
Abstract
Objective
To determine changes on craniofacial growth morphometrically in newborn mice with cleft palate induced by retinoic acid.
Design, Setting, Participants, Interventions
Gestation day 10 or 12 pregnant female C57BL/6N mice were given a single dose of all-trans retinoic acid (atRA) by gastric intubations via oral gavage. Sixty newborn mice with cleft palate (CP), 52 without CP from the experimental group, and 30 without CP from the control group were collected, and lateral cephalograms were taken of all of the mice.
Main Outcome Measures
Cephalometric analysis of the craniofacial skeleton was performed by means of a personal computer.
Results
Inhibition of craniofacial growth was found in the experimental groups but not in the control groups. In the maxillary bone and mandible, the amount of growth was significantly reduced.
Conclusions
These results suggest that craniofacial growth is inhibited in newborn mice with cleft palate induced by retinoic acid.
To provide optimal care for individuals with clefts of the palate, clinicians need to understand the separate effect of the cleft and of the surgical repair on the growth of the face and dental arches. The study of “unoperated” individuals with cleft should enable investigators to explore how the “cleft” face expresses itself when not influenced by surgical repair (Bishara et al., 1985). However, surgical care is becoming more available to populations all over the world. As a result, there are limited opportunities at present to examine large numbers of individuals with unoperated clefts (Isiekwe and Sowemimo, 1984). And most studies conducted on individuals with unoperated clefts have one or more of following limitations: small sample size; wide age distribution; mixture of unoperated, partially operated, and late operated individuals; and finally, a mixture of individuals with different cleft types (Bishara et al., 1986; Shetye and Evans, 2006). In order to overcome these limitations, animal models with cleft palate were introduced in the analysis of craniofacial malformations in present study.
Cleft palate (CP) is one of the major malformations induced by retinoic acid in both rodents and humans. Mouse models are particularly appropriate as models for human palate malformation because early facial development and morphology of the palate and lip is similar to that of humans (Trasler, 1968; Diewert and Wang, 1992; Parsons et al., 2008). These mouse models offer an opportunity to compare both the embryology and the response to environmental manipulation of genotypes predisposed or resistant to the defect (Juriloff and Trasler, 1976; Gong and Eulenberg, 2001).
The administration of pharmacologic doses of all-trans retinoic acid (atRA) during pregnancy results in a wide range of craniofacial and limb malformations (Sulik et al., 1988; Alles and Sulik, 1989; Satre and Kochhar, 1989). During skeletal development, derangements in levels of retinoic acid result in numerous, severe abnormalities in a number of skeletal tissues, including the appendicular and craniofacial skeletons (Means and Gudas, 1995; Collins and Mao, 1999; Schneider et al., 2001). High doses of retinoic acid can produce bilateral clefting of the lip and palate. This occurs via growth inhibition of the frontonasal and maxillary processes of the developing craniofacial complex and may be mediated by inhibition of sonic hedgehog expression (Helms et al., 1997). It may also be a result of impaired neural crest migration early during development (Yasuda et al., 1986; Johnston and Bronsky, 1995). Nowadays, several lines of evidence suggest that retinoic acid (RA) is a morphogenetic signaling molecule in vertebrate embryos and, in particular, has a role in organizing the body pattern along the anteroposterior axis. RA affects proliferation of cranial neural crest cells as well as oronasal mesenchymal cells. This appears to be related to the production of craniofacial malformations in rodents and humans (Watanabe et al., 1990). It has also been demonstrated that the teratogenic effects of RA are attributable to excessive cell death (Sulik et al., 1988; Abbott et al., 1990).
Various methodologic procedures have been utilized in cleft palate studies, but it has not been until the latter part of the last century that cephalometrics was extended to the nonhuman primate, despite its common usage in human dentistry for many years. Descriptions of cephalometric analysis of craniofacial growth are available for rats (Kiliaridis et al., 1985) and mice (Duke et al., 1996; Bloom et al., 2006), and dental malocclusions have been induced experimentally in monkeys (Harvold et al., 1973; Carlson, 1983) and beagles (Bardach et al., 1988). Therefore, x-ray based cephalometric analysis could be applied to the studies about prenatal experimentation of cleft palate and postnatal analysis of the induced defects.
This study uses C57BL/6N mice, which develop CP with retinoic acid (Kochhar and Johnson, 1965), to investigate the relationship between craniofacial malformations and variation and risk of abnormal palate formation.
