
Abstract
Background
Aging is associated with frequent complaints about earlier bedtimes and waketimes. These changes in sleep timing are associated with an earlier timing of multiple endogenous rhythms, including core body temperature (CBT) and plasma melatonin, driven by the circadian pacemaker. One possible cause of the age-related shift of endogenous circadian rhythms and the timing of sleep relative to clock time is a change in the phase-shifting capacity of the circadian pacemaker in response to the environmental light-dark cycle, the principal synchronizer of the human circadian system.
Methods
We studied the response of the circadian system of 24 older men and women and 23 young men to scheduled exposure to ocular bright light stimuli. Light stimuli were 5 hours in duration, administered for 3 consecutive days at an illuminance of ˜10,000 lux. Light stimuli were scheduled 1.5 or 3.5 hours after the CBT nadir to induce shifts of endogenous circadian pacemaker to an earlier hour (phase advances) or were scheduled 1.5 hours before the CBT nadir to induce shifts to a later hour (phase delays). The rhythms of CBT and plasma melatonin assessed under constant conditions served as markers of circadian phase.
Results
Bright light stimuli elicited robust responses of the circadian timing system in older people; both phase advances and phase delays were induced. The magnitude of the phase delays did not differ significantly between older and younger individuals, but the phase advances were significantly attenuated in older people.
Conclusions
The attenuated response to light stimuli that induce phase advances does not explain the advanced phase of the circadian pacemaker in older people. The maintained responsiveness of the circadian pacemaker to light implies that scheduled bright light exposure can be used to treat circadian phase disturbances in older people.
Introduction
Aging affects a number of characteristics of circadian rhythms, endogenous ∼24-hour rhythms in sleep and wakefulness, hormone levels, body temperature, performance, alertness, and other physiologic functions. Reported age-related changes in circadian rhythms include the phase (the time of a reference point of the rhythm, such as melatonin maximum or core body temperature [CBT] minimum), the amplitude (one half the difference between the maximum and the minimum), and the period (duration of a complete cycle) of circadian rhythms. These circadian rhythms are generated by a light-sensitive pacemaker located in the suprachiasmatic nuclei of the hypothalamus of mammals.
The best known example of a change in the phase of a biologic rhythm in older persons is the shift of bedtime and waketime to earlier hours. 1 Other circadian rhythms that have been reported to show an earlier circadian phase include activity, 2 CBT,1,3,4 the timing of Rapid Eye Movement sleep,5–8 and cortisol plasma concentrations.9–11
One possible explanation for this phase advance is a shortening of the period of the circadian oscillations generated by the circadian pacemaker. Because entrainment theory predicts that the phase angle difference between environmental synchronizers and circadian rhythms depends on the intrinsic period of this biologic oscillator, 12 such an age-related reduction of the intrinsic period could lead to an advance of the entrained phase of the endogenous circadian pacemaker, provided that the sensitivity of this oscillator to environmental stimuli remains unchanged. An age-related reduction of the intrinsic period of the human circadian pacemaker was hypothesized based on the results of studies in animals showing a shortening of the period of the activity rhythm in older animals. 13 This hypothesis was in part supported by data collected under experimental conditions in which humans could self-select the light-dark cycle to which they were exposed. Under these conditions, older people exhibited a shorter period of the temperature rhythm than young people.4,14 However, studies in healthy older and young humans 15 and in hamsters16,17 in which the exposure to light was carefully controlled have not detected a change in circadian period with aging. The data from these latter studies suggest that a reduction in intrinsic period is not a major contributor to age-related changes in circadian organization.
In addition to changes in circadian phase, age-related reductions in the amplitude of circadian rhythms have also been reported for body temperature,3,4,18,19 serum testosterone levels, 20 urinary electrolytes, 21 and systolic blood pressure. 22 However, recent data collected under controlled conditions suggest that age-related reductions in the amplitude of the plasma melatonin and cortisol rhythms do not occur in carefully screened healthy older individuals who have no current medical conditions and are not taking any medications at the time of study.23,24
Neither the etiology nor the functional and physiologic significance of the changes in phase and amplitude of circadian rhythms observed in older people is known. These changes in circadian organization, however, are associated with frequent complaints about early morning awakening and reduced vigilance during the daytime.25–31
Some of the changes in phase and amplitude that have been reported in older people are not simply the direct result of different social habits or different activity levels in young and in older people. The advanced phase and reduced amplitude of the temperature rhythm in healthy subjects, older than 65 years, compared with those of young subjects, persist in a highly controlled environment in which ambient illumination, posture, activity level, and feeding are maintained constant, thus controlling for potential behavioral differences between the groups.1,14,19,32 This suggests that aging is indeed associated with an earlier timing of endogenous circadian rhythmicity and with a reduced amplitude of some, but not all, physiologic variables.
Theoretically, circadian oscillators with a lower amplitude should adjust faster to a shift in environmental synchronizers (zeitgebers).33,34 However, a number of studies have suggested that as people and animals age, their ability to adjust to shifts of environmental time stimuli is reduced. Older people report greater difficulty adapting to rotating shift work schedules and transmeridian travel,35–37 situations in which the endogenous circadian pacemaker must be reset to a new external, environmental time. Older animals take longer to re-entrain (a longer time to resynchronize the endogenous circadian pacemaker) after an abrupt shift in the timing of environmental cues than younger animals in some but not all studies. 38 Slower re-entrainment of the circadian pacemaker in older animals in multiple measures has been reported, including sleep-wake cycles, 39 activity rhythms, 40 food and water intake rhythms, 38 rectal 41 and intraperitoneal temperature, 42 and the peak monoamine oxidase activity in different regions of the brain. 43 One potential cause of these difficulties in adjusting to changes in environmental stimuli in human and other animals may be changes in the endogenous circadian timing system involved in the genesis of all of these rhythms. We hypothesized that these difficulties in adjusting to changes in environmental synchronizers may be related to an altered phase-shifting capacity of the circadian pacemaker in older individuals. We further hypothesized that the advanced phase of endogenous circadian rhythmicity in older people could also be related to changes in the responsiveness of the circadian pacemaker to its main environmental synchronizer, the light-dark cycle. 44
To test directly the phase-shifting capacity of the circadian system in older people, young and older subjects were exposed to an ocular bright light stimulus and the magnitude of the resulting phase shift of their circadian systems was quantified. Similar phase response experiments in young subjects have demonstrated that the response of the human circadian pacemaker to light exposure depends on the phase of the endogenous circadian cycle at which light is administered. Light administered before the endogenous circadian temperature nadir (which in young subjects is, on average, located at approximately 6:00) shifts the endogenous rhythms to a later hour (induces phase delays). Light administered after the temperature nadir shifts endogenous circadian rhythms to an earlier hour (induces phase advances). We therefore exposed volunteers to ocular light scheduled to induce either phase advances or phase delays and compared the response in young and older people.
