
Abstract
Fungal cellulases have been shown to be less efficient in modified wood than in untreated wood (Lekounogou et al. 2008; Venås 2008). However, Verma and Mai (2010) showed that cellulase efficacy is partly restored in 1,3-dimethylol-4,5-dihydroxyethyleneurea (DMDHEU) treated wood by pre-treatment with Fenton's reagent, simulating the oxidative degradation phase in initial brown rot decay. In this study, we examined whether Fenton derived hydroxyl radicals (·OH) and cellulases are able to cleave polysaccharides in furfurylated and acetylated wood and to what extent enzyme efficacy is increased by oxidative pre-treatment of these materials. The results show that fungal cellulases were able to degrade acetylated wood and that the cellulase efficacy was increased by oxidative pre-treatment by 20%, which is half of the increase in untreated wood. Furthermore, the results indicate that poly(furfuryl alcohol) is degraded by Fenton derived ·OH. This indicates a possible route for the eventual degradation of modified wood.
Keywords
Introduction
Modified wood is an alternative to preservative treated wood and involves the action of a chemical, biological or physical agent upon the material, resulting in increased durability and/or other desired enhancements. During furfurylation of wood, furfuryl alcohol polymerises inside the wood and may bind to lignin (Nordstierna et al. 2008). Acetylation of wood is commonly achieved by reacting wood with acetic anhydride, which results in esterification of the accessible OH-groups in the wood cell wall (Rowell et al. 1994). Furfurylation and acetylation confers decay resistance and has been shown to be non-toxic (Pilgård et al. 2010; Lande et al. 2004). The mechanism behind the increased decay resistance in modified wood is not fully understood (Hill 2006; Ringman et al. 2013).
Brown rot fungi degrade wood through oxidative and enzymatic action (Goodell et al. 1997; Arantes et al. 2012). The fungus, growing in the lumen, induces production of ·OH inside the wood cell wall through secreting Fe3+-reductants and H2O2 (Goodell et al. 1997). In the wood cell wall, the reductants reduce Fe3+ to Fe2+, which react with the H2O2 and produce ·OH in the Fenton reaction (equation (1)) (Goodell et al. 1997; Goodell et al. 2006; Fenton 1894)
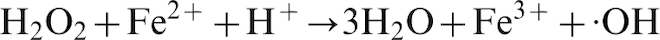
Verma and Mai (2010) suggested that Fenton derived ·OH is able to cleave DMDHEU treated wood polysaccharides in a similar manner as in brown rot, based on that brown rot cellulase efficacy is enhanced after oxidative pre-treatment. Their study also shows that fungal cellulases are able to degrade polysaccharides in DMDHEU treated wood to almost the same extent as in untreated wood when using oxidative pre-treatment, even though enzyme efficacy in modified wood materials that has not been oxidatively pre-treated is substantially lower than in untreated wood (Lekounougou et al. 2008; Venås 2008; Verma and Mai 2010). It is not known whether ·OH is able to cleave polysaccharides in other kinds of modified wood. Furthermore, it is not known whether fungal cellulase efficacy can be restored in these wood materials as it is in DMDHEU treated wood.
The aim of this study was to examine whether ·OH is able to cleave polysaccharides in furfurylated wood and to what extent enzyme efficacy is restored by oxidative pre-treatment in these materials.
Experimental methods
Wood material
Southern yellow pine sapwood was cut to 30×10×5 mm samples, leached according to EN84 (EN 84 1996) and dried. A portion of the samples was furfurylated in the laboratory at NFLI (Ås, Norway) according to Lande et al. (2004) with the exception that the samples were soaked in the modification mixture overnight to ensure full penetration, to an average weight per cent gain (WPG) of 91·5%. Another portion was acetylated at the laboratory at SP Wood Science (Borås, Sweden) according to Larsson-Brelid (1998), to an average WPG of 22%.
Oxidative and enzymatic degradation of wood powder
The degradation tests were carried out in triplicates and performed according to Verma and Mai (2010) with minor changes. For in vitro simulation of oxidative degradation followed by enzymatic degradation, 0·5 g of wood powder was incubated in 12·5 mL 100 mM sodium acetate (NaAc) buffer (pH 5·0) with 0·5 mM (NH4)Fe(SO4)2 and 1% H2O2 in 23°C on a shaker (150 rev min−1) for 24 h. The tubes were centrifuged at 7850 rev min−1 for 5–10 min and the supernatant was removed. The wood powder was washed three times with sterile deionised water. 10 mL sterile deionised water and 0·125 mL Kieralon antiperox (BASF, Limburgerhof, Germany) was added and the tubes were incubated for 3 h on a shaker (250 rev min−1) in 23°C. The tubes were again centrifuged at 7850 rev min−1 for 5–10 min and the supernatant was removed. The wood powder was washed three times with sterile deionised water. 10 mL NaAc buffer and 28 U of Celluclast 1·5 L (Sigma-Aldrich Sweden, Stockholm, Sweden) were added and the tubes were incubated on a shaker (250 rev min−1) in 23°C for ∼160 h.
