
Abstract
Objective
Although many studies have documented the tumor-promoting role of hepsin in several types of malignancies, little is known about its clinical and biological significance in breast cancer.
Materials and Methods
Hepsin expression was examined in 4 pairs of fresh breast tumor samples and corresponding nontumor breast tissues by Western blotting. Immunohistochemistry for hepsin was performed on an additional cohort of 215 archival breast cancer samples. The clinical significance of hepsin expression was analyzed. Knockdown of hepsin expression was performed in 2 breast cancer cell lines, MDA-MB-231 and HER18, with a high abundance of endogenous hepsin, and the effects of hepsin silencing on cell invasion and proliferation were evaluated.
Results
Hepsin was aberrantly overexpressed in breast cancer tissues relative to adjacent nontumor tissues. Its overexpression was significantly associated with tumor stage (P = 0.037), lymph node metastasis (P = 0.010), estrogen receptor positivity (P = 0.019), and progesterone receptor positivity (P < 0.0001) in patients with breast cancer. Down-regulation of hepsin expression by small interfering RNA (siRNA) significantly reduced cell proliferation and invasion in both the MDA-MB-231 and HER18 cells compared to nonspecific control small interference RNA.
Conclusion
Our data demonstrate that hepsin expression is frequently up-regulated in breast cancer tissues, which is associated with tumor growth and progression. Thus, inhibition of hepsin expression might be of therapeutic significance.
Metastasis is the most common cause of death in patients with breast cancer, which consists of multiple steps including local tumor invasion followed by dissemination to and re-establishment at distant sites.1–3 Extracellular protease-mediated degradation of neighboring extracellular matrix components is critical for tumor invasion and metastasis, allowing desquamation of tumor cells into the surrounding environment and invasion of basement membranes in target metastatic organs.4–7 A broad range of proteases has been identified to be involved in the proteolysis of the extracellular matrix, which can been classified into 4 families: (a) serine proteases, (b) metalloproteases, (c) aspartic proteases, and (d) cysteine proteases. 6
Hespin is a type II transmembrane serine protease that was originally cloned from complementary DNA libraries of human liver and hepatoma cells. 8 Its amino acid sequence is similar to that of other serine proteases and contains His, Asp, and Ser residues at position 203, 257, and 353, respectively, as the catalytic triad for enzymatic activity. 9 Several studies have shown that hepsin is up-regulated in 90% of prostate tumors, with levels often increased more than 10-fold, and exclusively expressed in tumor cells. 10 Moreover, it has demonstrated that hepsin levels increase with prostate cancer progression and reach maximum levels in more advanced (Gleason grade 4/5) prostate carcinomas.11,12 The study, in a mouse model of prostate cancer by Klezovitch et al., 7 has revealed a critical role for hepsin in disruption of the basement membrane and promotion of adenocarcinoma and metastasis. Tanimoto et al. 6 suggested that hepsin is frequently overexpressed in ovarian tumors. Antibodies neutralizing hepsin protease activity do not impact cell growth but inhibit invasion of prostate and ovarian tumor cells in culture. 13 Hepsin overexpression predicts poor prognosis in renal cell carcinoma.14,15 These findings suggest an important role for hepsin in tumor progression and metastasis.
Despite the extensive studies of hepsin in numerous malignancies, relatively little is known about its clinical and biological significance in breast cancer. In the present study, we examined hepsin expression in breast cancer cell lines and tumor samples from a large random breast cancer cohort. To evaluate its biological function, we knocked down the expression of endogenous hepsin in 2 breast cancer cell lines, MDA-MB-231 and HER18, using small interfering RNA (siRNA) and assessed the effects on tumor cell proliferation and invasion.
Materials and Methods
Patients and Tumor Characteristics
Four cases of fresh breast cancer samples coupled with adjacent nontumor breast tissues were obtained from the First Affiliated Hospital of China Medical University (Shenyang, China). Tissue samples were processed for detection of hepsin by Western blot analysis as described below. For immunohistochemical studies, tumor samples from resection specimens were collected from a consecutive cohort of 215 patients with breast ductal carcinoma who underwent modified radical mastectomy for the disease at the First Affiliated Hospital of China Medical University between 2007 and 2009. Patients with preoperative neoadjuvant therapy or with evidence of other malignancies were excluded from the study. The tissue samples had been routinely fixed in 10% neutral formalin, embedded in paraffin, and sectioned to 4 μm in thickness. Conventional hematoxylin and eosin staining was performed to confirm the diagnosis and tumor type. Estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER-2) status was determined and scored on immunohistochemistry as previously described.16,17 Staging followed the guidelines of the Cancer Staging Manual of the American Joint Committee on Cancer. 18 Of invasive carcinomas, 62% were associated with nodal disease. Of the 215 patients with breast cancer, 156 (73%) were positive with ER, 151 (70%) with PR, and 50 (23%) with HER-2 overexpression, as seen on immunohistochemistry. At the time of diagnosis, 24%, 59%, and 17% of the patients had tumors at TNM stages I, II, and III, respectively. The study protocol was approved by the Ethics Committee of China Medical University (Shenyang, China), and written informed consent was obtained from each patient.
