
Abstract
Background
Insulin-like growth factor 1 (IGF1) plays a critical role in estrogen-induced uterine proliferation, but it is unclear whether this estromedin function occurs in other estrogen-sensitive tissues such as the mammary gland. To elucidate this possibility, we investigated the cellular localization and hormonal regulation of mRNAs for IGF1 and 2, their cognate receptors (IGF1R, IGF2R), and IGF binding proteins 2-5 (BPs 2-5) in the rhesus monkey mammary gland.
Methods
Ovariectomized monkeys were treated with placebo, estradiol (E2), and E2 plus progesterone (E2/P4) for 3 days, after which mammary tissue was harvested for in situ hybridization and immunohistochemical analyses.
Results
IGF1 and IGF2 mRNA levels were significantly increased and BP2 mRNA decreased by E2 and by E2/P4 treatment. IGF1R mRNA was increased by combined E2/P treatment but not by E2 alone. BP5 mRNA was decreased by E2/P4. No differences in IGF2R, BP3, and BP4 mRNA levels were detected in any treatment group. Mammary IGF1 and IGF2 mRNA levels were both positively correlated with local epithelial proliferation, assessed by immunodetection of the proliferation-specific antigen, Ki67. IGF1 and IGF1R expression were negatively correlated with local programmed cell death, as assessed by the in situ TUNEL method. In contrast, BP2 expression was negatively correlated with epithelial proliferation and positively correlated with programmed cell death. IGF2R, BP3, BP4, and BP5 levels were not significantly correlated with either proliferation or death.
Conclusions
Thus, E2-induced proliferation is associated with upregulation of both IGF1 and IGF2 expression and downregulation of BP2 expression. These data suggest that the local mammary IGF system is involved in sex steroid-induced mammary epithelial cell hyperplasia.
Introduction
Insulin-like growth factors (IGFs) have been implicated in normal mammary gland development (mainly in rodents1,2) and in breast cancer.3–6 Both IGF1 and IGF2 have been detected in breast cancer specimens,3–6 and IGF2, but not IGF1, is highly expressed by breast cancer cell lines.5,7 The actions of both IGFs are mediated by the membrane-bound IGF1 receptor, which is a tyrosine kinase with high homology to the insulin receptor. 8 This receptor is also expressed by mammary tumor cells, and its overexpression has been linked with progression of malignant characteristics.4,7 The IGFs also bind with high affinity to a group of proteins present in the circulation and in tissues, termed the IGF binding proteins (IGFBPs 9 ). One of these proteins, IGFBP3, is abundant in mammary tissue and cell lines and has been associated with breast cancer.6,10
Although it seems clear that there is extensive IGF system expression in normal and cancerous mammary tissue, the exact cellular localization of each of the IGF system components and their sensitivity to estrogen and progestin regulation in vivo remains uncertain. 11 It is difficult to investigate the regulation and potential actions of the IGF system in cell lines, where cumulative genetic mutations have altered normal regulatory mechanisms. Furthermore, important interactions between stromal and epithelial compartments cannot be elucidated in cell lines. There are also significant limitations to the study of human biopsy specimens: for example, sampling error and inability to control the hormonal milieu. Therefore, in the present study, we have used ovariectomized rhesus monkeys to investigate the cellular localization and hormonal regulation of the mammary IGF system in a model with special relevance to humans.
Materials and Methods
Female rhesus monkeys (Macaca mulatta) 6-13 years of age from the National Institutes of Health Poolesville colony were used in accordance with a protocol approved by the National Institute of Child Health and Human Development Animal Care and Use Committee. Ovariectomies were performed under ketamine anesthesia via a midventral laparotomy in the late follicular phases of their menstrual cycles. Three weeks after surgery, animals were randomly assigned to groups, each group receiving placebo (n=7) or hormone-containing pellets inserted subcutaneously between their shoulder blades under ketamine anesthesia. The E2 group (n=5) received 3-day sustained release 17β estradiol pellets (5 mg). The E2/P4 group (n=5) was treated with both 17β estradiol (5 mg) and progesterone (10 mg) 3-day release pellets. Mammary tissue was removed, snap-frozen on dry ice, and stored at -70°C. Serial sections of 10-μm thickness were cut at -15°C and thaw-mounted onto poly-L-lysine-coated slides for histochemical analysis. Serum hormones were measured by radioimmunoassay 3 days after pellet implantation, at the time mammary tissue was collected for analysis.
The IGF1, IGF2, IGF1 and IGF2 receptor, and IGFBP clones used for riboprobe synthesis have been described previously.12,13 The in situ hybridization protocol has also been described previously in detail.13,14 Parallel sections were hybridized to sense probes and exposed together with the various antisense hybridized sections. The quantitation of hybridization signal was carried out in a blinded fashion. Tissue from both mammary glands was sectioned en bloc, so that tissue from both glands was present on each slide. The hybridization signal overlying cell groups was captured by a blinded observer at x400 using a monochrome video camera, and the results were analyzed with National Institutes of Health image v1.57 software. Background was determined from sense probe hybridized sections, and this value was subtracted from the raw data before further analysis. Values for nine to 10 areas in each of two to three sections per animal were meaned to obtain individual values. These values were statistically compared among groups using analysis of variance. Significant differences among means were determined by Fischer's least significant difference test.