In order to ensure the same experimental conditions, all samples were washed according to the routine above. Treatment with Kieralon antiperox was shown not to affect cellulase activity (data not shown), therefore this step was left out in the other samples. For in vitro simulation of oxidative degradation, 0·5 g of washed wood powder was incubated in 12·5 mL NaAc buffer in 50 mL tubes with 0·5 mM (NH4)Fe(SO4)2 and 1%H2O2 in 23°C on a shaker (250 rev min−1) for ∼160 h. For in vitro simulation of enzymatic degradation, 0·5 g of washed wood powder was incubated in 12·5 mL NaAc buffer with 28 U of Celluclast 1·5 L in 23°C on a shaker (250 rev min−1) for ∼160 h.
The pH of NaAc buffer inoculated with wood meal was 5·05±0·05 for all wood treatments.
Reducing sugar assay
Reducing sugar content was assayed according to Miller (1959) and Verma and Mai (2010). Each tube was assayed in triplicates. The tubes were centrifuged at 7850 rev min−1 for 5–10 min. 1 mL supernatant was transferred to a 2 mL tube. When necessary, the samples were diluted in NaAc buffer. 1 mL of DNS solution (0·63% 3,5-dinitrosalicylic acid, 0·57% phenol, 0·5%NaHSO3, and 2·14%NaOH) was added and the tubes were incubated at 100°C for 10 min. The tubes were subsequently cooled down to room temperature and the absorbance was measured at 540 nm. A standard curve was prepared from a glucose solution.
Results and discussion
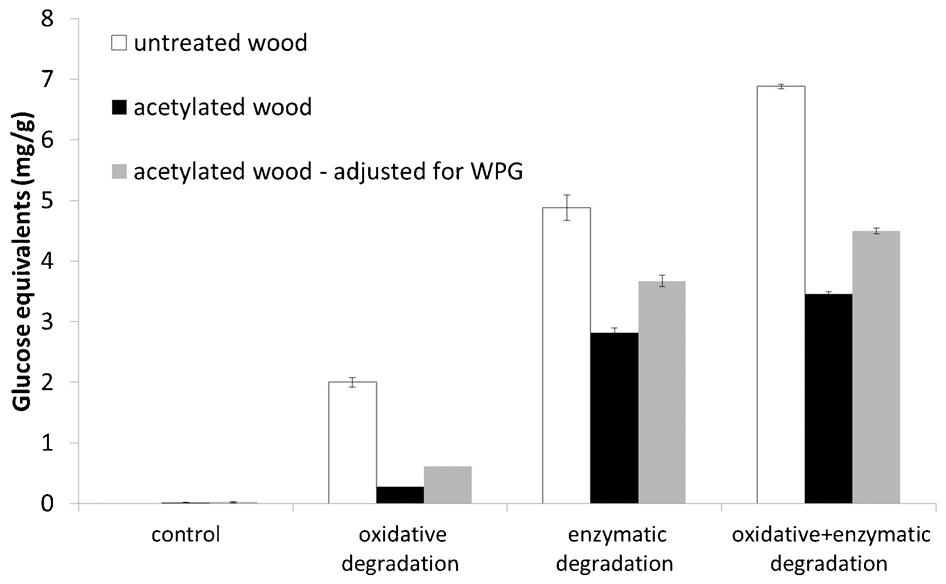
The results from the untreated wood are in accordance with previously published results by Verma and Mai (2010), except for the results after treatment with Fenton's reagent (Fig. 1). In our study, almost 2 mg reducing sugars per g wood meal were released after treatment with Fenton's reagent while in the study by Verma and Mai's levels of released reducing sugars are lower than 1 mg g−1 which was also almost the same as the control. However, in both studies, the efficacy of the cellulases was increased after pre-treatment with Fenton's reagent. In this study the increase was approximately 40%.