Cell Culture and Transfection
Four human breast cancer cell lines, MDA-MB-231, MDA-MB-435, MDA-MB-453, and MCF7-HER18 (HER18), as well as one normal human breast tissue cell line, MCF-10A, were purchased from the Institute of Cellular Research, Chinese Academy of Science (Shanghai, China). They were maintained in Dulbecco modified Eagle medium with 10% fetal bovine serum (Gibco BRL, Carpinteria, CA), 100-U/mL penicillin, and 100-μg/mL streptomycin in a 5% carbon dioxide (CO2) incubator at 37°C. Cells were seeded at a density of 1 × 105 cells per well in 6-well plates and, at 70% to 80% confluence, were transfected with SMARTpool hepsin siRNAs (50 nmol/L; Dharmacon, Lafayette, CO) using the cationic lipid DharmaFECT 1 reagent (Dharmacon) according to the manufacturer's instructions. After transfection, the hepsin messenger RNA (mRNA) level was assessed 48 hours later by real-time polymerase chain reaction (PCR), and the protein level was examined 72 hours later by Western blotting. For control experiments, siCONTROL Non-Targeting siRNA Pool (Dharmacon) was used under the same conditions.
Proliferation Assay
Cell proliferation was analyzed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. 19 MDA-MB-231 and HER18 cells transfected with hepsin-specific siRNAs or control siRNAs were separately seeded at a density of 1.5 × 103 cells per well in 96-well microplates and harvested daily for up to 5 days. The number of viable cells was determined using Cell Titer 96 Non-Radioactive Cell Proliferation Assay (Promega, Madison, WI) according to the manufacturer's protocol. The reduced 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide product was dissolved in 2% dimethyl sulfoxide, and the absorbance was measured at 570 nm. Each time point was done in sextuplicate wells, and the experiment was repeated 3 times.
Colony-Forming Assay
Detailed experimental procedures have been described previously. 20 Briefly, MDA-MB-231 and HER18 cells transfected with hepsin-specific siRNAs or control siRNAs were plated in 6-well plates at a density of 1000 cells per well. After a 14-day incubation, cells were washed with PBS, fixed in 10% methanol for 15 minutes, and stained in Giemsa for 20 minutes. Colonies that consisted of more than 50 cells were scored. The colony formation ratio was calculated according to the following formula: colony formation ratio (%) = (colony number/seeded cells number) × 100%. Each experiment was repeated at least 3 times.
Immunohistochemistry
Paraffin sections (4 μm thick) were deparaffinized with xylene, rehydrated, and heated for 10 minutes in a steamer containing 10-mmol/L sodium citrate (pH 6.0) to retrieve antigen. Endogenous peroxidase was quenched with 3% hydrogen peroxide for 10 minutes. Sections were incubated with rabbit polyclonal antihepsin (sc-18572; Santa Cruz Biotechnology, Santa Cruz, CA; 1:50) for 1 hour, followed by the secondary reaction with DAKO Envision+ Reagent (DakoCytomation, Carpinteria, CA). Negative controls were included by omitting the primary antibody, and a known positive control was included with each batch. The stained sections were independently assessed by 2 pathologists without prior knowledge of the clinical data. Immunoreactivity was scored according to the percentage of immunoreactive cells over the total tumor cells counted as 0 if less than 10% cells were stained, 1 if 10% to 30% cells were immunoreactive, 2 if 30% to 70% cells were immunoreactive, and 3 if greater than 70% cells were immunoreactive. All tumor specimens were classified into 2 groups, namely, Hepsin-H (scored 0 or 1) and Hepsin-L (scored 2 or 3).
Antibodies and Western Blot Analysis
The same amount of protein samples (50 μg) was loaded, separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and transferred onto polyvinylidene difluoride membrane. After blocking, the membrane was incubated with primary antibody antihepsin (1:200) or anti-β-actin (Santa Cruz Biotechnology; 1:500) overnight at 4°C, followed by incubation with appropriate secondary antibody for 1 hour. Blots were developed using an enhanced chemiluminescence kit from Santa Cruz Biotechnology. The intensities of immunoreactive bands were measured by computerized image analysis (Quantity One software, Bio-Rad, Hercules, CA) and normalized to β-actin levels.