Methods
Animals and atRA Treatment
Mature male and female C57BL/6N mice purchased from the medical animal center of Sun Yat-sen University (Guangzhou, China) were housed at a temperature of 22°C with a 12-hour light/dark cycle and were given food pellets and tap water ad libitum. They were mated overnight, and the date of vaginal plug detection was designated as gestation day (GD) 0. GD10 or GD12 pregnant female mice were given a single dose of atRA (100 mg/kg; Sigma-Aldrich, St. Louis, MO) by gastric intubation via oral gavage. The atRA was dissolved in corn oil. Control animals were given the equivalent volume of corn oil alone. These animal studies conformed with the standards for use of laboratory animals established by the Institute of Laboratory Animal Resources, U.S. National Academy of Science.
In the GD10 group, RA induced cleft palate in 66.67% of the fetuses (38/57), while in the GD12 group RA induced cleft palate in 44.94% of the fetuses (40/89). Sixty 1-day-old mice with cleft palate (30 from GD10 and 30 from GD12) comparable to Veau II cleft in humans, and 52 newborn mice without CP (12 from GD10 and 40 from GD12) from the experimental group and 30 without CP from the control group were collected and decapitated. Heads were fixed in 1% formaldehyde, and lateral cephalograms were obtained using a cephalostat for standardized action.
Radiographic Technique
In order to position each mouse's head in exactly the same position for each radiograph, a specially constructed cephalostat was used. Heads were positioned by inserting plastic extensions into the external auditory meatus of the skulls. The lateral and submental-vertex cephalographs were taken with dental occlusal film (DF-50, Eastman Kodak, Rochester, NY) under an electric condition of 7 mA and 60 Kvp with an exposure time of 0.15 seconds. After developing, cephalograms were scanned by means of an imaging scanner (GT-9000, Epson, Tokyo, Japan) and enlarged 5×. Cephalometric analysis was performed with the use of a personal computer according to the modified method of Kiliaridis et al. (1985). Landmarks and measurement items in Figure 1 were established for the present study (Fujita et al., 2004). In order to compensate radial distortion in the x-rays, the known distance of 5 mm on the radiopaque metal stick was used to calibrate images prior to bone measurement. Measurements were made with the Winceph 8.0 imaging system. Each measurement for each radiograph was made three times (intraclass correlation coefficient range from 0.79 to 0.94), and the mean of the three was used as the measurement for that skull.
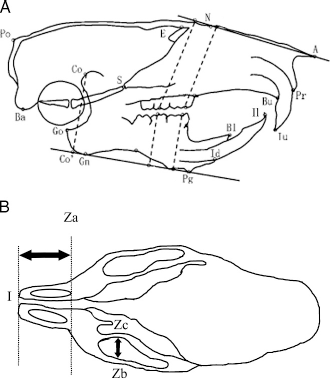
Landmarks and measurement items used for the cephalometric analyses. A: Lateral view. B: Ventral view.
Statistical Analysis
One-way analysis of variance (ANOVA) was used to assess the differences of the three groups (CP-GD10, CP-GD12, and control) on the same indices. Multiple comparison tests (Student-Newman-Keuls test, SNK-q) were used to assess differences between measurements for pairs of groups. The mean values for each cephalometric parameter of CP and non-CP (NCP) mice in the experimental groups were then compared using the Student's t test. Differences were considered significant if p < .05.
Results
Radiographs of craniofacial skeletons from neonates are presented in Figure 2. Direct cephalometric measurements and the results of the one-way ANOVA are presented in Table 1. The results of multiple comparison tests for pairs of groups are reported in Table 2. The results of Student's t test are reported in Table 3.
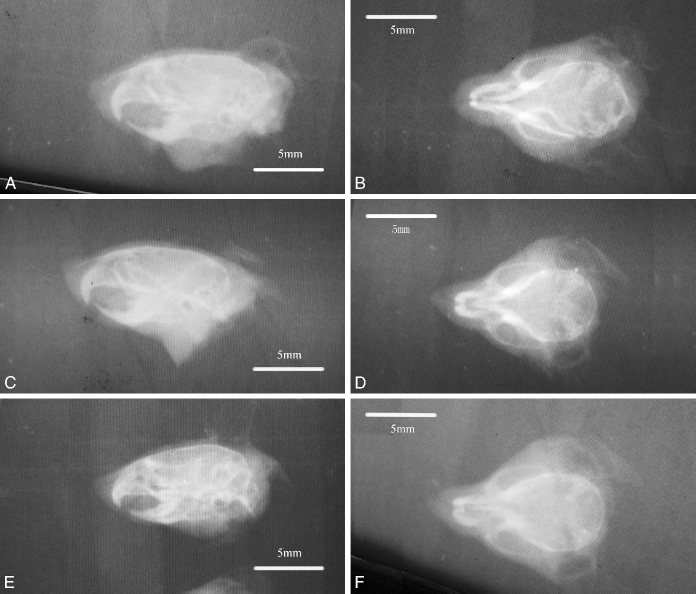
Radiographs of craniofacial skeletons from neonates of control group. A: Lateral view. B: Ventral view. CP-GD10 group (C, D) and CP-GD12 group (E, F).