Materials
Subjects
24 healthy older subjects (aged 65-81 years; six male, 18 female) and 23 young men (aged 19-27 years) participated in the study. All volunteers were free from any acute or chronic medical conditions as confirmed by normal physical examination, chest radiogram (older subjects only), electrocardiogram, and detailed blood and urine laboratory tests. The screening procedure for older subjects also included an ophthalmologic exam to exclude subjects with significant cataracts or other lens pathology and a monitored nocturnal sleep episode to exclude subjects with clinically significant sleep apnea or nocturnal myoclonus. Subjects with significant sleep complaints, a history of transmeridian travel in the past 3 months, or a history of regular shift work in the past 3 years were excluded. The subjects were instructed to abstain from the use of alcohol, drugs, and stimulant-containing products for at least 3 weeks before the study. Toxicologic analysis of blood and urine was performed to verify that they were drug-free at the start of their study.
Before participation, every subject received a description of the procedure and general purpose of the experiments and gave written informed consent. The experimental protocol was approved by the Brigham and Women's Hospital Human Research Committee.
Overall Study Design
During the 3 weeks preceding admission to the laboratory, subjects maintained self-selected regular bedtimes and waketimes (within a range of ±1 hour). Their sleep-wake times for the week before admission were verified by continuous ambulatory monitoring using a portable data collection device (Vitalog PMS-8 Recorder, Vitalog, Inc, Redwood City, Calif, or AMA-21 Actigraph, Ambulatory Monitoring, Inc, Ardsley, NY).
During the laboratory study, subjects remained in an environment free from external time cues. They maintained contact with staff members trained to avoid communicating the time of day or the nature of the experimental conditions. On admission to the laboratory, subjects lived for 3 baseline nights on a normal routine. Their habitual sleep-wake times as recorded in a log during the week before study were used to schedule their baseline sleep episodes: bedtimes and waketimes were scheduled 4 hours before and after their average mid-sleep time, respectively. Immediately after the baseline days, the subject's initial circadian phase was assessed by a 26- to 37-hour constant routine (CR) procedure (described below) that continued until 6.5 hours (group A), ∼9.5 hours (group B), or ∼11.5 hours (group C) after the fitted minimum of the CBT rhythm (CBTmin). This was followed by the intervention regimen, which consisted of 3 consecutive days, each containing a 5-hour exposure to enhanced lighting (∼10,000 lux; see Intervention Regimen, below), followed by a second CR lasting 29-54 hours. The study continued for 5 more days in 15 of the young male subjects. However, only the data through the end of CR2 (first 10 days) from those 15 individuals are included in this report. Results from those 15 individuals have been reported elsewhere. 45 Data from the first CR of all subjects were included in two previous reports.1,23
During the baseline days, subjects were exposed to room light (∼150 lux) during waking episodes. During the CRs and all other wake episodes, except when exposed to the experimental bright light, subjects were exposed to dim light (10-15 lux). During all sleep episodes, the rooms were dark (∼0.03 lux).
Throughout the laboratory study, CBT was recorded at 1-minute intervals from a disposable rectal thermistor (Yellow Springs Instrument Company, Yellow Springs, Ohio), urinary volume was measured at each micturition, subjective alertness was assessed every 20 minutes with use of a linear nonnumeric 100-mm bipolar visual-analog scale,46,47 and cognitive throughput was assessed at hourly intervals by measuring the number of sums of pairs of randomly generated two-digit numbers that were completed during a 4-minute testing session. 47 Plasma samples for melatonin were collected at hourly intervals.
The CR procedure 48 was used to unmask the endogenous phase of the circadian pacemaker, as reflected by the CBT and melatonin rhythms. It consists of a regimen of enforced semirecumbent wakefulness in constant dim light (∼10-15 lux), with minimal activity. Compliance with the instruction to stay awake was monitored by a technician who was continuously present in the room. Daily individual requirements of calories, fluids, and electrolytes were calculated according to the Harris Benedict nomogram, 49 adjusted by a 30-40% activity factor, and served in identical hourly snacks.
Intervention Regimen
After 3 baseline days and determination of CBTmin during CR1, each subject underwent a light exposure regimen, which was repeated on 3 consecutive days. On each of these days, subjects were exposed to a 5-hour episode of ocular bright (∼10,000 lux) light centered 1.5 hours before (group A), 1.5 hours after (group B), or 3.5 hours after (group C) the CBTmin as assessed in CR1. The midpoint of sleep/darkness was scheduled 12 hours opposite the midpoint of the 5-hour intervention stimulus; therefore, sleep/darkness episodes were ∼6.5-11.5 hours earlier than the subjects’ normal sleep times.
