
Abstract
Objective
Orofacial clefts such as cleft palate (CP) and cleft lip (CL) and/or cleft palate (CL/P) are the most common congenital anomalies of the head and neck. The aims of the present study were to evaluate the possible association between CDH1 (rs11642413 and rs16260) and MSX1 (rs12532 and rs3775261) gene polymorphisms and nonsyndromic cleft lip and/or cleft palate (NS-CL/P) in a sample of the Iranian population.
Design and Setting
This case-control study was performed on 100 subjects with NS-CL/P and 100 healthy unrelated control subjects. Tetra amplification refractory mutation system-polymerase chain reaction and multiplex polymerase chain reaction were used to detect the single-nucleotide polymorphisms.
Results
There was a significant difference between NS-CL/P subjects and control subjects regarding CDH1 rs16260 C > A polymorphism, and the rs16260 AC as well as the rs16260 AA genotypes were associated with NS-CL/P susceptibility (odds ratio [OR] = 3.02, 95% confidence interval [CI] =1.51 −6.00, P=.001; and OR = 8.05, 95% CI = 1.72–37.75, P=.002, respectively). No significant difference was found between the groups regarding CDH1 rs11642413 polymorphism. Although MSX1 rs3775261 polymorphism was not a risk factor for the disease, the rs12532 AG and rs12532 GG genotypes were associated with NS-CL/P risk (OR = 2.82, 95% CI = 1.55–5.15, P=.001; and OR = 8.42, 95% CI = 2.26–31.29, P=.004, respectively).
Conclusion
Our data suggest that CDH1 and MSX1 gene polymorphisms are risk factors for susceptibility to NS-CL/P in a sample of the Iranian population. Larger studies are required to validate our findings.
Cleft lip and/or cleft palate (CL/P) is a congenitally developmental disorder resulting from defects in growth and patterning of the facial primordial (Letra et al., 2010). The prevalence of orofacial clefts such as cleft palate (CP) and cleft lip (CL) with or without palate is approximately 1/500 to 1/2000 (Murray, 2002; Hashmi et al., 2005). It has been reported that the incidence of CL and CP in Iran is approximately 1/1000 and lower compared with other countries (Khazaei et al., 2011). Oral clefts are complex malformations proposed to be caused by environmental factors, genetic influences, and interactions among these factors. A large number of experimental and epidemiologic findings suggest that cigarette smoking, alcohol drinking, dietary deficiencies, and exposure to phenytoin (a medication used to control seizures) during pregnancy could all increase the risk of nonsyndromic cleft lip and/or palate (NS-CL/P) (Hernandez-Diaz et al., 2000; Loffredo et al., 2001; Wyszynski and Wu, 2002; Puho et al., 2007; Romitti et al., 2007). Several genome-wide association studies have identified specific susceptibility loci on chromosomal regions as a genetic risk factor for NS-CL/P (Birnbaum et al., 2009; Grant et al., 2009; Beaty et al., 2010; Mangold et al., 2010; Ludwig et al., 2012).
There is evidence that genetic polymorphisms in various genes, including TGF-beta3 (Saleem et al., 2012), BMP4 (Suazo et al., 2011), SLC6A4, TPH2, and SERPINA6 (Mostowska et al., 2011), RUNX2 (Wu et al., 2012), AXIN2 (Letra et al., 2012), ZNF533 (Wu et al., 2011), CDH1 (Song and Zhang, 2011), and MSX1 (Otero et al., 2007; Park et al., 2007), have been involved in the etiology of NS-CL/P.
The E-cadherin gene (CDH1), located at the 16q22.1 locus, comprises 16 exons separated by 15 introns. The encoded protein belongs to the family of cell-cell adhesion molecules and plays a key role in craniofacial morphogenesis, palatal fusion, and tumor growth inhibition (Kerrigan et al., 2000; Kordi-Tamandani et al., 2010).
The MSX1 gene, located on chromosome 4p16.2, contains two exons interrupted by an intron of approximately 1.6 kb and encodes a protein of 297 amino acids (Jezewski et al., 2003). The mature protein product plays a role in craniofacial development, in particular, odontogenesis and tumor growth inhibition. Mutation within this gene has been associated with NS-CL/P (van den Boogaard et al., 2000; Jezewski et al., 2003).