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
Total RNA was isolated using TRIzol (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The complementary DNA was reverse-transcribed from 2 μg of total RNA by SuperScript first-strand synthesis system for RT-PCR (Invitrogen). Real-time PCR was performed on a 7900 HT Fast Real-Time PCR System (Applied Biosystems, Foster City, CA) using SYBR Green PCR Master Mix (Life Technologies Corporation, Foster City, CA). Primer sequences used in this study included hepsin forward: 5′-TGTCCCGATGGCGAGTGTTT-3′, hepsin reverse: 5′-CCTGTTGGCCATAGTACTGC-3′, and β-actin forward: 5′-GAGCGGGAAATCGTGCGTGACATT-3′, β-actin reverse: 5′-GATGGAGTTGAAGGTAGTTTCGTG-3′. All samples were run in triplicate. Gene expression was normalized to the β-actin transcripts, and the relative mRNA expression between the nontumor and tumor samples were calculated using the 2-(ΔΔcycle threshold) method. 21
In vitro Matrigel Invasion Assay
In vitro Matrigel invasion assay was performed by using a 24-well millicell inserts (BD Biosciences, San Jose, CA) with polycarbonate filters (pore size, 8 μm). The upper side of polycarbonate filter was coated with Matrigel (50 μg/mL; BD Biosciences). The chambers were incubated at 37°C with 5% CO2 for 2 hours to allow the matrix to form a continuous thin layer. Then the MDA-MB-231 and HER18 cells transfected with hepsin siRNAs or control siRNAs (4 × 105 cells in 200 μL of 0.1% bovine serum albumin) were placed in the upper chamber. The lower chamber was filled with 10% serum-containing medium (700 μL). After incubating for 22 hours at 37°C in 5% CO2, cells on the upper surface of the filter were removed using a cotton swab. Cells invading through the Matrigel and filter to the lower surface were fixed with 4% neutral-buffered formalin and stained in 0.01% crystal violet solution. The cell numbers in 5 random fields at a magnification of ×200 were counted for each chamber, and the average value was calculated. Each assay was repeated 3 times.
Statistical Analysis
Statistical analyses were carried out using the SPSS.11 software (SPSS Inc., Chicago, IL). If not otherwise stated, means of at least 3 independent experiments ± SD are shown. The difference among means was analyzed by one-way analysis of variance followed by the Tukey test. The relationship between hepsin protein expression and clinicopathologic features of breast cancer was analyzed using the Fisher exact test for 2 categorical variables and the χ2 test for more than 2 variables.
Results
Hepsin Expression in Breast Cancer