During the intervention stimulus, the subjects wore clear, ultraviolet-excluding safety glasses with hard, coated polycarbonate lenses (Ultra-Spec 2000 with 4C coating; Uvex Winter Optical, Inc, Smithfield, RI). Subjects were exposed to light of ∼10,000 lux using a ceiling-mounted bank of cool-white fluorescent fixtures (North American Philips Lighting Corp, Bloomfield, NJ). The light intensity was progressively increased to this level during a 25-minute transitional period that preceded the 5-hour light exposure. It was progressively reduced to dim light during a 25-minute transitional period that followed the 5-hour light stimulus. During the light exposure, subjects were seated and were asked to gaze at a point on a wall where they received ∼10,000 lux for 10 minutes out of every 20 minutes. Their angle of gaze was unrestricted for the remaining 10 minutes. Illuminance measurements were performed at 10-minute intervals with a research photometer (analog recorder output, exponential sensitivity factor) equipped with a photopic detector and cosinor diffuser (Models IL 1700 photometer and SED(SEL)038/Y 7859/W 3339 detector; International Light, Newburyport, Mass) that was placed at the subject's forehead and directed toward the line of gaze.
Data Analyses
The phase and amplitude of the endogenous circadian pacemaker were assessed from CR CBT and melatonin data. Although CBT data were used during the experiment for real-time execution of the protocol, including determining the timing of the light exposures and sleep episodes, poststudy analyses of phase-shifts were performed using the melatonin rhythm as the phase marker, because melatonin is a more accurate marker of the circadian pacemaker. 50
CBT data were analyzed using a two-harmonic-regression-plus-correlated-noise statistical model. 51 To exclude masking effects from the previous sleep/darkness episode, the first 5 hours of CBT data collected during CR assessments were excluded from analysis. Endogenous circadian phase was defined as the weighted average of the fitted minima from single- and dual-harmonic regression models applied to CBT data. Endogenous CBT circadian amplitude was defined as one half the difference between the maximum and the minimum values from the single-harmonic fitted curve. Because estimation of endogenous circadian phase was unreliable when the endogenous circadian amplitude fell below 0.25oC, only data from subjects whose CBT amplitude exceeded that threshold 52 were used in secondary analyses; CBT data from only one older subject from the phase advance group was excluded for this reason.
Circadian phase and amplitude were also assessed using plasma melatonin collected via indwelling catheter during the CRs. Melatonin was assayed by a direct radioimmunoassay kit (Elias USA, Inc, Osceola, Wis). All samples from a single subject were run in the same assay. To exclude masking effects from the previous sleep/darkness episode, the first 5 hours of melatonin data collected during CR assessments were excluded from analysis. Melatonin phase was defined as the time of the midpoint between the upward and downward crossings of the mean value from hours 5-29 of the CR, or if the CRs were shorter than 29 hours, from hour 5 to the end of CR. Melatonin amplitude was defined as the average value during hours 5-29 or from hour 5 to the end of the CR, whichever was shorter.
For data analysis, the timing of the light pulses were referenced using the midpoint of the timing of the light stimulus. Phase shifts produced by the intervention regimens were defined as the absolute difference between the phases obtained during CR1 and CR2. By convention, positive phase shifts represent phase advances and negative phase shifts represent phase delays. All group data are expressed as average ±SEM.
Urine volume, subjective alertness, and cognitive throughput data were first normalized for comparisons across subjects; then, average waveforms were generated across all subjects in each group. This analysis was performed because circadian phase and amplitude could not be easily determined for individual curves for these variables. Urine volume was expressed as volume of urinary excretion per minute. Subjective alertness and cognitive throughput scores were referenced to each subject's mean value and normalized in terms of the variability (z-score) found in hours 5-29 of CR1, or if CR1 was shorter than 29 hours, from hour 5 to the end of CR1. For each subject, each measure (CBT, melatonin, urine volume/min, alertness z-score, and throughput z-score) was averaged in 2-hour bins referenced to the melatonin phase for that individual during CR1 and/or during CR2. These 2-hour bins were then averaged across subjects and expressed relative to the center time of that bin.
Statistics were performed using SAS for Windows version 6.12 (SAS Institute, Cary, NC). Unpaired t tests were used for comparisons between older and younger subjects. Two-factor (CR number and time) repeated-measures analysis of variance was used for the average waveform measures.
Results
Baseline Circadian Phase Characteristics
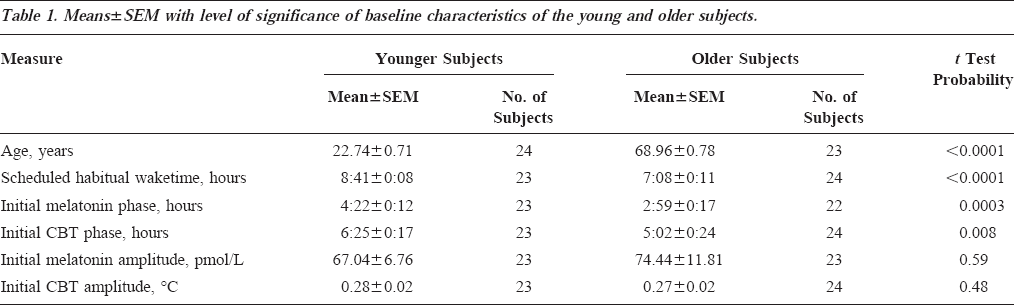
At baseline, the younger and older subjects differed in the timing of circadian and sleep parameters, but not in the amplitude of the circadian rhythms of melatonin and CBT (Table 1). The circadian phases of melatonin and of CBT as assessed during CR1 were located at a significantly earlier clock time in older subjects than in younger subjects. The timing of the baseline scheduled sleep-wake cycle differed between the two age groups such that the scheduled waketimes of older people occurred at a significantly earlier clock time. There was no statistically significant difference in the amplitude of either the melatonin rhythm or the CBT rhythm.
Means±SEM with level of significance of baseline characteristics of the young and older subjects.