Immunohistochemical detection of the proliferation-specific Ki67 antigen was performed by the avidin-biotin-immunoperoxidase technique. The mouse antihuman Ki67 serum (Boehringer Mannheim, Indianapolis, Ind) was used at a dilution of 1:50. Fresh-frozen tissue sections were fixed in 4% formalin-phosphate-buffered saline (PBS) for 30 minutes. After blocking in 1% hydrogen peroxide/10% methanol PBS, avidin-biotin blocking reagent, and 10% normal sheep serum, tissue sections were incubated overnight at 4°C with mouse antihuman Ki67 antibody or with 1% PBS as the control. Thereafter, tissue sections were treated with biotinylated sheep antimouse IgG 1:40 for 30 minutes at room temperature, followed by a 45-minute incubation with the avidin-biotin-peroxidase complex (Vectastain ABC Elite Peroxidase Kit, Vector Laboratories, Burlingame, Calif). The antigen-antibody complex was visualized by incubation with freshly prepared 3,3'-diaminobenzidine (3,3’ diaminobenzidine Substrate Kit, Vector Laboratories), and the tissue was counterstained with methyl green. The ApopTag® kit was used for in situ apoptosis detection as described by the manufacturer (Oncor, Gaithersburg, Md). Fresh-frozen tissue sections were fixed in cold acetone and cold 2:1 ethanol:acetate at -20°C for 10 and 5 minutes, respectively. After blocking with 1% bovine serum albumin PBS, tissue sections were incubated for 1 hour at 37°C with digoxigenin-dUTP and TdT or with 1% PBS as the control. Tissue sections were then incubated in antidigoxigenin-peroxidase for 30 minutes at room temperature. The antigen-antibody complex was visualized with 3,3′-diaminobenzidine and stained with methyl green as described above. Rat dam mammary glands 4 days after weaning were used as positive controls for the ApopTag® method.
Ki67 and ApopTag® positive cells were counted by a blinded observer at x400 magnification using a Leitz Laborlux microscope (Leitz, Rockleigh, NJ). Several hundred nuclei per lobule were scored for nine to 10 lobules in each animal. Data are means±SEM representing the ratio of positive-staining nuclei to total number of counted nuclei. Mean values for each group were compared by analysis of variance followed by the least significant difference test. Correlations were analyzed using the StatView statistical analysis software (SAS Institute, Cary, NC). A simple regression analysis was independently applied to each IGF system component's relation to tissue apoptosis and pro-liferation.
