparent. They were further monitored for 48 h to check for possible morphological changes.
Statistical analysis
Fungal growth and decolourisation data were subjected to Mann–Whitney U tests (Mann and Whitney 1947), at α = 0·05, using SPSS 9·0 for Windows. Mean values of replicates (in triplicate for fungal growth and decolourisation tests; eightfold repeats for conidial germination tests; sixfold repeated for tests of wood preservation against fungal action) are shown.
Results and discussion
Effect of metal supplements on mycelial growth on culture medium
Initial growth rates varied greatly. Unlike T. versicolor, whose mycelium covered the entire dish area after 10 days’ incubation in control medium, Gloeophyllum trabeum and Inocutis sp. control cultures had not covered the entire plate area after 21 days of incubation.
Fungal behaviour in the presence of Zn2+ and Zn2+/Mn2+ supplements in the culture medium varied according to fungal strain and metal concentration (Fig. 1). Like P. atropurpurascens (Ibáñez et al. 2011), G. trabeum and Inocutis sp. were more sensitive to the presence of metals than T. versicolor or P. chrysosporium. Results show that 1 mM of Zn2+ completely inhibited the growth of Inocutis sp., while G. trabeum and T. versicolor were inhibited at 2 mM. Similar results had been found in preliminary experiments (unpublished data) using Cu2+ supplements, suggesting a similar fungicidal potency of added Zn2+ and Cu2+ supplements in the culture medium.
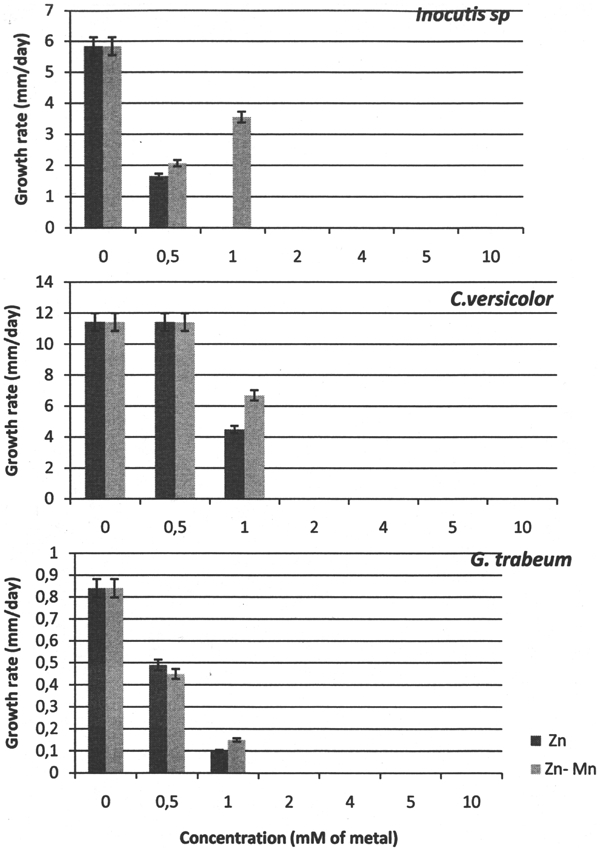
Initial growth rate of tested basidiomycete species as function of concentration of salt solution (mM of metal: Zn2+ or Zn2+/Mn2+)
In preliminary studies, Mn2+ alone as a supplement (0·25–20 mM) either did not affect or slightly stimulated the growth of certain basidiomycetes. P. chrysosporium showed a higher tolerance (MIC) of up to 50 mM of Mn2+ (Ibáñez et al. 2011).
While the combined effect of Zn2+ and Mn2+ on the growth of T. versicolor did not differ from that of Zn2+ alone at Zn2+/Mn2+ concentrations of 0·5 and 0·25 mM respectively, the effect of 0·5 mM of Mn2+ offset that of 1 mM of Zn2+, further resulting in a 48% increase in initial growth rate compared with Zn2+ alone. Similar stimulatory behaviour was found for Inocutis sp. (Fig. 1). G. trabeum did not appear to react appreciably to the inclusion of Mn2+ in a Zn2+ supplemented media.