Cephalometric Measurements of Mice by Group With Statistical Analysis
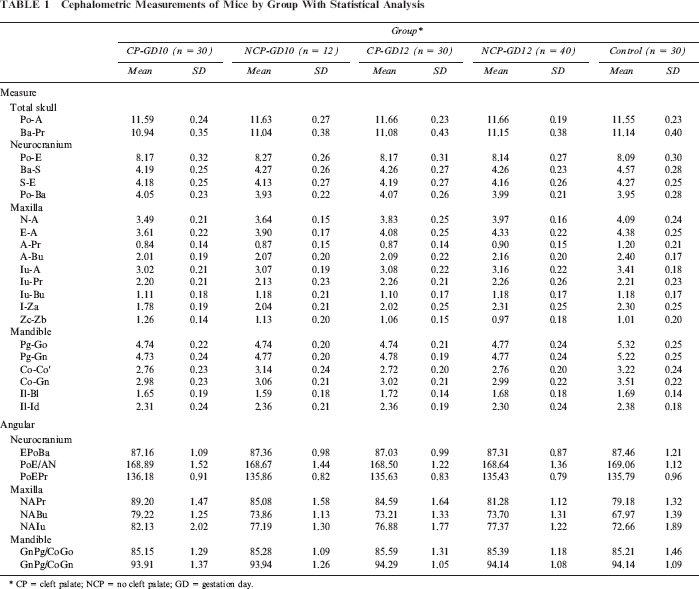
CP = cleft palate; NCP = no cleft palate; GD = gestation day.
Results of Multiple-Comparison Tests for Pairs of Groups Between Measurements of Mice †
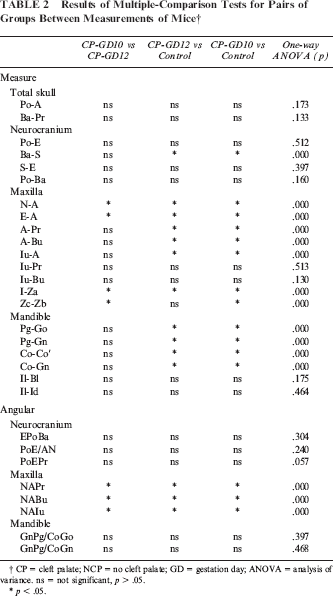
CP = cleft palate; NCP = no cleft palate; GD = gestation day; ANOVA = analysis of variance, ns = not significant, p > .05.
p < .05.
Results of Student's t Test for Pairs of Groups Between Measurements of Mice †
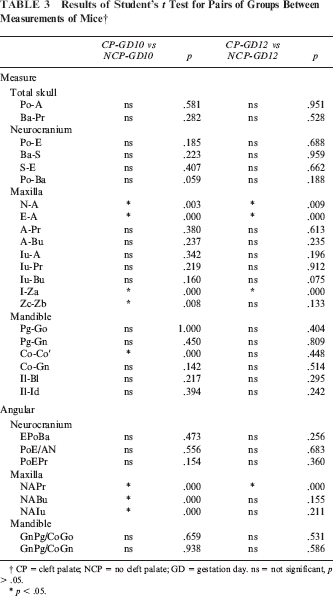
CP = cleft palate; NCP = no cleft palate; GD = gestation day. ns = not significant, p > .05.
p < .05.
No significant differences in total skull size or neurocranium size were found between the experimental (CP-GD10 and CP-GD12) and control groups, except for Ba-S distance. All cleft palate CP mice were significantly smaller than their control littermates, but the differences between the GD10 and GD12 groups were not significant.
ANOVA revealed significant statistical differences between groups CP-GD10, CP-GD12, and control in the measurements of maxillary length (N-A, E-A). Significant differences were also found between groups CP-GD10 and CP-GD12 in these measurements. The greatest mean maxillary length measurement was observed in the control group, followed by groups CP-GD12 and CP-GD10.