Phase Delay Stimulus
During the phase delay stimulus, group A subjects were exposed to bright light during late night/early morning hours, approximately coincident with the middle and end of habitual sleep time. The protocol and melatonin data from one older subject who was exposed to this stimulus are shown in Figure 1. In this individual, during CR1 the phase of the melatonin rhythm occurred at 3:47 whereas after the exposure to bright light for 3 consecutive days, the melatonin phase occurred at 10:42, ie, the circadian rhythm was delayed by almost 7 hours. A total of seven older and seven younger subjects were exposed to this stimulus with light centered approximately 0.2 hours before the melatonin phase (0.18±0.24 hours earlier; range, 1.67 hours earlier to 1.93 hours later). Complete melatonin data were not available in one older subject; a phase delay was observed in the CBT data. The average melatonin phase shift observed in six older subjects was -6.83±0.49 hours (shift to a later hour) and -6.63±0.25 hours in seven younger subjects (Table 2). These phase shifts were not statistically significantly different between the two age groups (t test probability, 0.70) Given our sample sizes of six and seven and a common SD of ∼0.9, with an alpha value of 0.05 (two-sided) and a beta value of 0.80, we had the power to detect a difference of 1.7 hours. When the size of the phase delay was calculated using CBTmin as the marker of circadian rhythms, the average time of the CBTmin shifted -6.73±0.21 hours (range, -5.58 to -7.25 hours) in the older subjects, whereas the average phase shift observed for the CBT rhythm in the younger subjects was -6.91±0.49 hours with a range of -4.88 to -9.03 hours. A t test yielded no significant difference in the phase shifts observed in the two age groups (P=0.74).
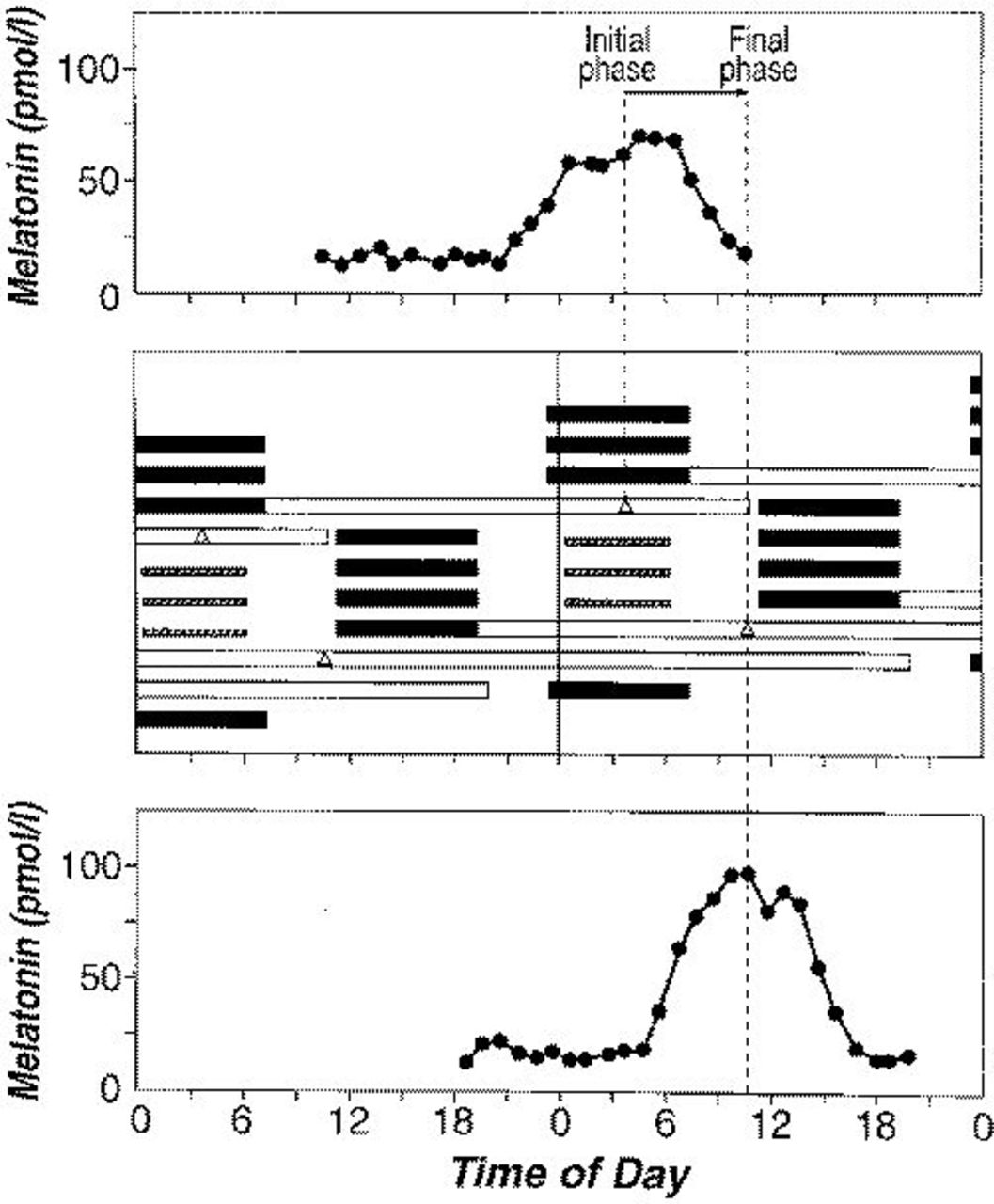
Plasma melatonin data from a single subject (subject 1229, a 73-year-old woman) during CR1 (top panel) and CR2 (bottom panel). The middle panel is a double-raster plot of the protocol. The data are double-plotted so that each day is plotted both next to and below the day before it. Dark bars indicate scheduled sleep times, open bars indicate CR times, and hatched bars indicate times of bright light exposure, whose midpoint occurred 1.47 hours before the CBT phase and 0.40 hours before the time of the melatonin phase (Δ). Time (in hours) is along the x *-axis, and days are along the y *-axis. Dotted vertical lines indicate the time of the melatonin phase during CR1 and CR2.
Changes in melatonin and CBT rhythms after exposure to bright light stimuli.