Results
Localization of IGF System Components in Mammary Gland
IGF1 mRNA were localized in stromal cells adjacent to the glandular epithelium, whereas IGF2 mRNA was concentrated in both epithelial and associated stromal cells of the monkey mammary gland (Figure 1). IGF2 mRNA was also detected in vascular components of the mammary gland. IGF1R was concentrated in the glandular and ductal epithelium, whereas IGF2R mRNA was present in both epithelial and stromal cells (Figure 2) Figure 3. shows IGF binding proteins mRNA expression in the mammary gland. IGFBPs 2 and 5 mRNAs were homogeneously expressed in glandular and ductal epithelium. In contrast, IGFBP3 mRNA was heterogeneously distributed. It was focally concentrated in isolated epithelial cells throughout most lobules and was more extensively expressed in a subpopulation of lobules. IGFBP3 mRNA was also localized in vascular endothelium (Figure 3C and D). BP4 mRNA was very abundant in stromal cells and was detected at relatively low levels in the glandular epithelium (Figure 3E and F).
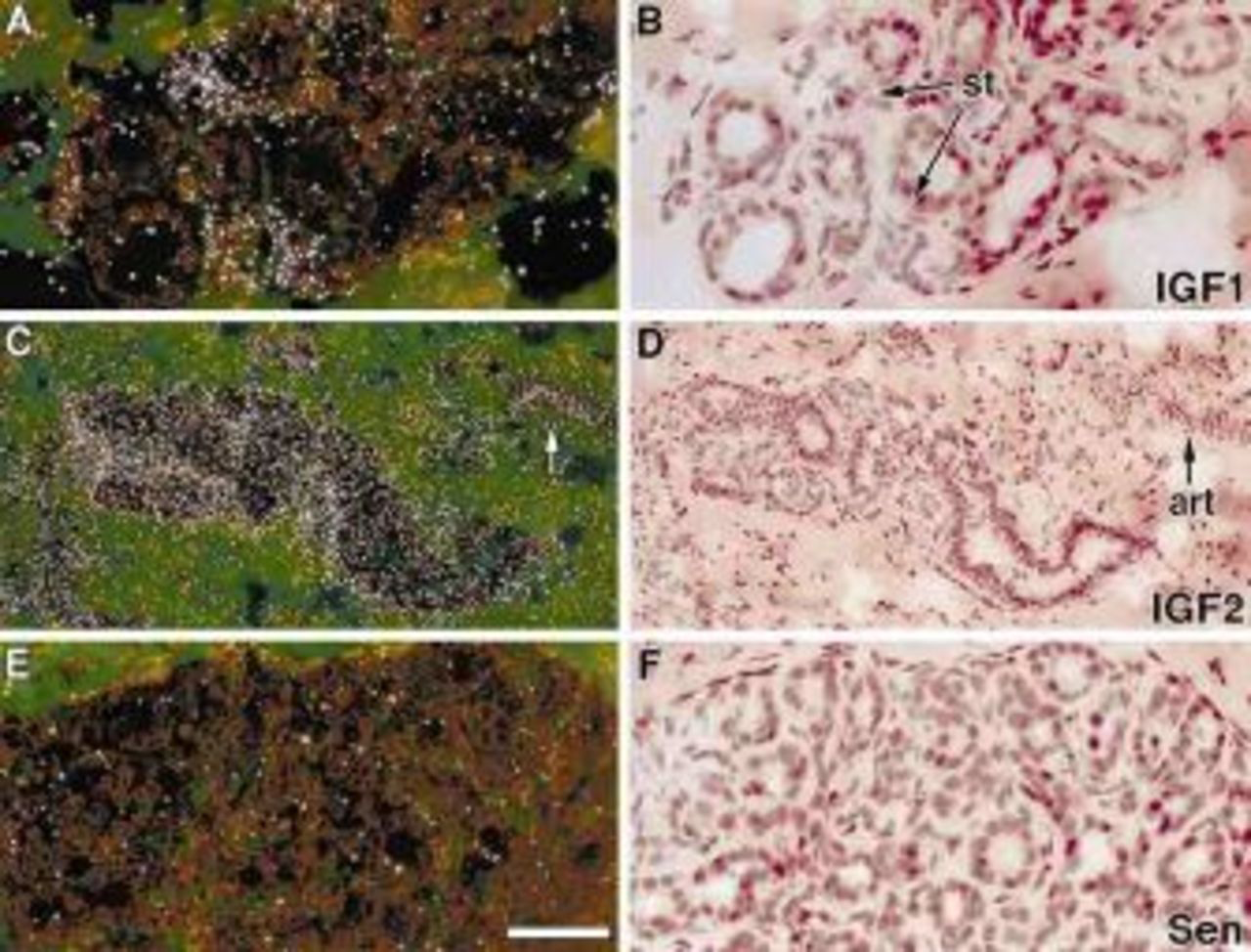
IGF1 and IGF2 mRNA localization in rhesus monkey mammary glands. These are representative sections from E2-treated animals. The hybridization signal appears as white grains in the dark field illumination. IGF1 mRNA was localized in stromal cells (st) (A and B), whereas IGF2 mRNA was found in both glandular epithelium and stroma cells (C and D). It was also concentrated in the blood vessels (art). Panels E and F show nonspecific signal derived from IGF1 sense probe hybridization (Sen). Bar=50 μm.
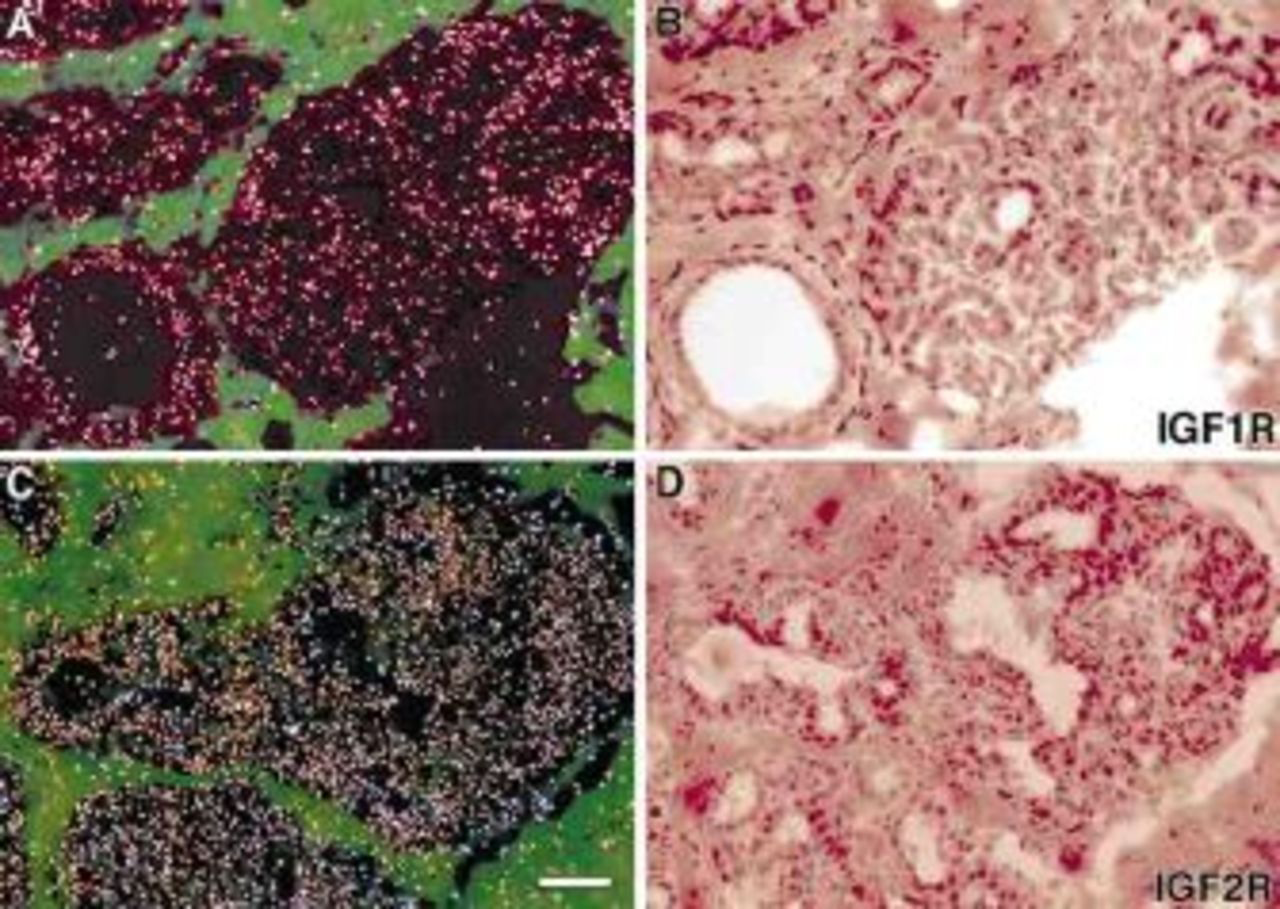
IGF1 and IGF2 receptor mRNA localization in rhesus monkey mammary glands. These are representative sections from E2-treated animals. IGF1R mRNA was concentrated in the glandular epithelium (A and B), whereas IGF2R mRNA was present in both glandular epithelium and stroma cells (C and D). Bar=100 μm.