Overexpression of hepsin has been reported in many cancers. Its expression status in breast cancer, however, remains unclear. To investigate the expression of hepsin in breast cancer, Western blotting analysis was done in paired breast cancer and noncancerous tissues adjacent to cancer lesions. Hepsin was found to be overexpressed at the protein level in all 4 examined breast cancer samples compared to adjacent nontumor tissues (Fig. 1A). To further examine the frequency and the localization of hepsin protein expression, we carried out an immunohistochemical analysis on a large cohort of tumor samples. Hepsin showed diffuse cytoplasmic staining pattern in breast cancer cells. The hepsin immunoreactivity was heterogeneous. In some cases, the invasive cells were negative for hepsin, whereas in others (Fig. 1B), almost all invasive cells stained positive (Fig. 1C). A more intense expression of hepsin was frequently detected at the invasive edge of the tumor areas (Fig. 1D). In contrast, very weak or no staining was found in surrounding nonneoplastic breast tissues. In addition, 143 (66.5%) of the 215 studied cases showed high expression of hepsin, defined as more than 30% cells stained positive. These results indicate an enhanced expression of hepsin in breast cancer tissues.
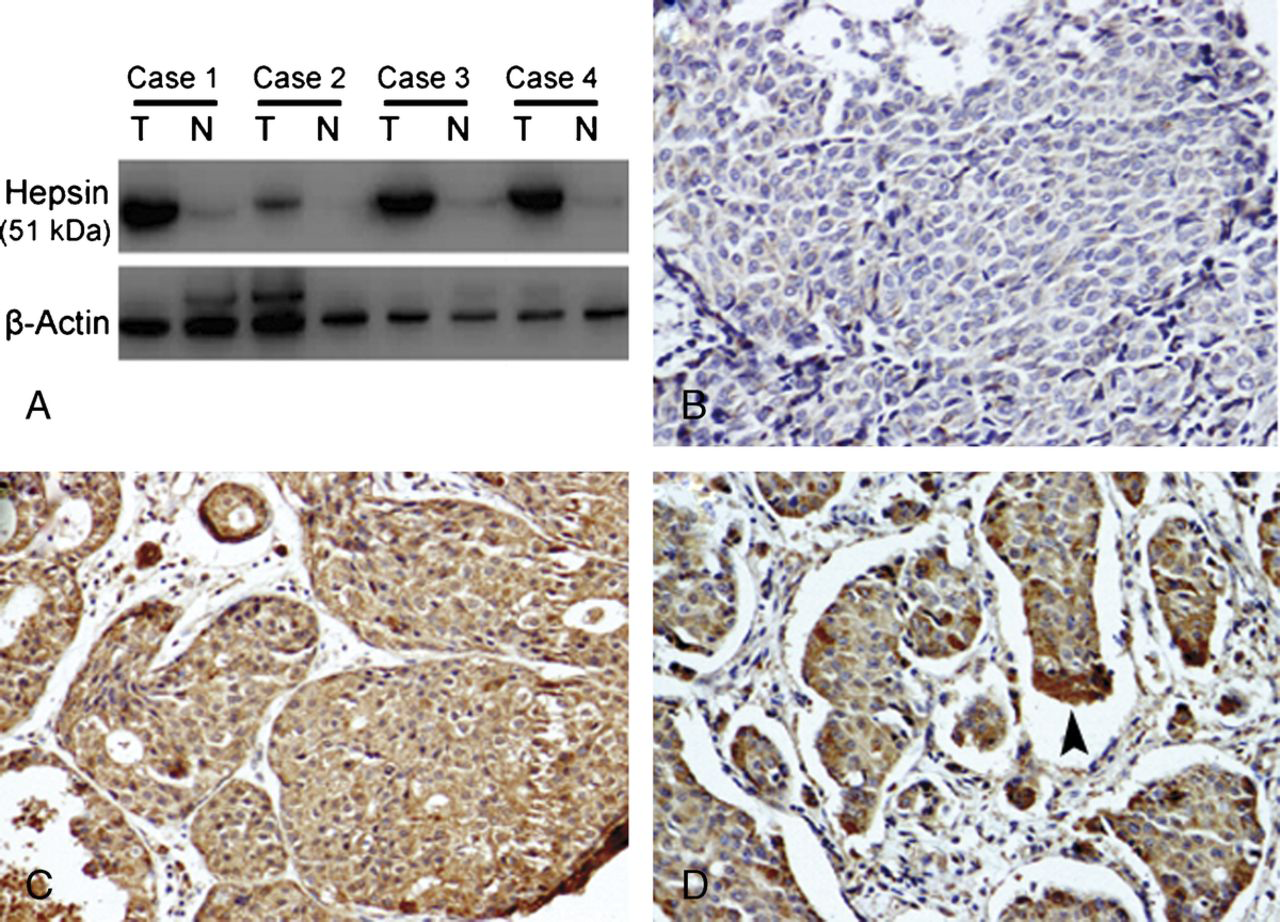
Expression of hepsin in breast cancer. A, Western blot analysis of hepsin expression in 4 cases of fresh breast tumor (T) and adjacent nontumor (N) breast tissues. Representative blots of 3 independent experiments are shown. The size of the hepsin band is approximately 51 kd. β-Actin was used as loading control. B-D, Representative breast cancer tissue sections stained with an antibody to hepsin (original magnification ×200). Hepsin shows diffuse cytoplasmic staining pattern. The hepsin immunoreactivity was heterogeneous: in some cases, the invasive cells were negative for hepsin (B), whereas in others, almost all invasive cells stained positive (C). Arrowhead in (D) indicates a more intense expression of hepsin in the invasive edge. In contrast, very weak or no staining was found in surrounding nonneoplastic breast tissues.
Clinical Significance of Hepsin Overexpression in Breast Cancer
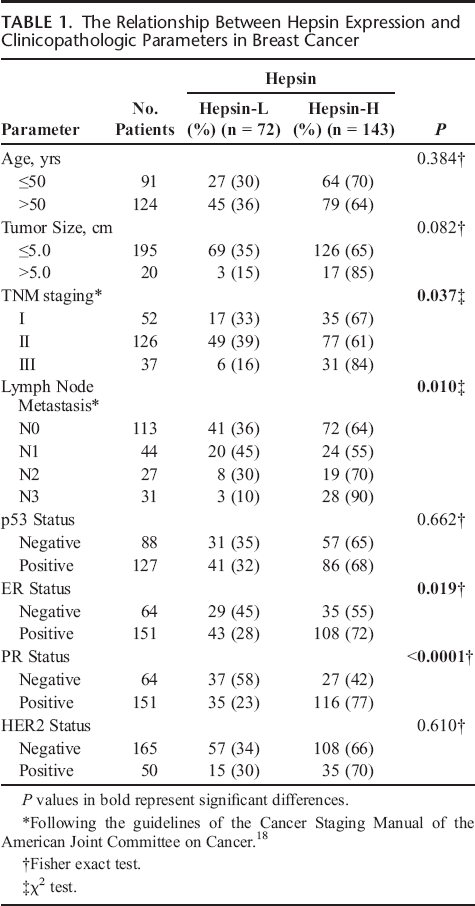
Next, we investigated the association of hepsin overexpression with clinical characteristics of patients with breast cancer. As shown in Table 1, hepsin overexpression was significantly correlated with tumor stage (P = 0.037), lymph node metastasis (P = 0.010), ER positivity (P = 0.019), and PR positivity (P < 0.0001) (Table 1). However, there was no significant correlation between hepsin expression and the age of the patients, tumor stage, HER2 status, and p53 status.