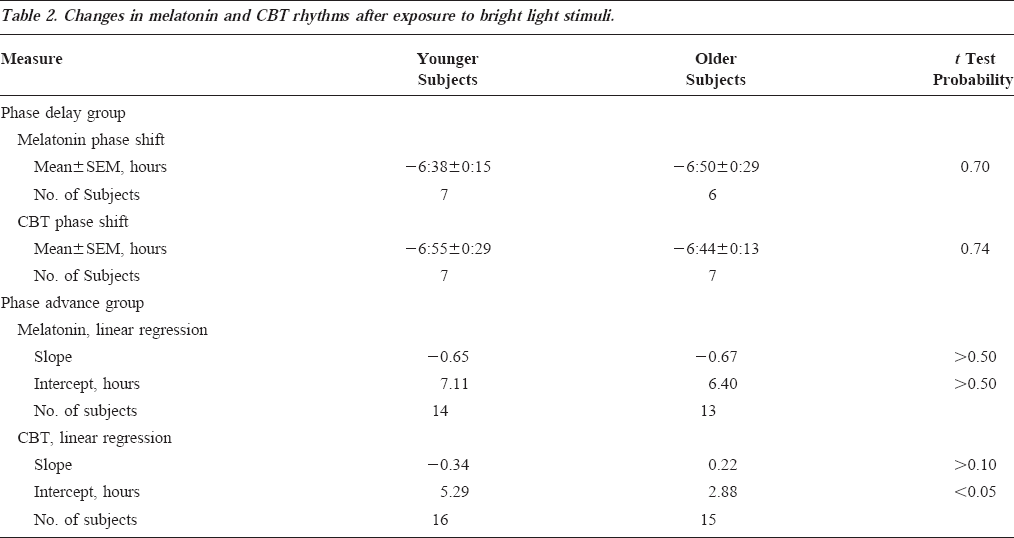
Phase Advance Stimulus
Seventeen older subjects and sixteen younger subjects in groups B and C were exposed to light centered 2.50-8.45 hours after the melatonin phase, corresponding to 5 hours of light beginning in the early/mid-morning hours. In one older and one younger subject, the amplitude of the melatonin rhythm was so low during the second CR that the phase could not be determined. In this older subject with a low amplitude melatonin rhythm during CR2, a 7.35-hour phase delay was observed for the CBT data. In the younger subject, a phase advance was observed in CBT data. In four older subjects, complete melatonin data were not available; only CBT data could be used to estimate the phase shift: in two of these subjects, a phase advance of CBTmin was observed; in one, a phase delay of CBTmin was observed; and in one of these subjects, the amplitude of the circadian CBT rhythm was reduced to such a low level that accurate determination of the phase could not be made. In one younger subject, complete melatonin data were not available; a phase advance was observed using CBTmin data.
Because of the range of light exposure times (2.50-8.45 hours), linear regression was performed on these data. Linear regression was performed separately on the size of the phase shift and the timing of the light pulse relative to the melatonin or CBT phase marker (Table 2). The slope of the lines fit by linear regression of the phase advance stimuli were not significantly different between younger and older subjects for CBT or for melatonin; the intercept was significantly higher in younger subjects as compared with the older subjects for phase shifts of the CBT rhythm but not for phase shifts of the melatonin rhythm (Table 2).
All Data
The individual phase shifts are plotted in Figure 2 together with the theoretical phase response curve (PRC) 53 derived from a recent mathematical model for the effects of light on the human circadian pacemaker. For both the phase delay and phase advance shifts, the observed data are close to the predicted phase shifts in both young and older people. However, there were more phase shifts that were smaller than the PRC predicted in older than in younger subjects (χ2 probability, 0.044) (Figure 2).
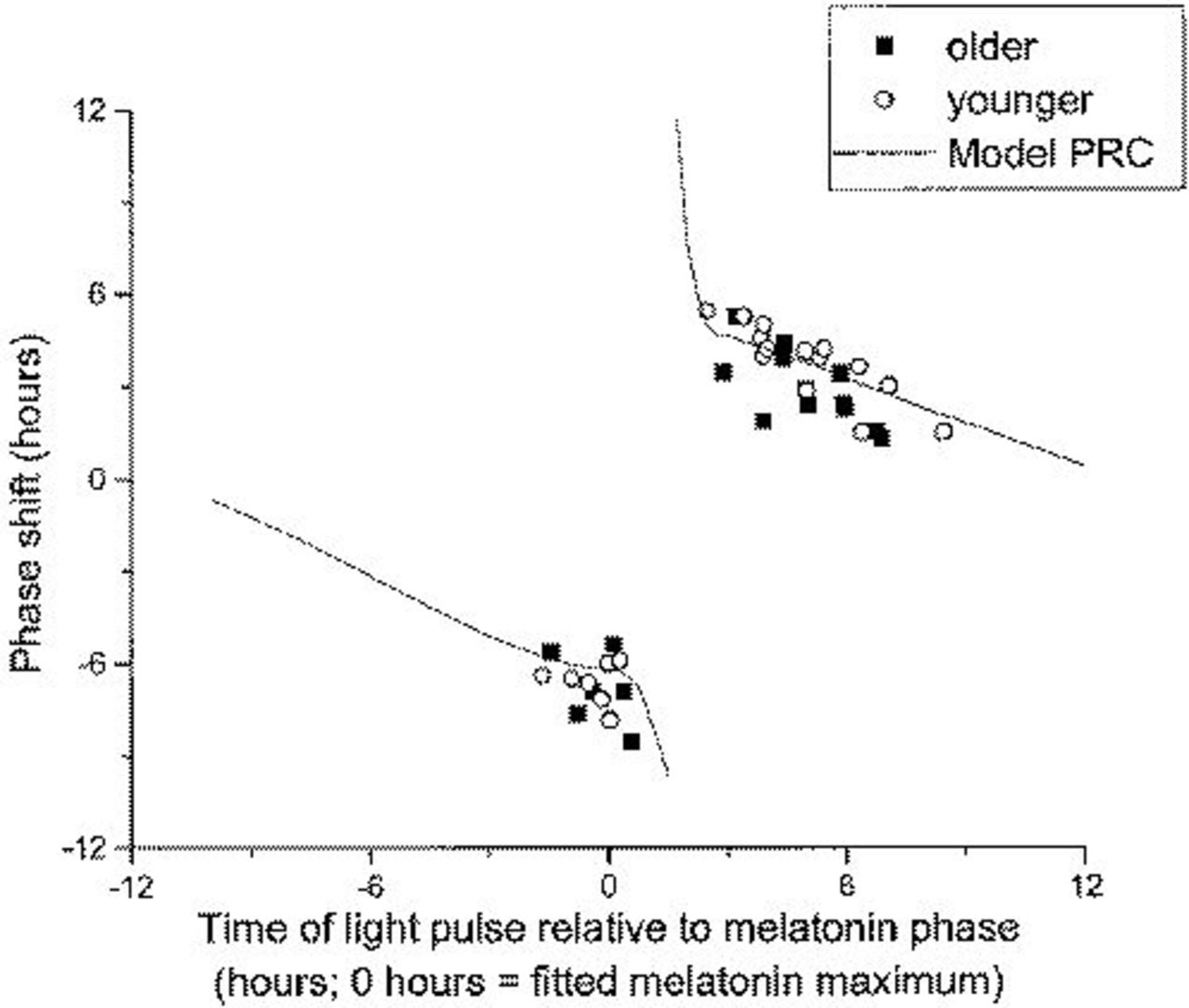
Plasma melatonin phase shifts in younger (○) and older (▪) subjects compared with results from a mathematical simulation of the protocol 53 (solid line). Time of the light pulse relative to the melatonin phase is plotted on the x *-axis, and the size of the phase shift is plotted on the y *-axis. Abbreviation: PRC, phase response curve.