Genetic risk factors for NS-CL/P may differ between different populations. Consequently, repeating previously reported genetic associations (Letra et al., 2009; Huang et al., 2011; Song and Zhang, 2011; Singh and Ramu, 2012) in different populations is required to determine the associations of the genetic risk in each population.
The current study was carried out to determine the impact of CDH1 rs11642413 (intronic variation) and rs16260 (promoter variation; −160 C/A) as well as MSX1 rs12532 (3′-UTR variation) and rs3775261 (intronic variation) gene polymorphisms on risk of NS-CL/P in a sample of the Iranian population.
Materials and Methods
The study group consisted of 100 subjects with CL/P (61 male and 39 female) with an average age of 12.12 years (range, 1 year to 54 years) and 100 healthy subjects (61 male and 39 female) with an average age of 12.03 years (range, 1 year to 51 years). There was no significant difference between the groups regarding sex and age (P > .05). All participants were unrelated to each other.
The Ethics Committee of the Zahedan University of Medical Sciences approved the project, and written informed consent was obtained from all participants. Blood samples were collected in Na-EDTA tubes from patients and healthy control subjects and stored at −20°C until DNA extraction. Genomic DNA extraction from blood samples was carried out as described previously (Hashemi et al., 2010a).
To detect CDH1 rs11642413 and MSX1 rs12532, we performed tetra amplification refractory mutation system-polymerase chain reaction (T-ARMS-PCR), which is a simple and rapid method for detecting single-nucleotide polymorphisms (SNPs) (Hashemi et al., 2010b; Hashemi et al., 2011; Hashemi et al., 2012a; Hashemi et al., 2012b). The CDH1 and MSX1 genomic sequences (NT_010498.15 and NT_006051.18, respectively) were obtained from the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov). The polymorphisms were searched and primers for T-ARMS-PCR were designed (Table 1). In the T-ARMS-PCR method, two external primers (control band) and two inner primers (allele specific primers) are used. This method simultaneously amplifies both alleles in a single polymerase chain reaction (PCR) tube.
Primers Used to Determine CDH1 rs11642413 and MSX1 rs12532 Genetic Polymorphisms Using T-ARMS-PCR

PCR was performed using commercially available PCR premix (AccuPower PCR PreMix, BIONEER, Daejeon, South Korea) according to the manufacturer's recommended protocol. Into a 0.2-mL PCR tube containing the AccuPower PCR Pre-Mix, 1 μL template DNA (˜100 ng/μL), 1 μL of each primer (10 μM), and 15 μL DNase-free water were added. The PCR cycling condition were 5 minutes at 95°C followed by 30 cycles of 30 seconds at 95°C, 30 seconds at 54°C for CDH1 rs11642413, 20 seconds at 63°C for MSX1rs12532, respectively, and 30 seconds at 72°C, with a final extension of 10 minutes at 72°C. The PCR products were analyzed by electrophoresis on a 2% agarose gel containing 0.5 μg/mL ethidium bromide and visualized by transillumination with ultraviolet (UV) light; a photograph was then taken (Fig. 1).
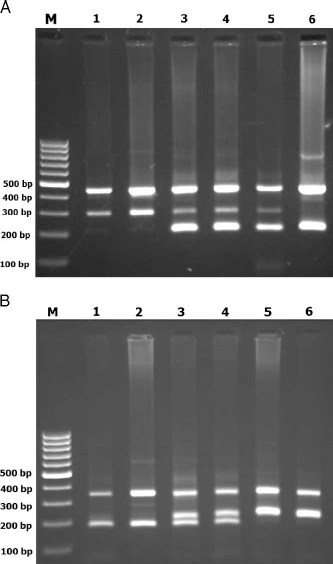
Photograph of the PCR products of CDH1 rs11642413 (A) and MSX1 rs12532 (B) gene polymorphism. In the T-ARMS-PCR method, two external primers (control band) and two inner primers (allele specific primers) are used. For the CDH1 rs11642413 polymorphism, the product sizes were 210 bp for A allele, 297 bp for G allele, and 449 bp for control band (A). M: DNA marker; Lanes 1, 2: rs1642413 GG; Lanes 3, 4, 5: rs1642413 AG; Lane 6: rs1642413 AA. The product sizes for detecting the MSX1 rs12532 polymorphism were 176 bp for G allele, 228 bp for A allele, and 353 bp for control band (B). M: DNA marker; Lanes 1,2: rsl2532 GG; Lanes 3,4: rs12532 AG; Lanes 5, 6: rs12532 AA.