ANOVA revealed significant statistical differences between groups CP-GD10, CP-GD12, and the control in maxillary height (A-Pr, A-Bu, Iu-A), except for Iu-Pr and Iu-Bu. The maxillary height measurements in the control group were significantly higher than the measurements in groups CP-GD10 and CP-GD12, but the differences between the CP groups were not significant.
In the two experimental groups, similar to the maxilla, the amount of mandibular growth was reduced as compared with the control groups in terms of dimensional changes. In the CP group, four measurement items (Pg-Go, Pg-Gn, Co-Co′, Co-Gn) were significantly smaller than in the control group, but the differences between the CP-GD10 and CP-GD12 groups were not significant. No significant differences were found in II- BI and Il-Id between three groups. The nature of mandibular growth and the differences observed in those affected by cleft palate were almost the same as those previously described for the maxilla.
Two measurements were taken (right and left) for snout length (I-Za) and zygomatic arch width (Zb-Zc) in the submental-vertex cephalograph. Group CP-GD10 had the significantly shortest maxillary length and widest facial width in the three groups. While CP-GD12 had shorter maxillary length than the control, there was no significant difference between CP-GD12 and control in face width.
No significant differences in all of the angular measurements of the neurocranium were found among the three groups. N-A-Pr, N-A-Bu, and N-A-Iu were significantly larger in the CP group, and significant differences were also found between groups CP-GD10 and CP-GD12 in these measurements. In the experimental group, it was thus shown that point A is located more posteriorly, as noted by the reduced maxillary dimensions mentioned in the preceding linear measurements. No significant differences were found in the shape of the mandibular or gonial angle (GnPg/CoGo, GnPg/CoGn) between the experimental and control groups.
The result of the Student's t test revealed that maxillary length (N-A, E-A) and snout (I-Za) of CP mice were shorter than NCP mice. In GD10 groups, CP mice also had a shorter Co-Co′ and longer Zc-Zb distance and bigger N-A-Pr, N-A-Bu, and N-A-Iu angles than NCP mice. In the GD12 group, CP mice showed bigger N-A-Pr than NCP.
Discussion
The impact of clefting on the morphogenesis of patients with cleft palate has been described by a number of investigators (Sasaki et al., 2004). Orthodontists have observed that compared with their normal peers, patients with cleft palate have a rather collapsed maxilla instead of an arch bridge supported by bilateral maxillary tubercles. It may be the result of the V-shaped defect in the hard palate and asymmetric muscles, which presumably cause a changed and unbalanced stress distribution (Boyne and Sands, 1976). Therefore, we hypothesize that cleft palate may be an intrinsic cause of craniofacial malformation.
Attempts have been made in the past to study the growth pattern in unoperated cleft patients to identify causes of the abnormal craniofacial growth, and there have been conflicting opinions. Some studies combined different cleft types (Isiekwe and Sowemimo, 1984), included patients with lip repair (Ortiz-Monasterio et al., 1966), compared cleft patients with normal patients from different racial groups, or used a sample with an extended age group (Mars and Houston, 1990). Because of the circumstances in which data on unoperated individuals are collected, a number of these limitations are beyond the control of the investigator. In the present study, C57 mice were used as an appropriate animal model for CP induced by retinoic acid. Therefore, the untreated cleft and normal individuals were from very similar genetic backgrounds; the high suitability of the controls is an important advantage in a study of oral clefts because variability is more likely to be related to the issues being considered. And this animal model has some advantages compared with human patients, such as rapid growth, ease of care, and the availability of the data from other investigators that could be used for comparison with our studies. The present study was designed to examine the role of animal models in investigations of craniofacial growth altered by cleft palate induced with retinoic acid. An examination of unoperated cleft palate mice eliminates surgery as a cause of any abnormal growth observed and helps determine whether the abnormal growth pattern observed in treated cleft patients is an intrinsic defect or a side effect of treatment in future study.
In this study, we succeeded in inducing an experimental model of CP in newborn mice by using retinoic acid that was not lethal to the mice. The results of this study suggest that cleft palate exerts a substantial influence on craniofacial growth, due to the fact that suppression of craniofacial growth was significantly induced, as compared with control groups, by GD10 or GD12 groups in animals at 1 day old. Furthermore, the influence was more prominent in the GD10 group; thus, early effect at gestation period seems more important for growth control. These results also suggest that retinoic acid might have a teratogenic effect not only of cleft palate but also of reduced craniofacial growth (Hendrickx et al., 1980).