The overall Pearson correlation between melatonin and CBT phase shifts in combined data from phase delay and phase advance protocols was 0.989 in older subjects (P=0.0001) and 0.979 (P=0.0001) in younger subjects (Figure 3).
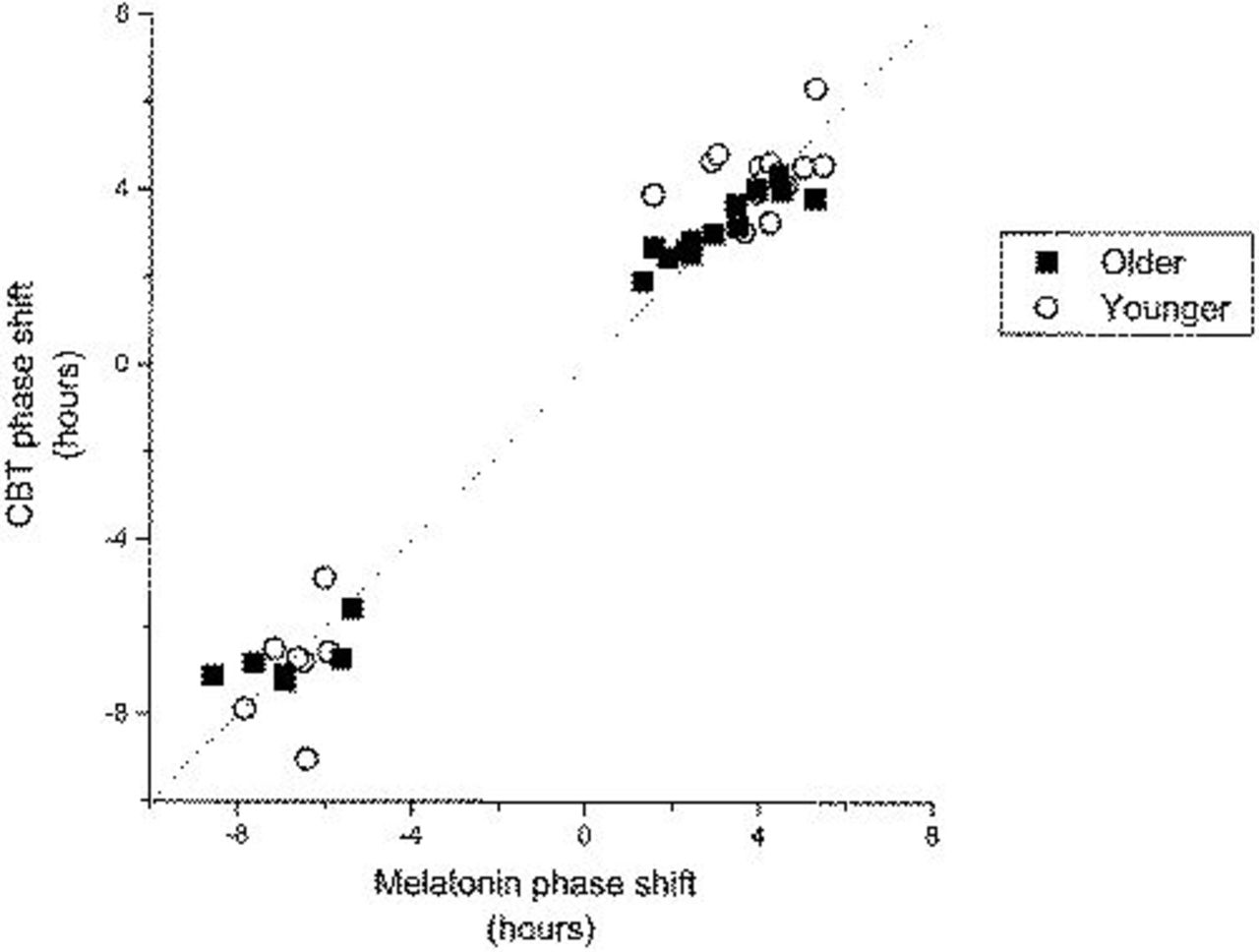
Comparison of the melatonin phase shifts (x *-axis) and CBT phase shifts (y *-axis) for each subject. Data from older subjects are represented by ▪; data from younger subjects are represented by ○. The dotted line represents equal phase shifts in melatonin and CBT.