To evaluate CDH1 rs16260 and MSX1 rs3775261 polymorphisms, we designed a multiplex PCR. The primers used are shown in Table 2. Two tubes were used to determine rs3775261 and rs16260 genotypes simultaneously. Tube 1 contained command primers, A allele CDH1 rs16260 primer, and C allele MSX1 rs3775261 primer, while tube 2 contained command primers, C allele CDH1 rs16260 primer, and A allele MSX1 rs3775261 primer (Table 2). The PCR cycling condition were 5 minutes at 95°C followed by 30 cycles of 30 seconds at 95°C, 23 seconds at 61°C, and 30 seconds at 72°C with a final step at 72°C for 10 minutes to allow for complete extension of all PCR fragments. The PCR products were analyzed by electrophoresis on a 2% agarose gel containing 0.5 μg/mL ethidium bromide and visualized by transillumination with UV light; a photograph was then taken (Fig. 2). We regenotyped approximately 20% of the samples to verify the initial results. The check confirmed the previous genotyping results by 100%.
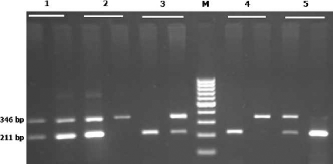
Representative multiplex PCR products resolved by agarose gel electrophoresis to detect the CDH1 rs16260 and MSX1 rs3775261 polymorphisms. The PCR products were 211 bp for the C and A allele of the CDH1 rs16260 polymorphism and 346 and 349 bp for the C and A allele of the MSX1 rs3775261 polymorphism, respectively. M: Marker; Lane1: rs3775261 CA, rs16260 CA; Lane 2: rs3775261 CA, rsl6260 AA; Lane 3: rs3775261 AA, rs16260 CA; Lane 4: rs3775261 AA, rs16260 AA; Lane 5: rs3775261 CC, CDH1 rs16260 CA.
Oligonucleotides Used to Detect CDH1 rs16260 and MSX1 rs3775261 Polymorphisms Using Multiplex PCR

Statistical Analysis
Statistical analysis of the data was performed using the SPSS 18.0 software (SPSS for Windows, SPSS, Inc., Chicago, IL). Demographics and biochemical parameters between the groups were analyzed by independent sample t test for continuous data and χ2 test for categoric data. The associations between genotypes and NS-CL/P were estimated by computing the odds ratio (OR) and 95% confidence interval (95% CI) from logistic regression analyses. The Bonferroni correction was applied by multiplying P values by the number of SNPs analyzed.
Results
Of the 100 patients, 43 had CL, 26 had CL with CP (CLP), and 31 had CP. The distribution of genotype and allele frequencies of CDH1 and MSX1 is shown in Table 3. The frequencies of all SNPs were in Hardy-Weinberg equilibrium in case and control groups (data not shown). No significant difference was found between case and control groups regarding CDH1 rs11642413 polymorphism (χ2 = 1.23, P = .541). A significant difference was observed between the groups concerning the CDH1 rs16260, polymorphism (χ2=18.40, P =.0001). The frequencies of common homozygous rs16260 CC, heterozygous rs16260 CA, and rare homozygous rs16260 AA were 56%, 33%, and 16%, respectively, in the case group and 82%, 16% and 2%, respectively, in the control group. The findings showed that, compared with CC wild homozygote, either AC heterozygote or AA homozygote was associated with CL/P susceptibility (CA versus CC, OR = 3.02, 95% CI = 1.51–6.00, P=.001; AA versus CC, OR = 8.05, 95% CI = 1.72–37.75, P = .002). We also found that the A allele is a risk factor for CL/P (OR = 2.11, 95% CI = 1.18–3.77, P = .015).