Although the presence of Mn2+ generally reduced the inhibitory effectiveness of Zn2+, reductions occurred to a lesser degree in the three basidiomycetes of this study compared with the values noted for P. atropurpurascens and P. chrysosporium in previous work (MICs of 4 and 5 mM of Zn2+ respectively; Ibáñez et al. 2011). Following 21 days’ incubation, the MIC of the Zn2+ supplement was 1 mM of Zn2+ for Inocutis sp. and 2 mM of Zn2+ for T. versicolor and G. trabeum; whereas that of the Zn2+/Mn2+ (2∶1) supplement was 2 mM (of Zn2+) for all the strains under study. According to our experience, this is due to different strains’ behaviour.
While both Zn2+ and Mn2+ are essential in some primary and secondary metabolic processes (Jellison et al. 1997), these fungi rely on intra- and/or extra-cellular detoxification processes that are activated when a critical metal concentration is reached (Gadd et al. 2001; Gadd 2010). Such defence mechanisms against the presence of high Zn2+ concentrations may differ from those used by the fungi when a critical concentration of Mn2+ is reached. These results suggest that the presence of Mn2+ in the culture medium may lead to reduced Zn2+ bioavailability.
Effect of metal supplements on fungal growth using a wood substrate
All three fungi produced weight losses exceeding 20% on either Pinus taeda or Eucalyptus grandis substrate (Bravery 1978; Brown et al. 1991). These results indicate that conditions were suitable for fungal attack.
The highest retention used in these tests (29 kg m−3 of solution in pinewood and 26 kg m−3 of solution in eucalyptus) resulted in complete wood protection. These retentions were obtained with a Zn2+/Mn2+ solution with a concentration the same as the concentration in Zn2+/Mn2+ of the acidic solution obtained from the battery recycling process. These levels are higher than those obtained by Freeman and McIntyre (2008) (6·4 kg m−3) for copper. The MIC of the Zn2+/Mn2+ supplement was 3% Zn2+ (458 mM of Zn2+). These results are consistent with previous work, with a 2∶1 mixture of pure ZnSO4 and MnSO4 at a concentration of 3%Zn2+ (1·5% of Mn2+) fully inhibiting mycelial growth in P. chrysosporium and P. atropurpurascens cultures on pinewood substrate; whereas the wood substrate was heavily degraded at lower retentions (Ibáñez et al. 2011).
The three basidiomycetes evaluated were more highly sensitive (i.e. lower MIC) to the presence of Zn2+/Mn2+ than were P. atropurpurascens or P. chrysosporium (Ibáñez et al. 2011), when assayed on wood substrate. This was expected as their sensitivity was higher on MA.
The Zn2+/Mn2+ supplement appeared to be more active in eucalyptus wood than in pinewood. These differences may be directly ascribed to lower metal retention in eucalyptus (Table 1). Also, consistently higher weight losses were found for pinewood with values similar to those of the untreated controls.
Weight losses caused by Inocutis sp. and G trabeum on Pinus taeda and Eucalyptus grandis*
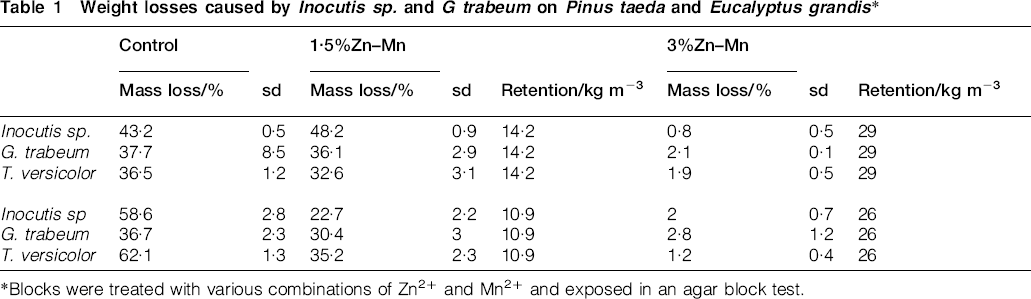
*Blocks were treated with various combinations of Zn2+ and Mn2+ and exposed in an agar block test.