To investigate whether the phase shifts observed for CBT and melatonin in the older people also occurred in other variables, the circadian waveforms of melatonin, CBT, urine volume, calculation throughput, and subjective alertness were computed for the CR before the phase shift (CR1) and the CR after the phase shift (CR2). All waveforms were aligned with respect to the melatonin phase during that CR (Figure 4; time 0 on the x-axis). The circadian waveforms of subjective alertness, calculation performance, urine volume, CBT, and melatonin and their phase relationship to the melatonin rhythm both overlap and were very similar during CR1 and CR2. Two-factor repeated-measures analysis of variance showed a significant (P < 0.01 in all measures) change with time within the CR but not a significant difference with time relative to the melatonin phase between CR1 and CR2 in any measure except calculation performance measures in the phase delay condition. This indicates that, on average, the size and direction of the phase shift was the same for all variables.
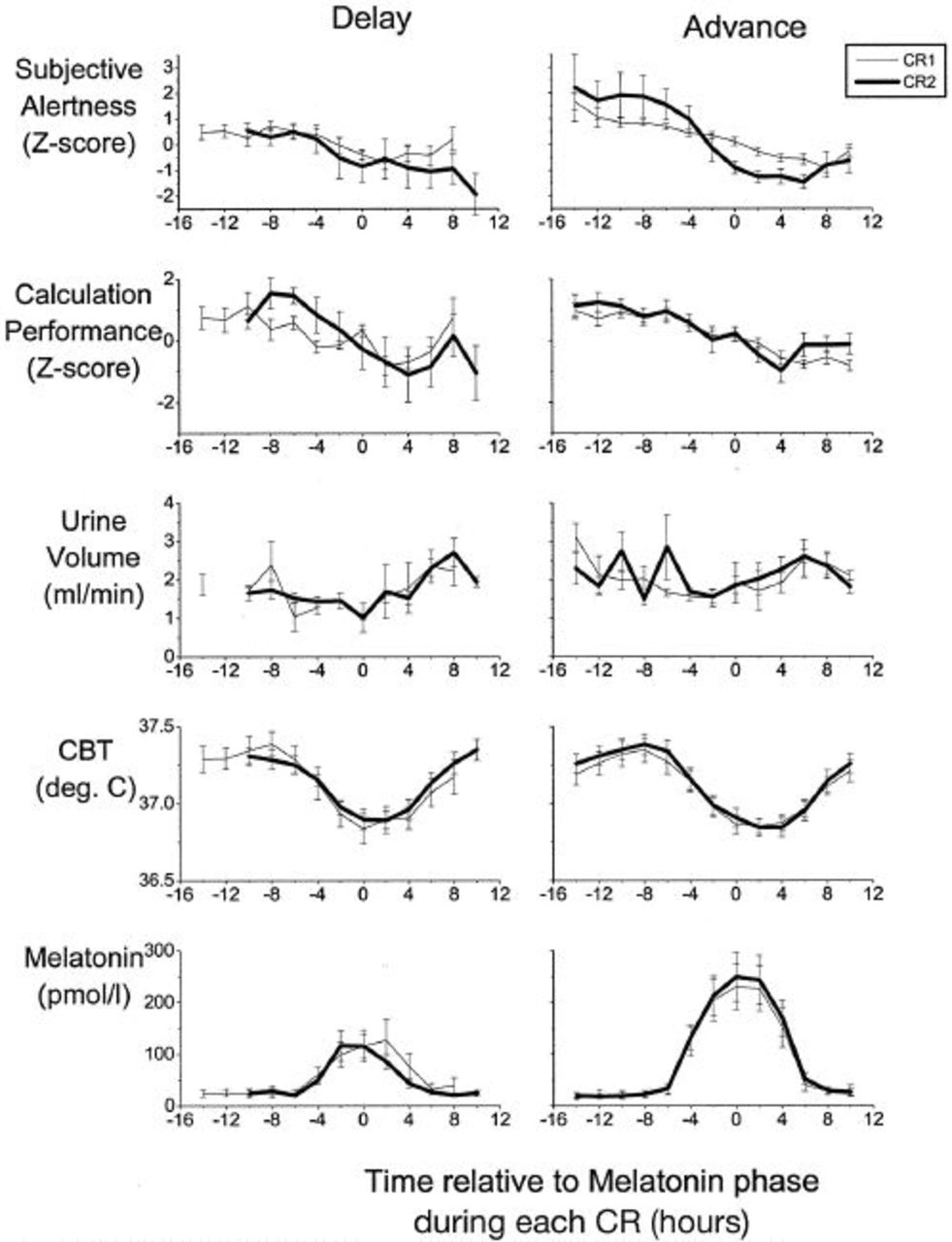
Average (±SEM) waveforms of plasma melatonin, CBT, urine volume, calculation throughput, and subjective alertness during the first CR (thin line) and second CR (heavy line) in the older men and women in the phase delay and phase advance groups. Data were compiled in 2-hour bins relative to the melatonin phase and are shown referenced to melatonin phase in each CR (time 0 on the x *-axis).
Discussion
The data demonstrate that the circadian pacemaker of older people, monitored by assessing multiple rhythms, can be shifted by scheduled light exposure and that the direction of the advance or delay shift is dependent on the phase of the circadian cycle at which the bright light exposure occurred. Therefore, the phase-resetting response of the pacemaker of older people to bright light is qualitatively similar to the response of the pacemaker of young people. However, quantitative changes in the response to light may occur with aging: the phase-resetting capacity in response to a bright light stimulus at some phases of the circadian cycle may be attenuated in healthy older men and women. Although older men and women had similar size phase shifts after the phase delay stimulus, they had smaller phase advances after morning light exposure, when compared with results from younger men, and smaller phase shifts than predicted from a theoretical PRC.
Although the difference in the response to phase advance stimuli in this study could be a result of gender differences in the two age groups, we did not observe significant differences in the light-induced phase shifts between the older men and women. However, additional subjects will be needed to further evaluate potential gender differences in the response to light stimuli of reduced strength in older people to better understand quantitative changes that may occur with aging.