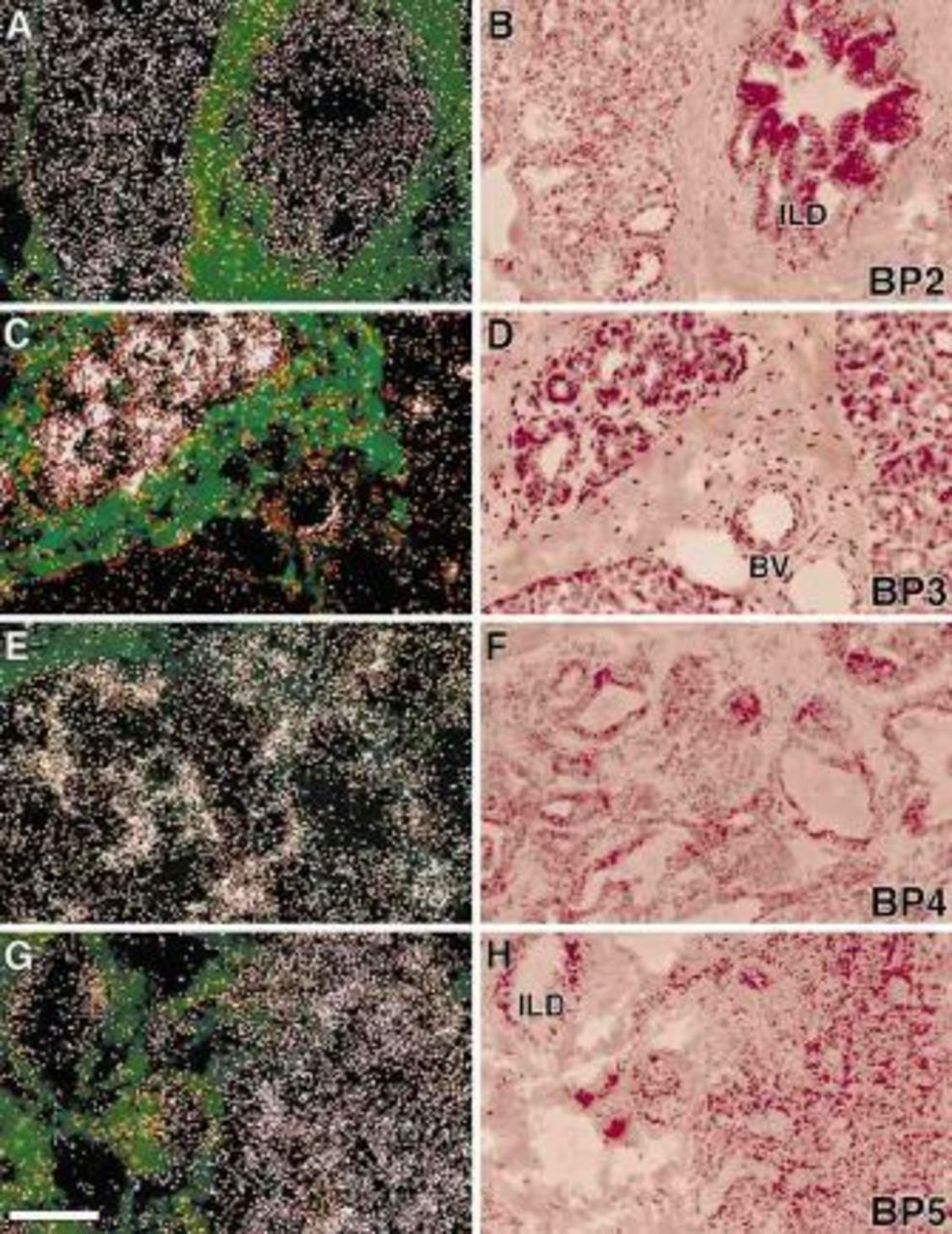
IGFBP mRNA localization in rhesus monkey mammary glands. BP2 mRNA (A and B) was homogeneously expressed in glandular, and ductal epithelium; BP3 mRNA (C and D) was heterogeneously distributed in mammary epithelial cells and was also localized in the endothelium of mammary blood vessel (BV). BP4 mRNA (E and F) was very abundant in stromal cells and was relatively lower in the glandular epithelium. BP5 mRNA (G and F) was detected in glandular and ductal epithelium and also in some stromal cells. Abbreviation: ILD, intralobular duct. Bar = 50 μm.