No significant differences in total skull size or neurocranium size were found between the experimental (CP-GD10 and CP-GD12) and control groups. In this study, environmental factors may greatly influence the growth of the neurocranium. It may be supposed that cleft palate induced with retinoic acid has no influence on the growth of the neurocranium.
From morphometric analyses with lateral cephalograms, it was shown that cleft palate substantially influences craniofacial growth in newborn mice. All cleft palate mice were significantly smaller than their control littermates for all of the craniofacial measurements, except for Iu-Pr, Iu-Bu, Il-Bl, and Il-Id. In particular, remarkable changes in the height and length of the maxilla and mandible were found in the experimental groups, whereas no significant differences in the total size of the skull or neurocranium were found between the experimental and control groups, except for Ba-S. In this study, the growth of the maxilla, which was mainly determined by sutural growth, was inhibited in the experimental groups, as was that of the mandible, determined by membranous and cartilaginous growth. Mice in the CP-GD10 and CP-GD12 groups have shorter maxillary length (N-A, E-A) and snout (I-Za), compared with NCP mice respectively. But the difference of other indices was not so significant. These results indicated that mice with CP have more poor growth in maxillary length than those without CP, although they all had the same retinoic acid induction in the embryonic period. From these considerations, it might be possible that CP induced with retinoic acid may affect sutural growth in some area of the maxilla. It is of interest to consider whether the smallness of the craniofacial structure is an intrinsic genetic condition induced with retinoic acid or whether the smallness is due to altered growth related to changes in structure and function due to the cleft. The difference in cranial base length observed in this study could theoretically be related to the altered craniofacial structure and forces of related muscles (Wang and Mao, 2002; Shetye and Evans, 2006).
In the angular measurements, the increased N-A-Pr, N-A-Bu, and N-A-Iu angle in CP subjects can be explained geometrically by the more posterior location of the A point and the surrounding dentoalveolar process. Similar findings of reduced maxillary dimensions in patients were noted by other researchers (Isiekwe and Sowemimo, 1984; Yoshida et al., 1992). Retrusion of the maxilla is a common finding in most patients with cleft palate or with cleft lip and palate. Several causes are suggested for the retrusion. These can be classified into two categories: (1) intrinsic primary defects or (2) interference with growth secondary to surgical repair. Clinical experience and scientific studies have shown that early repairing surgeries dramatically reduce facial convexity by retropositioning the premaxilla. It seems that the pressure created by the repaired lip and palate is enough to correct the position of the projected premaxilla. However, there are few studies in the literature that relate intrinsic primary defects of craniofacial growth in cleft palate patients because of the limitations mentioned in the introduction section of this article. The reduced maxillary dimensions in this study could be related to intrinsic primary defects of craniofacial growth in cleft palate induced with retinoic acid (Semb, 1991; da Silva Filho et al., 2003). Anterior crossbite is a common finding in patients with clefts. In CP groups, the maxillary incisor position was found to be more posterior than the control group, and this might be because of the collapse of the maxilla due to the gap of cleft palate bone. Although the results reflect inhibition of bone growth in the experimental groups, no significant differences in the gonial angle were found between the experimental and control groups. Gonial angle may be influenced by mechanical and functional changes in occlusion (Sugiyama et al., 1999).
In this study, significant differences were also found between groups CP-GD10 and CP-GD12 in some measurements (N-A, E-A, N-A-Pr, N-A-Bu, N-A-Iu, I-Za, and Zb-Zc). Bishara reported a wide range of variation in the maxillofacial growth within each cleft group, even within the same cleft type (Bishara et al., 1985). Many studies reported different cleft palate types could be formatted if the retinoic acid was administered orally on different gestation days (Abbott et al., 1989; Abbott and Birnbaum, 1990; Horie and Yasuda, 2001; Campbell et al., 2004). The induction of cleft palate by atRA varies depending on the stage of development at which exposure occurs. In vivo studies have indicated that after exposure of embryonic mice to RA on GD10, abnormally small palatal shelves form. After exposure on GD12, shelves of normal size form but fail to fuse as the medial cells proliferate and the normal growth and differentiation process of palatal mesenchymal cells is inhibited (Abbott et al., 1989, 1990). The differences dimensions between groups GD10 and GD12 might be because retinoic acid was administered on different embryonic phase and induced different cleft palate type.
Conclusions
Inhibition growth existed for three dimensions development of craniofacial structure in newborn mice with cleft palate induced by retinoic acid. The severity of craniofacial malformations depended on the stage of development, GD10 or GD12, at which RA exposure occurred.