Baseline Characteristics
The observed advanced phase of the temperature and melatonin rhythm as well as the earlier clock time of scheduled awakening are all in accordance with our previous reports1,54,55 and reports from others.3,4,25–27,56–59 This confirms that an advance of the phase of endogenous circadian rhythms is a salient characteristic of the aging process, even in very healthy older people. In contrast to the observed robust differences in circadian phase, we did not observe robust age-related changes in endogenous amplitude of either the melatonin or the temperature rhythm. Individuals in this report are a subset of those in our recent reports 23 on the absence of major changes in endogenous amplitude of the melatonin rhythm in healthy older people.
Phase Shifts in Younger and Older Persons
These data do not support the concept of a global reduction in phase-shifting capacity with advancing age. The lack of a difference between young and older subjects to a phase delay stimulus reduces the possibility that any gross change in the suprachiasmatic nuclei with aging60–64 is a cause of potential differences in phase-shifting capacity to stimuli that cause phase advances of the circadian system. Many pathophysiologic mechanisms could account for the differences in phase, amplitude, and phase-shifting speed observed in the rhythms of older subjects. However, given the differential changes with aging that seem to occur at only some circadian phases of bright light exposure, these simple explanations do not seem sufficient. Therefore, it remains unclear how any gross changes could explain both the earlier entrained phase of older subjects as well as the reduced capability to adjust to shifts in environmental cycles.
The differential response to phase advance but not phase delay stimuli with aging could be explained in several ways. First, this may indicate that in both younger and older subjects separate mechanisms, including separate neuroanatomical and/or neurotransmitter systems, may be responsible for phase advances and delays of the circadian system and that these systems are affected differentially by aging. Some evidence for the involvement of separate mechanisms in advances and delays has been obtained in some experimental situations (for review, see 65 ). One mechanism for the difference in phase advances but not phase delays between our younger and older subjects could be that phase advances usually involve transients, whereas phase delays are virtually complete within the next circadian cycle after administering the stimulus.66,67 Increased transients, or a slower change to final phase, could be a possible explanation for our results, as well as for those reports in the literature involving slower phase shifts in older subjects. However, the similar magnitude of the phase shifts in the melatonin, CBT, urine volume, subjective alertness, and cognitive throughput rhythms suggests that these transients occur at the level of the circadian pacemaker, and not just at the level of one single output of the pacemaker. If transients were present, they were in the same direction and the same magnitude and therefore indistinguishable in these experiments from an alteration of the central circadian pacemaker. Furthermore, our recent data obtained in young subjects do not provide evidence for marked transients in human circadian phase resetting measured in this manner. 68
Alternatively, a reduction in the magnitude of the phase advances but not the phase delays could be explained by assuming that the light stimulus used in the present experiment is so strong that it induces a saturating response in the phase delay region of both young and older subjects and that an age-related reduction in the sensitivity of the system was therefore not detected. If the light stimulus were near saturating strength in the phase advance region in the young subjects, an overall age-related reduction of the system to light would lead to submaximal advances in the older subjects. This scenario points to the importance of the assessment of dose-response curves in which not only the maximum response of the system to light is assessed but also its sensitivity. Changes in the retina 69 of older mammals have been observed. The age-related difference in the size of the phase shift we observed could therefore be a result of reduced light input to the retina or reduced sensitivity of the retina to light among older persons. 70 However, this would seem unlikely because the older subjects in our study had an ophthalmologic examination to verify the absence of eye pathology before entry into the study; mild cataracts were found in only one of the older subjects. Furthermore, we did not observe a global difference between younger and older subjects in the size of the phase shift, as one would expect if change in the anatomical structures involved in processing this information were the cause of the observed differences in the phase advance groups.
We and others have reported that the timing of circadian rhythms in older subjects is advanced when compared with data from younger adults. If that advanced timing were solely the result of age-related changes in the resetting sensitivity of the circadian system to light, we would expect an increased phase advance and/or decreased phase delay response to light in older people. Thus, our present finding of a reduced phase advance response and a similar phase delay response to light in older adults suggests that age-related changes in the phase response to light cannot explain the age-related advanced timing of sleep and circadian rhythms.
However, we have found that the phase relationship between the circadian timing system and the habitual sleep-wake cycle changes with age, such that the self-reported awakening in older subjects occurs at an earlier circadian phase.1,55 We have further found that this altered phase angle of entrainment with age is associated with a reduced ability to maintain sleep in the early morning hours as derived from self-reports 1 and polysomnographic sleep recordings. 71 Because during sleep the light input to the circadian pacemaker is reduced, both the response to light and the phase angle between the sleep-wake cycle and circadian rhythmicity contribute to the phase angle of entrainment. For a complete understanding of the age-related changes we have reported in entrainment and for understanding human entrainment under natural conditions, both factors (response to light and phase angle between sleep and circadian rhythmicity) need to be taken into account. Finally, for a more complete understanding of age-related changes in the sensitivity of the human circadian system to light stimuli, intensity-response experiments over a broad range of illuminances and across many circadian phases will be required.
Footnotes
Acknowledgments
We thank Dr Gordon H. Williams for continued support, David W. Rimmer and the laboratory technicians involved in data collection during the experiments, Dorothy Chen for the processing and organization of melatonin data, Dr Jamie Zeitzer for the melatonin analysis programs, Atesa Farshian for help with subjective alertness and cognitive throughput data, and Jason Sullivan and Drs Diane B. Boivin and Richard E. Kronauer for helpful comments on the manuscript. This work was supported in part by grants NIA-1-R01-AG06072 from the National Institute on Aging and NIMH-1-R01-MH45130 from the National Institute of Mental Health and by General Clinical Research Center grant NCRR-GCRC-M01-RR02635 from the National Center for Research Resources; E.B.K. is supported by fellowship 1-K01-AG00661-01 from the National Institute on Aging.