Efficacy of in vitro simulated oxidative, enzymatic and oxidative+enzymatic degradation

Furfurylated wood meal (from left to right): untreated, treated with Fenton's reagent, treated with cellulases, treated with Fenton's reagent followed by cellulases
The results from the acetylated wood meal show that brown rot cellulases were able to degrade acetylated wood (Fig. 1). Furthermore, the efficacy of the cellulases was increased after pre-treatment with Fenton's reagent by approximately 20%. The levels were overall lower in the acetylated wood. Also after adjustment for the average WPG of the acetylated wood, the levels of released reducing sugars were lower in the acetylated wood than in untreated wood (Fig. 1).
This is in accordance with the study by Verma and Mai (2010), where the levels of released reducing sugars were shown to be lower in DMDHEU treated wood compared to untreated wood. When treating modified wood meal with Fenton's reagent, lower levels of released reducing sugars may be due to that ·OH is reacting also with the modification chemical. When treating modified wood meal with cellulases, lower levels of released reducing sugars may be due to that the acetyl groups are physically hindering the cellulases to gain access to the polysaccharides. The increased efficacy of the cellulases seen after pre-treatment with Fenton's reagent may in that case be due to that the ·OH has cleaved off the acetyl groups.
The increase in cellulase efficacy after pre-treatment with Fenton's reagent is higher in DMDHEU treated wood than in untreated wood (Verma and Mai 2010), while the increase in acetylated wood was lower than the untreated wood. This may be due to that the WPG of the DMDHEU treated wood meal used was 7%, while the WPG of the acetylated wood meal in this study was 22%. If this is true, a more effective treatment with Fenton's reagent would increase cellulase efficacy in acetylated wood even further.
When treating furfurylated wood meal with Fenton's reagent, the supernatant turned brown, while the supernatant in the control was uncoloured (Fig. 2). Even after washing with water and treatment with cellulases the brown colour remained, although somewhat lighter. This is an indication that the Fenton derived ·OH was able to degrade poly(furfuryl alcohol).
However, the degraded poly(furfuryl alcohol), possibly in the form of furfurals or as sugar contaminations from the production process, reacted with the DNS reagent and thus made it impossible to separate the reducing sugars released from the holocellulose from the poly(furfuryl alcohol) remnants. We were, however, able to confirm previous results by Venås (2008) that brown rot cellulases are able to degrade furfurylated wood.
Today, four established theories of mode of action of modified wood are prevalent: fungal enzyme inefficacy due to non-recognition (Rowell 2005); fungal enzyme inefficacy due to lack of water at glycosidic bonds (Rowell 2005; Rowell et al. 2009); reduced flow of fungal molecules into the wood cell wall due to micropore blocking (Hill et al. 2005); and inhibition of diffusion of fungal molecules due to insufficient amounts of moisture in the wood cell wall (Papadopoulos and Hill 2002; Boonstra and Tjeerdsma 2006). To what extent the different modes of action contribute to decay resistance is unknown; however, the biochemical mechanisms of brown rot decay support the theory of inhibition of diffusion due to moisture exclusion (Ringman et al. 2013).
The results from this study suggest that poly(furfuryl alcohol) can be degraded by Fenton derived ·OH. This may explain why furfurylated wood is eventually degraded. At a place in the wood cell wall where the WPG is locally low enough to allow diffusion of the fungal molecules needed for degradation, ·OH might form. The radicals might attack the modification chemicals inside the wood cell wall and thereby the area with low WPG will expand and even more ·OH might form. Alternatively, very low amounts of ·OH may be formed in the lumen and attack the wood cell wall from the outside.
Conclusion
Brown rot cellulases were able to degrade acetylated wood. Cellulase efficacy was increased after treatment with Fenton's reagent, but not to the same extent as in untreated wood. This shows that Fenton derived ·OH facilitates cellulase degradation, possibly by depolymerisation of polysaccharides and/or removal of acetyl groups from wood polymers. Furthermore, the results from this study indicate that Fenton derived ·OH is able to degrade poly(furfuryl alcohol). These results makes it tempting to speculate that the eventual degradation of furfurylated wood might be due to low levels of ·OH formation in the lumen or at places inside the wood cell wall where the WPG is low enough to allow for diffusion. Once ·OH has been formed, poly(furfuryl alcohol) may start to be degraded.
Footnotes
Acknowledgements
The authors gratefully acknowledge financial support from The Swedish Research Council Formas 213-2011-1481 and COST actions FP0904 and FP1303. The authors would also like to thank Lena Brive and colleagues at SP Chemistry for their support in the chemistry lab.