Effects of Gonadal Steroids on IGF System Expression
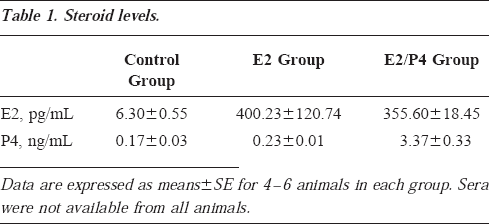
The effects of E2 and E2/P4 treatments on the IGF system mRNA levels are shown in Figure 4. Both IGF1 and IGF2 mRNAs were significantly increased by E2 and E2/P4 treatment. IGF1 mRNA was increased by approximately 40% in both the E2 and E2/P4 groups. IGF2 mRNA was increased by approximately 20% in both groups. IGF1R and BP5 mRNA were not altered by E2 treatment alone, but IGF1R was increased by approximately 50% (P<0.01), and BP5 was reduced by 25% (P<0.05) after E2/P4 treatment. BP2 mRNA was significantly decreased by both E2 and E2/P4 treatment. No differences in IGF2R, BP3, and BP4 mRNA expression were detected in any treatment group. Steroid levels in the different treatment groups are reported in Table 1. The E2 levels in the treatment groups approximated late follicular-periovulatory estradiol levels in humans and monkeys, and the P4 levels were characteristic of early luteal phase.
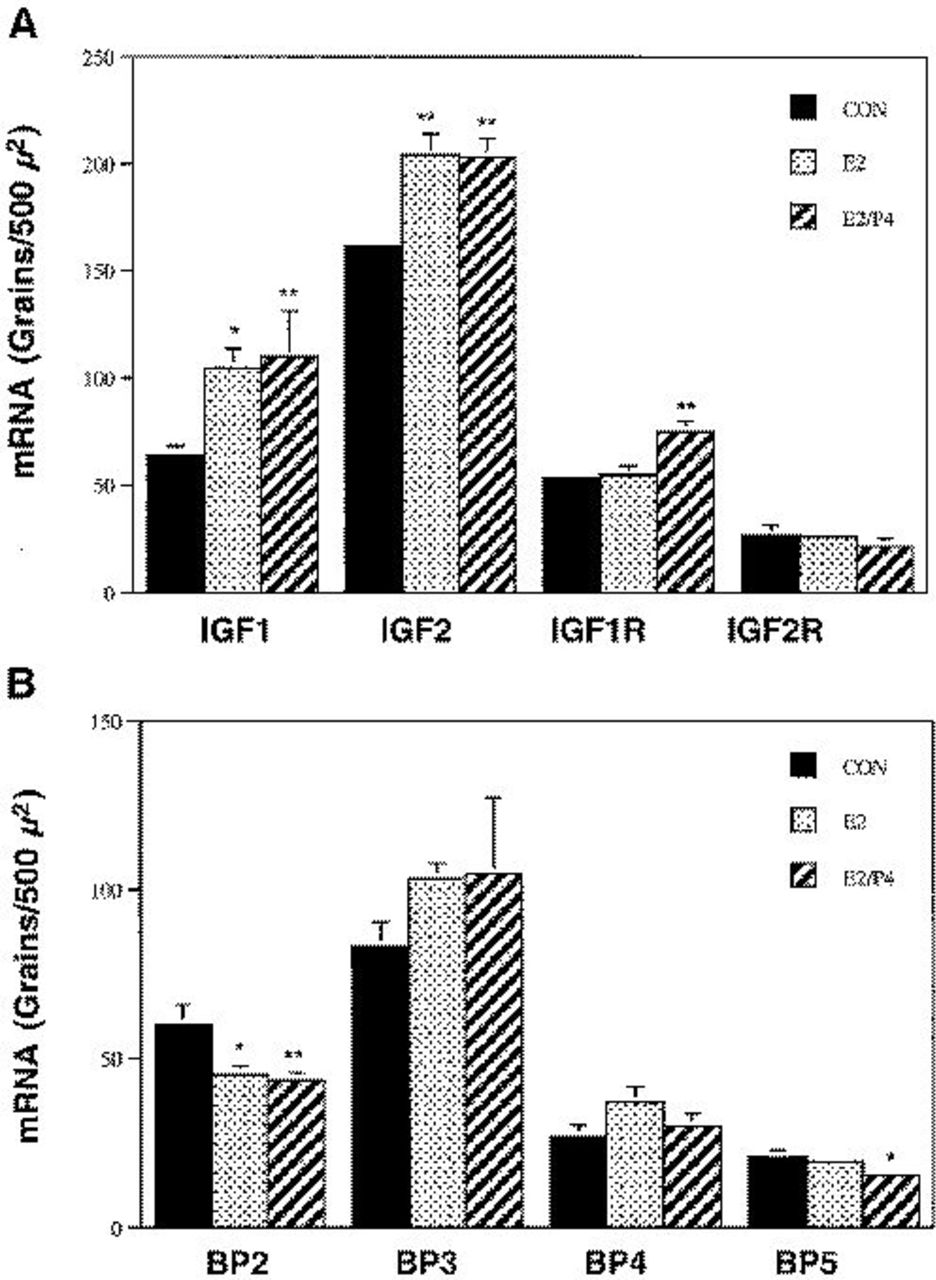
Effect of E2 and E2/P4 on IGF system mRNA levels in the primate mammary gland. Data represent means±SEM for 4-6 animals per group.