The above differences may be explained in terms of the chemical composition of these two wood types, in particular the difference in the relative percentage and the type of lignin, as well as relative percentages of carbon sources used by the fungi (i.e. cellulose, hemicellulose and sugars). As fungal response under conditions of toxicity stress depends on the substrate's nutritional characteristics (Gadd et al. 2001), it is likely that eucalyptus may be a more suitable substrate for the growth of these basidiomycetes. On-going research at our laboratory is aimed at determining possible chemical reactions between the Zn2+ and/or Mn2+ supplements and major wood constituents, in particular lignin.
Effect of metals on decolourisation ability
Potential wood preservatives should be capable of suppressing both normal growth as well as oxidative activity (Borokhov and Ruthenburger 2000). Under natural conditions, the non-specific and non-stereoselective oxidative extracellular system of WRF targets the polyphenolic constituents of lignin and other, structurally similar molecules. The system is composed of laccases, lignin peroxidase and manganese peroxidase, all of which act in combination with H2O2-producing oxidases and secondary metabolites (Kirk and Farrell 1987).
The effect of the studied ions on the ligninolytic systems of the studied basidiomycetes was analysed in terms of their ability to degrade polymeric dyes (Freitag and Morrell 1992; Borokhov and Rothenburger 2000). Normal growth results in dye-decolourisation around and below growth zones, the total area of decolourised zones being generally smaller than that of the corresponding colony, consistent with the fact that decolourisation is a secondary metabolic activity of mature mycelia.
A preliminary study showed that the addition of RBBR and Poly R-478 did not significantly affect (α = 0·05; p>0·8) the initial growth rate of basidiomycetes fungi. However, initial decolourisation rates differed among the tested fungi. While T. versicolor had decolourised 100% of the dish area by incubation day 6, Inocutis sp. did not decolourise appreciably until day 8.
Results obtained in the presence of RBBR were identical to those for Poly R-478 (Table 2). This does not agree with previous works reporting RBBR, an anthraquinonic dye, to be more readily degraded than Poly R-478. This assumption is based on a preliminary comparison of the structural complexity of the two dyes, including location and number of functional groups on aromatic rings, electronic distribution and charge density (Hernandez–Luna et al. 2008).
Minimum decolourisation-inhibiting concentration values of tested metal supplements corresponding to RBBR and Poly R-478 decolourisation by tested basidiomycetes

The presence of Zn2+ or Zn2+/Mn2+ completely inhibited both RBBR and Poly R-478 decolourisation in cultures of Inocutis sp. In contrast, Mn2+ offset the effect of Zn2+ in T. versicolor, counter balancing the effect of the latter. The MDIC of the Zn2+/Mn2+ supplement was twofold higher than that of Zn2+ alone, regardless of dye type. These results are consistent with preliminary tests showing that copper led to the decolourisation of both dyes at concentrations below 5 mM in cultures of these basidiomycetes.
No appreciable decolourisation was noted in G. trabeum cultures over the range of conditions tested.
In addition to a clear antagonistic effect of Mn2+, as evidenced in cultures of T. versicolor, these results show that, in cultures of any of the four WRF, mycelial growth and ligninolytic enzyme activity were affected to a similar extent by the presence of Zn2+.
Effect of metals on enzyme activity
Extracellular fungal enzymes interact directly with heavy metals, where metal-detoxification mechanisms associated with intracellular functions are not available (Baldrian 2003). These enzymes may be inactivated by different mechanisms, such as active site modifications, the hindrance of particular functional groups or the displacement of enzyme metal co-factors.
MnP activity has been known to depend on the Mn2+ concentration in the medium (Vyas and Molitoris 1995; Singh and Chen 2008). Maximum MnP activity was noted on day 16 in the control culture of P. chrysosporium and on day 9 in the P. atropurpurascens control.