The Relationship Between Hepsin Expression and Clinicopathologic Parameters in Breast Cancer
Role of Hepsin in Breast Cancer Cell Invasion and Proliferation
We examined hepsin mRNA and protein expression levels in several human breast cell lines by real-time PCR and Western blot analysis, respectively. Notably, hepsin expression, either at mRNA or protein level, was abundant in 2 cancer cell lines, MDA-MB-231 and HER18 cells, whereas the other 2 cancer cell lines, MDA-MB-435 and MDA-MB-453, as well as the nontumor cell line MCF-10A had markedly lower levels of hepsin (Fig. 2, A and B). To explore the biological significance of hepsin in breast cancer, we specifically knocked down its expression in MDA-MB-231 and HER18 cells that were chosen owing to their high abundance of hepsin. The transfection of both the cell lines with hepsin-specific siRNAs caused a 60% to 80% reduction in hepsin transcripts compared to control siRNA-transfected corresponding cells (Fig. 2, C and D).
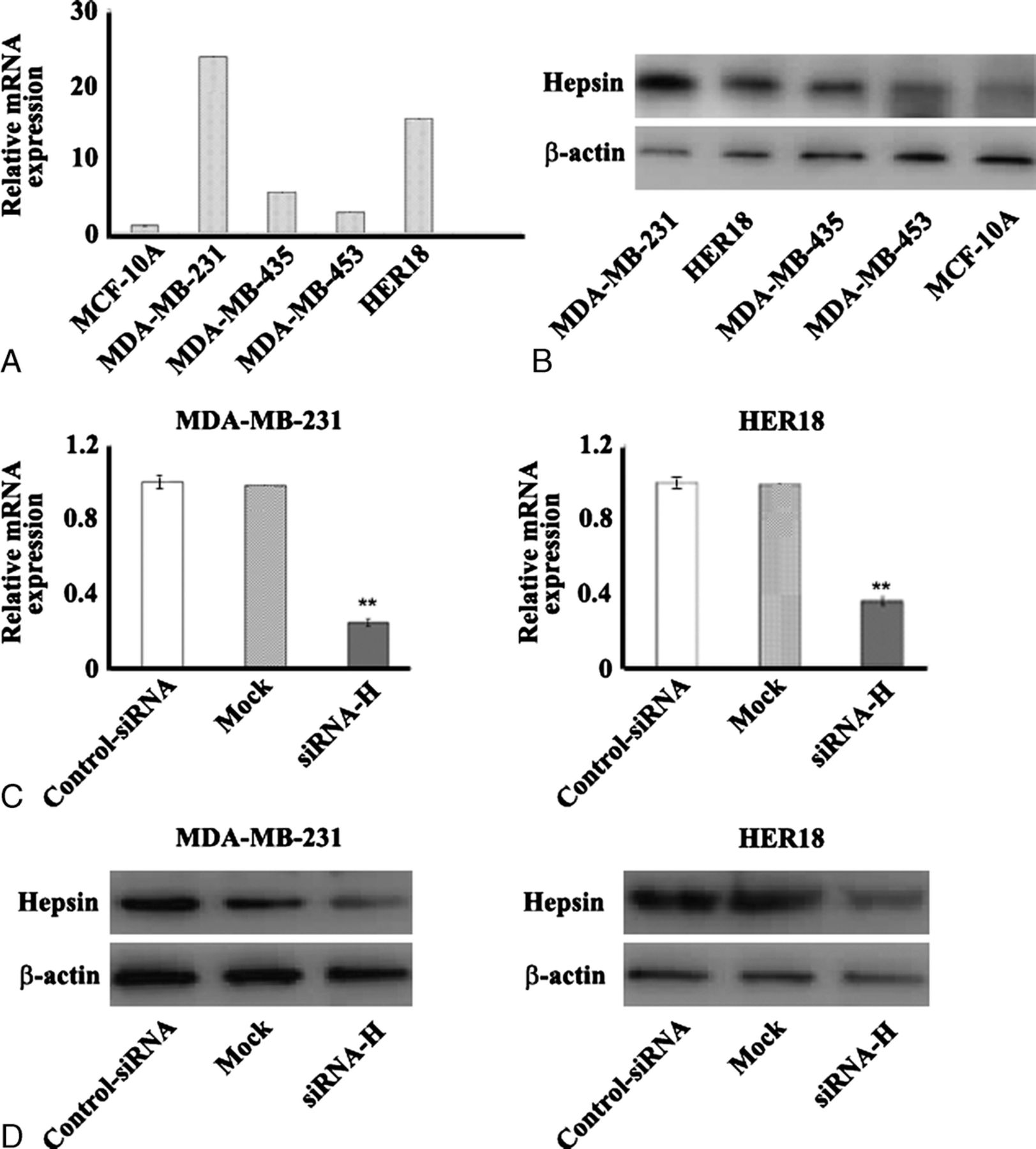
Targeted reduction of hepsin expression in breast cancer cell lines using siRNA. Quantitative real-time PCR (A) and Western blot (B) analysis of endogenous hepsin expression in several breast cancer cell lines (MDA-MB-231, MDA-MB-435, MDA-MB-453, and HER18) and one nontransformed human breast epithelial cell line, MCF-10A. C and D, Evaluation of the knockdown efficiency of hepsin-specific siRNAs (siRNA-H). After the transfection of the tested siRNAs, the hepsin mRNA level was assessed 48 hours later by real-time PCR (C), and the protein level was examined 72 hours later by Western blotting (D). Representative blots of 3 independent experiments are shown. The data shown in (C) are the mean ± SD of 3 independent experiments. The relative mRNA expression was given as a ratio with respect to the mRNA expression of hepsin in untreated cells (mock) that was taken as 1.0. β-Actin was used as an internal control. **P < 0.01 compared to scrambled control siRNA-transfected cells.