Steroid levels.
Correlations between IGF System mRNA Concentrations and Local Mammary Epithelial Proliferation and Apoptosis
Mammary epithelial proliferation and apoptosis were evaluated in tissue sections using immunodetection of the proliferation-specific Ki67 antigen 15 and the TUNEL method for detecting DNA fragmentation, respectively (Figure 5). Correlative analyses comparing serum levels of E2, mammary Ki67 and apoptosis, and mammary expression of different IGF system components are shown in Table 2. IGF1 and IGF2 mRNA levels were both positively correlated with local epithelial proliferation, whereas IGF1 and IGF1R expression were negatively correlated with programmed cell death in individual mammary glands. In contrast, BP2 mRNA concentration was negatively correlated with epithelial proliferation and positively correlated with programmed cell death. No correlation between IGF2R, BP3, BP4, and BP5 and these parameters of mammary epithelial turnover were found.
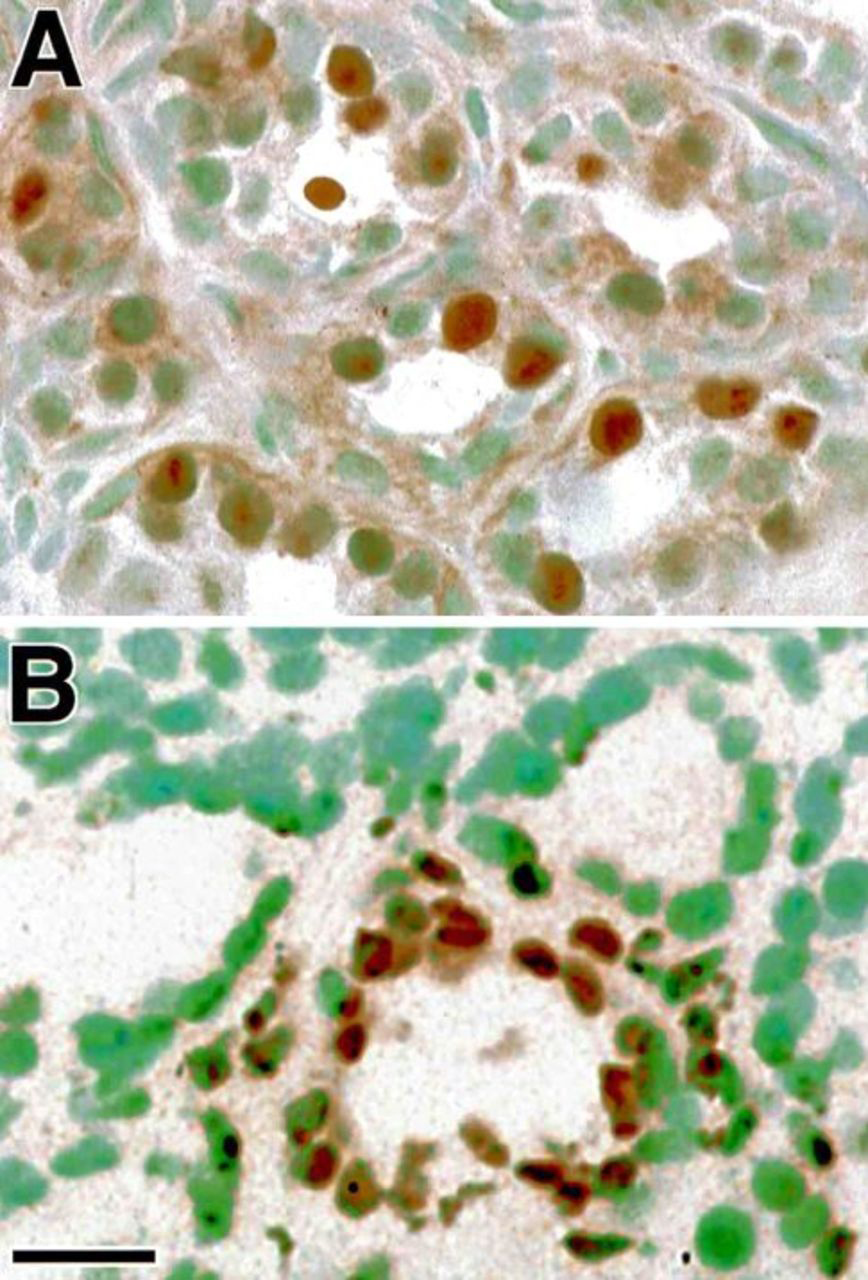
Mammary epithelial proliferation shown by Ki67 immunoreactivity (A) and programmed cell death detected by the TUNEL method (B) in ovariectomized monkeys treated with estradiol E2. Ki67 positive nuclei have a typical appearance with the brown immunostain concentrated in subnuclear vesicles (A). The TUNEL-positive apoptotic nuclei are shrunken or pycnotic in appearance, and some fragmented nuclei are stained. Bar=25 μm.
Correlations between IGF system components, sex steroid levels, proliferation, and apoptosis in the primate mammary gland.