Genotype and Allele Frequencies of CDH1 and MSX1 Genes in Patients With Nonsyndromic Cleft Lip and/or Cleft Palate (NS-CL/P) and Control Subjects*
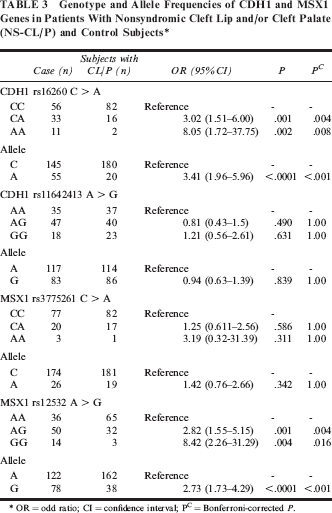
OR = odd ratio; CI = confidence interval; PC = Bonferroni-corrected P.
We evaluated the possible association between rs12532 and rs3775261 polymorphisms of MSX1 and CL/P. As shown in Table 2, there were significant differences in the genotype frequencies of rs12532 between the groups (χ2 = 19.40, P < .0001), although no significant differences of genotype frequencies for rs3775261 were found between the groups (χ2= 1.40, P = .496). Compared with rs12532 AA wild homozygote, the rsl2532AG and rs12532GG genotypes were associated with CL/P risk (OR = 2.82, 95% CI = 1.55–5.15, P = .001; and OR=8.42, 95% CI = 2.26–31.29, P = .004, respectively). Furthermore, the G allele was found to be a risk factor for CL/P (OR=2.72, 95% CI = 1.73–4.29, P < .0001).
Genotype and allele frequencies of CDH1 and MSX1 gene polymorphisms in subjects with CL, CLP, and CP and control subjects are shown in Table 4. The results indicated that CDH1 rs16260 polymorphism was associated with increased risk for CL and CLP but not CP. Although no significant association was found between CDH1 rs1642413 polymorphism and CL or CLP, the rs11642413 AG genotype was a risk for CP. The MSX1 rs12532 polymorphism, but not the MSX1 rs3775261polymorphism, was associated with risk of CL, CLP, and CP.
Genotype and Allele Frequencies of CDH1 and MSX1 Gene Polymorphisms in Subjects With Cleft Lip (CL), Cleft Lip With Cleft Palate (CLP), and Cleft Palate (CP) and Control Subjects*
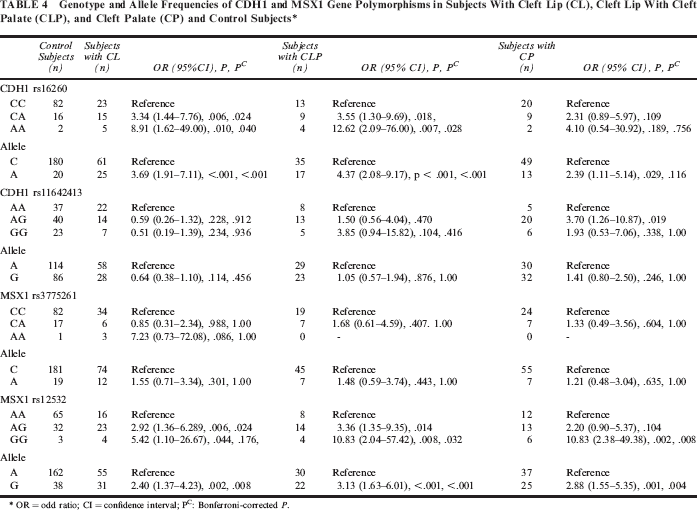
OR = odd ratio; CI = confidence interval; Pc: Bonferroni-corrected P.
Discussion
Both genetic and environmental factors may contribute to NS-CL/P. In the present study, we have investigated the impact of CDH1 and MSXlgene polymorphisms and susceptibility to CL/P in a sample of the Iranian population. A significant difference was found between patients with CL/P and control subjects regarding CDH1 rs16260 and MSX1 rs12532 polymorphisms. No significant difference was found between the groups concerning CDH1 rs11642413 and MSX1 rs3775261 polymorphisms in our population.
CDH1 plays an essential role during craniofacial morphogenesis. The rs16260 C> A polymorphism, located upstream of the transcriptional start site of CDH1 promoter, has a substantial effect on transcriptional activity and CDH1 expression (Li et al., 2000; Nakamura et al., 2002). Consequently, it is possible that this polymorphism might be a risk factor for NS-CL/P. We found that there was a significant difference between the case and control groups regarding this polymorphism, and rs 16260 CA as well as rs 16260 AA genotypes were risk factors for NS-CL/P in our population.