The ligninolytic system of P. atropurpurascens was capable of decolourising RBBR and Poly-R in the presence of metal concentrations that were even higher than the MIC of the different supplements, i.e. limited growth of the basidiomycete did not result in the inhibition of decolourisation (Ibáñez et al. 2011). With a view to determining which of the three main ligninolytic enzymes (MnP, LiP and Lac) accounted for the above behaviour, the ligninolytic activity of each enzyme was determined at the various metal concentrations used in the study. These results were compared with those obtained for P. chrysosporium, a reportedly fast growing basidiomycete.
Zn2+ alone affected the activity of the three MnPs in the two studied fungi even at the lowest concentration tested (Table 3). Mn2+ acted antagonistically with Zn2+ in cultures of P. chrysosporium and P. atropurpurascens. However, the observed levels of MnP activity increased in the presence of Mn2+ only for P. chrysosporium. The Zn2+/Mn2+ supplement was associated with an increase in MnP activity to levels that were even higher than those obtained in the unsupplemented control medium for P. chrysosporium. In contrast, MnP activity of P. atropurpurascens was virtually absent in cultures growing on supplemented media.
Enzymatic activity (mean±sd) of P. chrysosporium and P. atropurpurascens (mU mL−1) at different concentrations of added metals in final medium
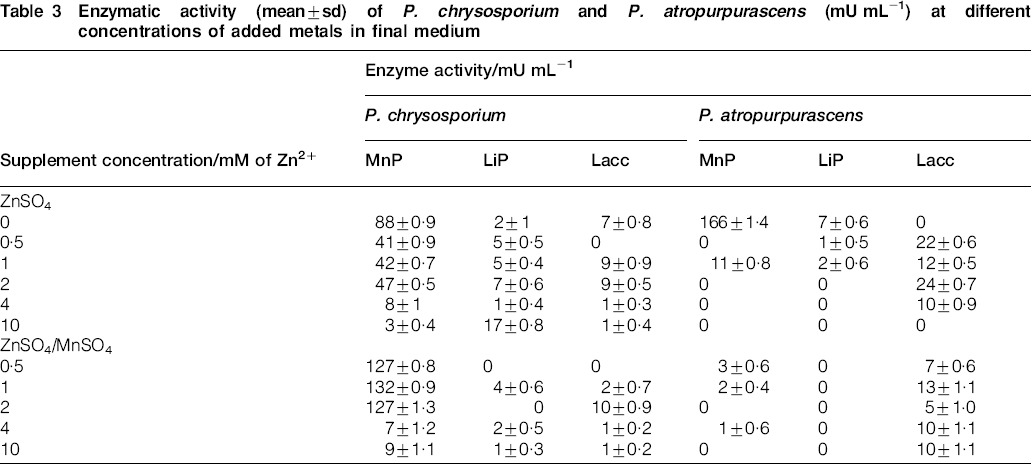
Lignin peroxidase activities were extremely low under all tested conditions.
Laccase activity values were consistently low for P. chrysosporium. In contrast, laccase activity increased in P. atropurpurascens cultures supplemented with Zn2+ (with respect to the unsupplemented control) to values even higher than those found in cultures growing on media supplemented with both cations. Laccase activity in cultures of P. atropurpurascens was detected at Zn2+ concentrations even greater than those at which no growth was observed. Zn2+ appears to induce the expression of laccase, even in non-growing mycelia. Laccase activity was also found using the Zn2+/Mn2+ supplement at a Zn2+ concentration in the medium of up to 10 mM, i.e. the MDIC of the supplement.
In sum, MDIC values obtained by decolourisation of polymeric dyes were inconsistent with the ligninolytic activity levels determined spectrophotometrically. MnP activity was consistently found at higher metal concentrations than was suggested by the MDICs of the corresponding supplements.
Effect of metals on conidium germination
Conidial germination was noted after 12 h incubation in cultures of P. chrysosporium and after 17 h in P. atropurpurascens. Table 4 shows the results (%) obtained at 19 h incubation, when conidia had germinated on the un-supplemented control media. P. atropurpurascens conidia appeared to show a higher sensitivity to the presence of metals.