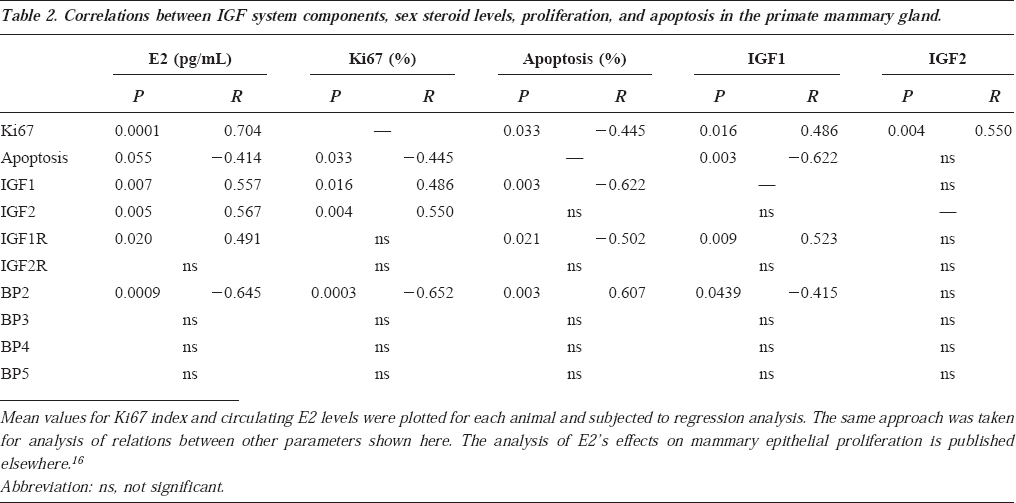
Serum E2 was positively correlated with mammary epithelial proliferation and IGF1 and IGF2 mRNA levels and negatively correlated with apoptosis and BP2 mRNA levels. IGF1 and BP2 mRNA levels were negatively correlated (Table 2). The mean group values for proliferation (Ki67 index) were 7.8±2.6% for control, 48.2±3.6% for E2-treated, and 49.5±6.2% for E2/P-treated (five to seven animals per group; P<0.0001 for both sex steroid groups vs control). The mean group values for apoptotic index were 34.8±6.6% for control, 27.3±4.4% for E2-treated, and 9.2±3.1% for E2/P-treated (five animals per group; P=0.003 for E2/P vs control and P=0.02 for E2/P vs E2).
Discussion
There has been intense interest in the role of the IGF system in breast cancer (reviewed in4,5). IGF1R activation by IGF1 or IGF2 enhances the proliferation of breast cancer cell lines. 15 Pharmacological elevation of circulating IGF1 levels in the rhesus monkey stimulates marked increases in mammary epithelial hyperplasia, 17 and IGF1 levels within the high normal range are associated with increased breast cancer risk in premenopausal women. 18 Many components of the IGF system are detected in breast cancer tissue, suggesting that local autocrine/paracrine IGF effects in addition to endocrine stimulation by circulating IGFs may be involved in the promotion of breast cancer risk.
The present study demonstrates, for the first time, the normal cellular patterns of gene expression for all components of the IGF system in the primate mammary gland and the effect of estrogen alone and in combination with progesterone on their expression. The finding that IGF1 is localized in stromal cells adjacent to the mammary epithelium is consistent with in situ hybridization findings in the human, 3 and the fact that IGF1 is generally not detectable in breast cancer cell lines, which are epithelial in origin.5,7 The finding that IGF2 mRNA is found in mammary epithelium is consistent with the abundant IGF2 expression by epithelial breast cancer cell lines.4,5 It is not clear whether previous studies have investigated IGF2 gene expression in normal mammary tissue, and there is some variability in the detection of IGF2 mRNA in breast cancer. For example, Giani et al 19 report IGF2 mRNA localization in stromal fibroblasts surrounding malignant epithelium but not in the tumor epithelium. Other groups have reported IGF2 mRNA detection in tumor cells as well as in adjacent stroma.20,21 The present study provides clear, unequivocal evidence for IGF2 expression in normal mammary epithelium, as well as in normal stromal cells and vascular components of the mammary gland.
IGF1 receptor mRNA was concentrated in mammary alveolar and ductal epithelium, whereas IGF1 was expressed by the adjacent stromal cells. This constellation suggests that IGF1 exerts paracrine effects on the epithelial IGF1R. Interestingly, this pattern of IGF system expression parallels that seen in the primate uterine endometrium, where IGF2 and the IGF1R receptor are concentrated in the epithelium and IGF1 is localized in stromal cells adjacent to the epithelium (13,22 and our unpublished data). This is an interesting parallelism, because in the uterus, stromal IGF1 is clearly upregulated by estrogen and is critically involved in mediating estrogen's proliferative effects on the uterine epithelium, 23 suggesting that similar stromal-epithelial interactions may operate in the mammary gland. The role of the IG2R, also termed the type II IGF receptor and mannose-6-phosphate receptor, in IGF signaling is unclear. 24 This receptor may be primarily involved in metabolism of IGF2 and mannose-6-phosphate enzymes unrelated to the IGF system in the mammary gland.