We next assessed the biological function of hepsin in cancer cell invasion and proliferation. Silencing of hepsin in MDA-MB-231 and HER18 significantly compromised their invasion capacities compared to control siRNA-transfected counterparts in a reconstituted basement membrane invasion chamber assay (P < 0.01; Fig. 3). Targeted reduction of hepsin resulted in a significant loss of viability (25%-40% reduction) in MDA-MB-231 and HER18 cells after 5-day culture, as assessed by the MTT assay (Fig. 4A). Moreover, the colony formation assay revealed that hepsin downregulation in the 2 breast cancer cell lines significantly decreased the colony formation ratio by approximately 40% (P < 0.05 relative to control siRNA-transfected cells; Fig. 4B).
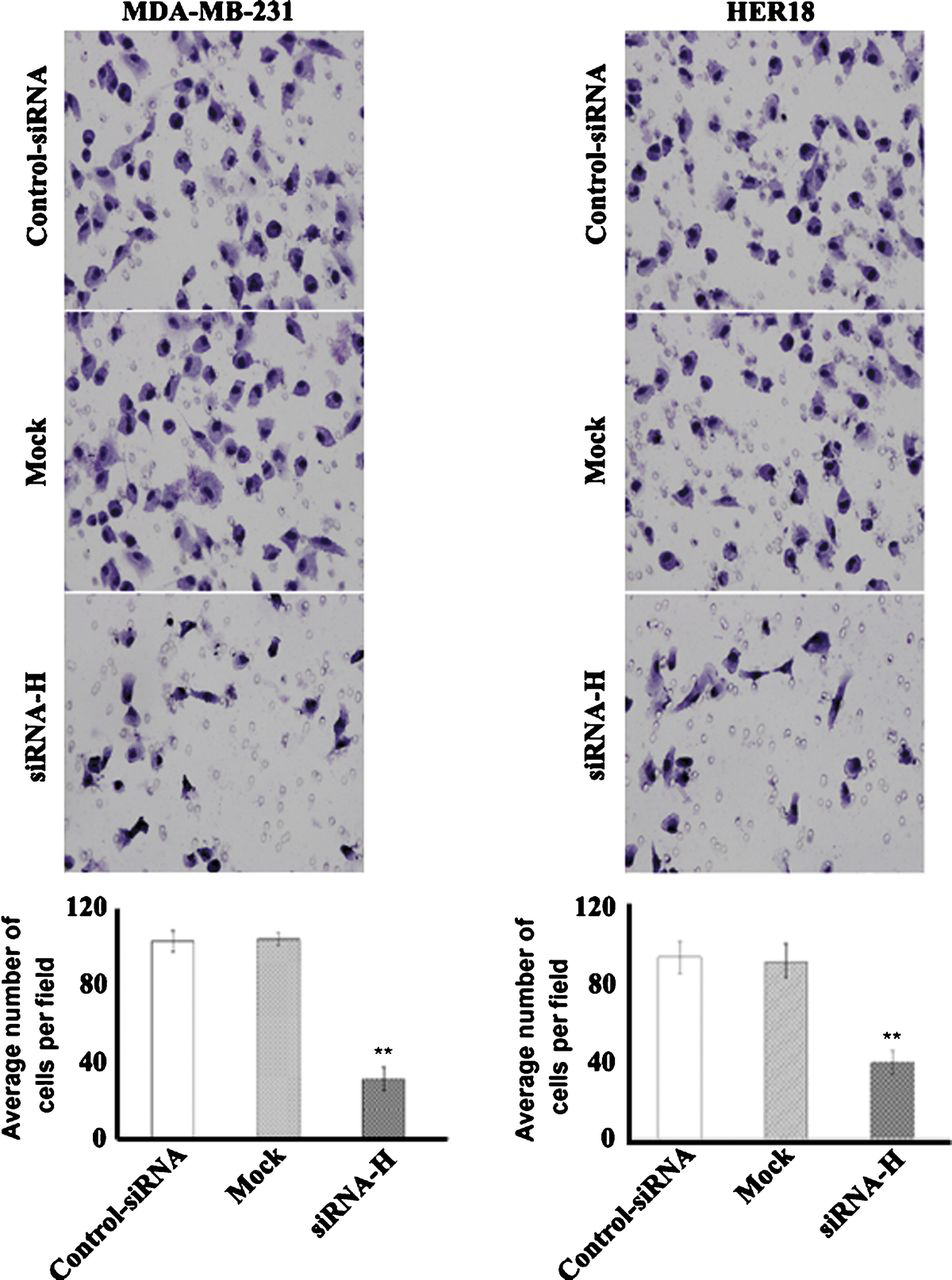
Hepsin silencing decreases cell invasion in MDA-MB-231 and HER18 cells. Cells transfected with hepsin-specific siRNAs (siRNA-H) or control siRNAs or left untreated (mock) were examined for their invasion capacity using an in vitro Matrigel invasion assay. Cells were seeded on the upper surface of the insert membrane and allowed to invade for 22 h. The invaded cells were stained and photographed under a microscope at × 200 magnification. A representative field of the invading cells for each treatment is shown. Bar graphs demonstrate the quantitative analyses of the invasion capacity of cells in top panel.