Percentage of germinated conidia at different metal concentrations at 19 h of incubation for P. crhysosporium and P. atropurpurascens
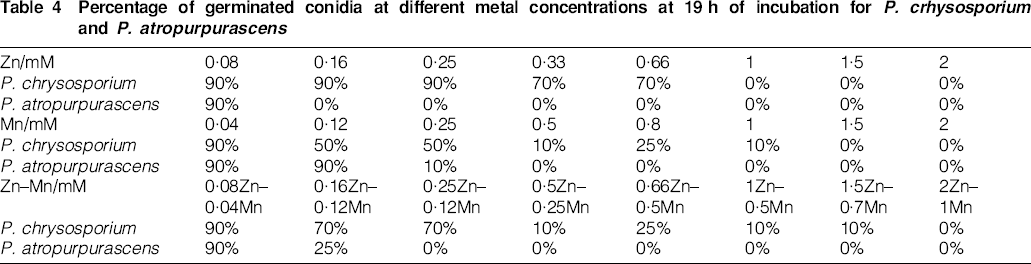
The addition of Mn2+ slightly induced growth, decolourisation ability and enzyme activity in cultures of the tested fungi, but did not appreciably reduce spore germination for the two WRF. Nonetheless, a certain effect of Mn2+ was evidenced by an apparent increase in the tolerance (in terms of MIC) of P. chrysosporium to the combined-metal supplement in terms of Zn2+ concentration.
Additions of 1·5 mM of either metal alone or a 2∶1 (Zn2+/Mn2+) mixture of both led to complete inhibition of germination for both species, indicating a protective effect of these salts against initial colonisation of wood by basidiomycete conidia.
The overall physiological response to the presence of metals in the substrate results from the combined action of several metal toxicity and tolerance mechanisms. Also particularly relevant in the synthesis of metal-binding peptides and proteins, the regulation of homeostasis and toxic responses relies on mechanisms of efflux, transport and internal compartimentation (Gadd 1993, 2001, 2010). Reaching a critical metal ion concentration, defence mechanisms were triggered by the studied fungi against Zn2+ and Mn2+ at a concentration – depending on species – within the range of values tested.
Conclusion
Pure ZnSO4 and ZnSO4/MnSO4 supplements used in the culture medium were found to inhibit the physiological traits of the five WRF considered for the analysis. Regardless of species, the addition of Zn2+/Mn2+ mixed salts (2∶1 molar ratio) completely inhibited the growth and ligninolytic activity of the five fungi at a Zn2+ concentration (MIC and MDIC respectively) slightly higher than that of the supplement containing Zn2+ alone that led to the same result.
Unlike their effects on spore germination (determined for P. chrysosporium and P. atropurpurascens), Zn2+ and Mn2+ were shown to have antagonistic effects on the growth and enzymatic activity of the five basidiomycetes being considered, with Mn2+ leading to an increase in a particular strain's tolerance to Zn2+ and/or a reduction in its initial growth rate of each particular strain.
Moreover, comparing the results obtained for the pure sulphates with those obtained for a 2∶1 Zn2+/Mn2+ solution resulting from battery recycling (MIC of 2 mM of Zn2+), the combined effect of trace heavy metals contained in the latter on the growth and ligninolytic activity of wood-rotting basidiomycetes was reflected in lower MIC and MDIC values than those obtained for cultures supplemented with the pure sulphates, indicating a higher toxicity.
Adjustments may be made in the Mn2+ concentration with a view to the definition of a wood preservative formulation of adequate toxicity for particular environments (depending on local or regional prevalence, wood preservative use or application, among other factors). However, as these salts are prone to leaching when in contact with moist soil, the addition of boron may be nonetheless required.
Footnotes
Acknowledgements
This work was supported by a financial grant from CSIC (Sectoral Commission for Scientific Research of Uruguay's Universidad de la República) and ISUSA (Industrias Sulfúricas del Uruguay, SA).