Although IGF1 and IGF2 mRNAs are both significantly increased by estrogen in the primate mammary gland, the IGF1R is not increased by estrogen alone. IGF1R mRNA is, however, increased by combined E2/P4, suggesting that progesterone stimulates its expression. These findings with respect to IGF1&2 are consistent with information from breast cancer cell lines, in which their expression is increased by estrogen. 7 Some cell lines, however, also upregulate IGF1R expression in response to estrogen treatment, 25 whereas we did not find an estrogen-induced increase in mammary epithelial IGF1R mRNA in vivo. This applies to the present study in the mammary epithelium, and to our previous investigations of sex steroids’ effects on IGF system expression in the uterus.13,22 Thus, estrogen's proliferative effects on normal mammary epithelium may be promoted by increased ligand availability rather than by increased IGF1R expression. The finding in the present study that IGF1R mRNA was substantially and significantly increased by the addition of progesterone to E2 treatment suggests that progesterone enhances IGF1R expression in the mammary gland, whereas it does not in the uterus.13,22 This is an interesting observation, which may help explain why progesterone is not protective of the mammary epithelium as it is for the uterine epithelium, because amplification of IGF1R expression is associated with enhanced tumorigenesis in many different model systems.
The mechanisms whereby estrogen stimulates epithelial proliferation are unclear. IGF1 is critically involved in this process in the murine uterus, as shown by the fact that mitosis is arrested in the G2 phase of the estrogen-induced mitotic cycle in the uterine epithelium of the Igf1 targeted gene deletion mouse. 23 Whether IGF1 and IGF2 have identical or distinct roles in promoting estrogenic effects is unclear. Both these homologous peptides interact with the IGF1R to stimulate mitosis, so it is unclear how they could have distinct effects. IGF1 and IGF1R mRNA levels are actually more closely correlated with reduced mammary epithelial apoptosis than with proliferation, whereas IGF2 mRNA level is tightly correlated with proliferation. Increased IGF2 gene dosage is associated with breast cancer in humans and mice,26,27 supporting the significance of the correlation between IGF2 expression and mammary epithelial proliferation. Transgenic IGF1 overexpression in the murine mammary gland has been associated with inhibition of the normal involution process and reduced apoptosis after lactation,28,29 suggesting that IGF1 may indeed have an antiapoptotic role in breast biology. However, a recent study of human mammary tissue based on biopsy specimens revealed an increase in Ki67 index associated with oral contraceptive use, which was also associated with increases in tissue IGF1 mRNA levels. 30
One possible explanation for apparent differential effects by IGF1 and IGF2 could be differential interactions with one or more of the high-affinity binding proteins, which are thought to modulate IGF action. This is the first study to characterize the cellular expression patterns and hormonal regulation of all the major IGFBPs in the primate mammary gland. Preliminary studies revealed very little BP1 or 6 expression, whereas BPs 2-5 were highly abundant, so these were studied more comprehensively. BP2 was the only IGFBP that was responsive to sex steroid regulation, being significantly decreased by both E2 alone and E2/P4. BP2 mRNA levels were, moreover, positively correlated with mammary epithelial apoptosis and negatively with proliferation. The IGF binding protein most intensively studied with respect to breast cancer is BP3.6,10,31 The present study is, to our knowledge, the first analysis of the cellular localization of BP3 mRNA in normal mammary tissue. We have found that BP3 mRNA is intensely and focally expressed by mammary epithelial cells in a heterogeneous pattern throughout the lobular structure. This is in distinct contrast to other components of the IGF system, which are homogeneously distributed throughout the mammary gland. BP3 mRNA does not seem to be regulated by E2 or P4, and its local expression is not correlated with mammary epithelial cell proliferation or death. These data suggest that mammary BP3 is not directly related to the IGF system or to mammary cell turnover. There is indeed experimental evidence supporting IGF-independent effects for BP3 in mammary epithelial cells. 29
IGF BP 4 and 5 expression has not heretofore been mapped in the primate mammary gland, to our knowledge. This study demonstrates that BP4 is heavily expressed in mammary stroma and, to a lesser extent, in epithelium. BP5 is highly expressed in glandular and ductal epithelial cells. Neither BP4 nor 5 seemed to be altered in the E2 or E2/P4 treatment groups, suggesting that these mammogenic steroids do not regulate their expression. Moreover, BP4 and BP5 levels were not significantly correlated with mammary epithelial proliferation or apoptosis.
In summary, this work has shown that mRNAs encoding IGF1, IGF2, their cognate receptors, and IGF BPs 2-5 are each expressed in some abundance in the primate mammary gland. IGF1 was expressed by stromal cells immediately adjacent to the mammary epithelium, whereas IGF2 was expressed by the epithelial cells themselves, in addition to stromal elements. The IGF1R was concentrated in the epithelium, suggesting a paracrine mode of action for IGF1 and an autocrine effect for IGF2 on this IGF signal-transducing receptor. E2 increased both IGF1 and 2 expression, whereas combined E2/P4 significantly augmented IGF1R expression. In contrast to its positive effects on the ligands and IGF1R, E2 downregulated BP2 expression in the mammary gland. Unfortunately, the present study, limited to mRNA evaluation, did not provide important information about growth factor peptide or binding protein levels or posttranslational processing, which may have critical effects on IGF bioactivity. However, the regulation of these IGF system mRNAs by sex steroids and correlation of their local expression with mammary epithelial proliferation and apoptosis strongly suggests that locally-produced IGFs are involved in mediating the sex steroids’ actions in the mammary gland.