CDH1 in particular is a most reasonable candidate gene for NS-CL/P. The association of CDH1 mutations with CL/P was previously identified in families with hereditary diffuse gastric cancer (Frebourg et al., 2006).
CDH1 belongs to the family of cell adhesion molecules and plays a key role in cell adhesion, which is essential for normal development, including craniofacial morphogenesis and palatal fusion (Kerrigan et al., 2000). Thus, it is in close relationship with a phenomenon known as epithelialmesenchymal transformation, which happens throughout the process of palatogenesis to remove epithelial cells from the palatal medial edge epithelia and allow for mesenchymal continuity and palatal fusion. Therefore, defects in CDH1 are involved in cell-cell adhesion system dysfunction and could contribute to the complex events that cause human orofacial clefts (Hay, 1995).
There is little information concerning CDH1 gene polymorphisms in NS-CL/P. It has been reported that the rs16260 C > A polymorphism, located upstream of the transcriptional start site of CDH1 promoter, has a significant effect on CDH1 expression (Li et al., 2000; Nakamura et al., 2002). Consequently, it is likely that this polymorphism may be a risk factor for NS-CL/P. Song and Zhang (2011) found no association between CDH1 rs16260 polymorphism and risk of NS-CL/P. Although they found a significant difference between control subjects and cleft palate only (CPO) subjects regarding CDH1, the rs16260 polymorphism and rs16260 AA genotype significantly increased the risk of CPO by 5.90-fold (OR = 6.90, 95% CI = 1.47–32.40) in a Chinese Han population. Our finding showed that the rs16260 CA and rs16260 AA genotypes were risk factors for susceptibility to CL/P in our population. Letra and colleagues (2009) found that two genetic variants, rs11642413 and rs9929218, in CDH1 were associated with susceptibility to NS-CL/P in a Brazilian population.
Huang and colleagues (2011) found no significant association between NS-CL/P and rs3821949 or rs12532 in the MSX1 gene, whereas an association was observed between the P147Q variant (a missense mutation) and CLP in the case-control analysis. However, we found an association between the rs12532 polymorphism and NS-CL/P in our population. It has been found that the rs12532 A allele had a 2.08-fold increase in the risk of NS-CL/P when inherited from the father (Suazo et al., 2010).
Singh and Ramu (2012) showed a positive correlation between the MSXl:c.799G > T gene variant and NS-CL/P patients. Jagomagi and colleagues (2010) observed a significant association between the MSX1 rs6446693 polymorphism and NS-CL/P in an Estonian population. Zhang and colleagues (2008) sequenced the exon 1 of the MSX1 gene and found no mutation in the NS-CL/P subjects and unrelated healthy subjects. The microsatellite DNA marker CA repeats in MSX1 have been reported to be associated with NS-CL/P in Hunan Hans and a Colombian population (Otero et al., 2007; Wu et al., 2007). Park and colleagues (2007) observed a significant association between 1170 G/A polymorphism of exon 2 and orofacial clefts. The A allele was protective against the disease. Furthermore, MSX1 point mutations were found in 2% of cases of clefting (Jezewski et al., 2003). The discrepancy between the results may be due to complex traits of oral clefts determined by multiple, interacting genetic and environmental factors.
Our findings indicate that CDH rs16260 is associated with increased risk for CL/P (Table 3), although in subgroup analysis (Table 4) we found that CDH rs16260 is associated with CL or CLP but not CP. No significant association was found between the CDH1 rs11642413 polymorphism and CL/P, but this SNP was found to be associated with increased risk of CP. Because of the relatively small sample size in subgroup analysis, the findings should be interpreted cautiously. Future research with larger sample sizes is required to validate our findings.
In conclusion, our data suggest that the CDH1 and MSX1 gene polymorphisms are risk factor for susceptibility to CL/P in a sample of the Iranian population. Larger studies with different ethnicities are required to confirm our findings.
Footnotes
Acknowledgments
This work was supported by a dissertation grant from Zahedan University of Medical sciences. The authors thank the patients and healthy subjects who willingly participated in the study.