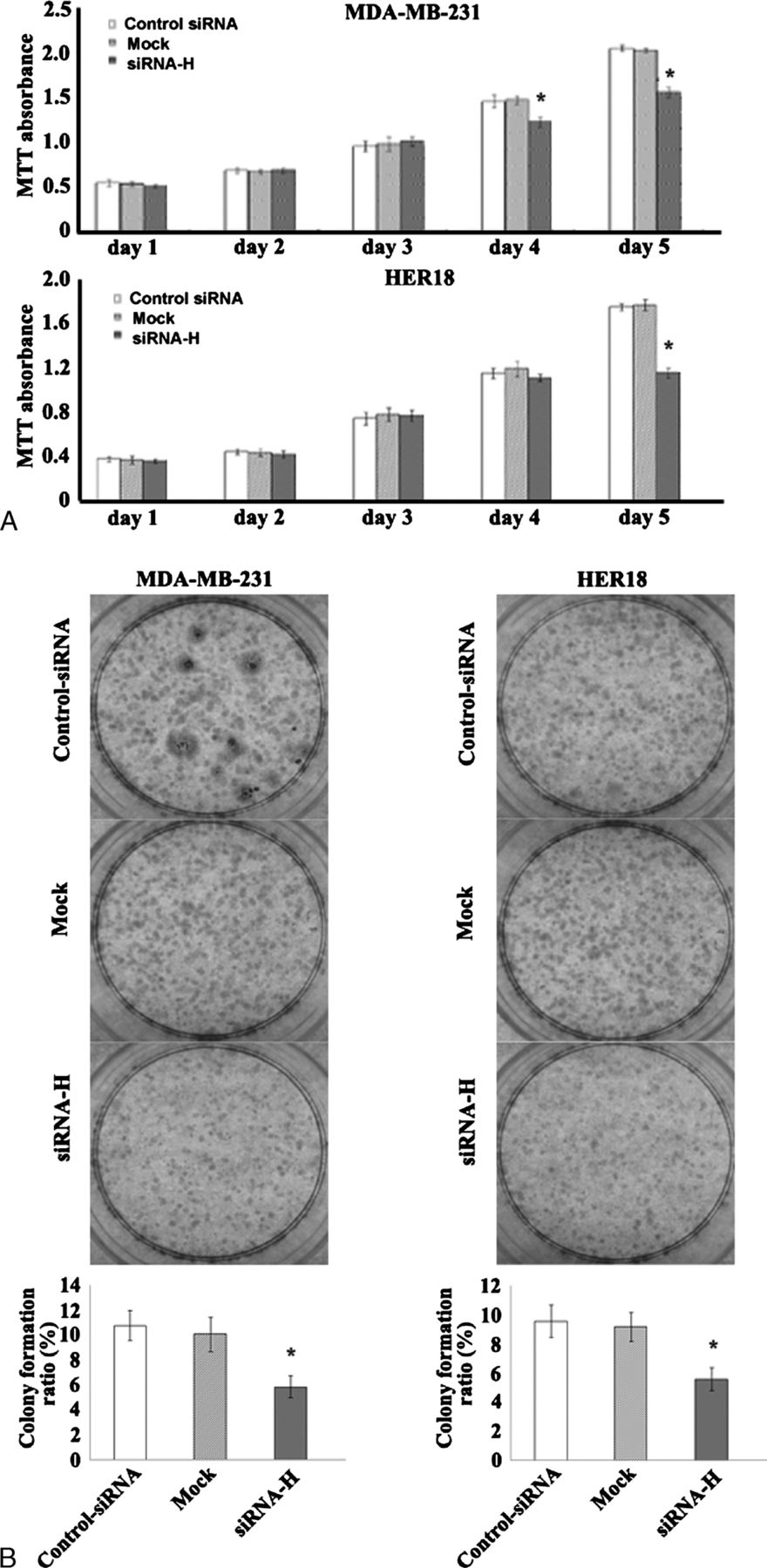
Down-regulation of hepsin inhibits cell proliferation and colony formation of MDA-MB-231 and HER18 cells. A, Assessment of cell proliferation by the MTT assay. Cells transfected with hepsin-specific siRNAs (siRNA-H) or control siRNAs or left untreated (mock) were seeded at a density of 1.5 × 103 cells per well in 96-well microplates, and cultured for 1 to 5 days before determination of the viable cell number. Data represent the mean±SD of 3 independent experiments.
Discussion
In the present study, we show that hepsin is aberrantly overexpressed in breast cancer relative to adjacent nontumor breast tissues. This overexpression is significantly associated with ER and PR positivity, lymph node metastasis, and tumor stage in patients with breast cancer. Moreover, gene knockdown studies indicate that hepsin is required for breast cancer cell proliferation and invasion. These findings highlight an important role for hepsin in breast cancer growth and progression.
Up-regulation of hepsin has been documented in a variety of cancers, including prostate cancer, 22 renal cell carcinoma, 14 endometrial cancer, 23 and ovarian cancer. 6 Clinical association studies have revealed that hepsin overexpression is usually associated with the disease progression and predicts poor prognosis.6,14,22,23 In agreement with the findings in other types of cancer, our data show that hepsin expression is also enhanced in cancerous versus matched noncancerous tissues in patients with breast cancer. Of the 215 studied breast cancer specimens, 143 (66.5%) displayed enhanced hepsin immunostaining. Notably, increased hepsin levels are significantly correlated with ER and PR positivity. Consistent with our histological results, Tozlu et al., 24 using a large-scale real-time PCR approach, have demonstrated a significant elevation of hepsin mRNA levels in ER-positive breast tumors compared with ER-negative breast tumors. Because the levels of both hormone receptors are closely linked to the response to hormone therapy such as tamoxifen in breast cancer.25,26 our data suggest an involvement of hepsin in the estrogen response. However, to the best of our knowledge, it is still unclear whether estrogen had a direct regulatory effect on hepsin expression. Moreover, we also observed a significant association between hepsin overexpression and lymph node metastasis and tumor stage, suggesting that hepsin could contribute to breast cancer growth and progression. Indeed, data from numerous animal models have demonstrated that hepsin has the capacity to promote tumor spread and metastasis.7,27,28 Li et al. 29 have reported that hepsin-overexpressing LNCaP-34 prostate cancer cells are more invasive than low hepsin-expressing LnCaP-17 prostate cancer cells. Moreover, treatment of established LNCaP-34 tumors in mice with a potent hepsin active site inhibitor decreased contralateral prostate invasion and lymph node metastasis, indicating a prometastatic activity of hepsin in malignant disease.
Hepsin is abundantly expressed in some aggressive breast cancer cells such as MDA-MB-231 and HER18, whereas its expression level is very poor in normal breast epithelial cell, MCF-10A. Small interfering RNA-mediated targeting of hepsin in breast cancer cell lines that spontaneously express a high level of hepsin resulted in a markedly diminished capacity of the cells to invade in vitro. Hepsin is a type II transmembrane serine protease, and its enzymatic activity is causally linked to the metastasis-promoting function. Xuan et al. 13 have shown that antibodies neutralizing hepsin protease activity can inhibit invasion of prostate and ovarian tumor cells in culture. Several putative proteolytic substrates of hepsin have been identified, such as hepatocyte growth factor (HGF 30 ), urokinase-type plasminogen activator (uPA 31 ), and laminin-332. 32 Hepatocyte growth factor is of particular interest because it plays critical roles in cancer metastasis and tumor growth by stimulating tumor cell-cell interactions, matrix adhesion, migration, invasion, and angiogenesis.33–35 Hepatocyte growth factor is produced as an inactive precursor, which requires proteolytic conversion. Elevated serum HGF levels have been reported in patients with breast cancer with more advanced TNM staging. 36 Likewise, uPA has been shown to be implicated in breast cancer metastasis. Knockdown of uPA is capable of inhibiting the metastasis of MDA-MB-231 tumors in nude mice. 37 In light of these findings, we speculate that the oncogenic role of hepsin in breast cancer might be mediated, at least partially, through the up-regulation of invasive factors such as HGF and uPA. Further experimental studies need to confirm this hypothesis.
Despite the consistent favorable effects on cell invasion, several previous studies7,38,39 have reported contradictive results regarding the impact of hepsin on cell growth. Torres-Rosado et al. 39 reported that the treatment of hepatoma cells with antihepsin antibodies caused a substantial cell growth arrest, suggesting the requirement of hepsin molecules for normal cell growth. Ectopic expression of hepsin significantly inhibited cell growth in the monolayer, anchorage-independent cell growth in the soft agar in vitro, and tumorigenicity in vivo in ovarian cancer cell lines. 40 In contrast, overexpression of hepsin in a mouse model of nonmetastasizing prostate cancer had no impact on cell proliferation. 7 Srikantan et al. 41 found that exogenous hepsin expression negatively regulated cell growth in metastatic prostate cancer cell lines such as PC-3 and LNCaP cells. These observations suggest that the function of hepsin in cell growth is dependent on cell context. Our data complement and extend the findings in other types of cancer and demonstrate that targeted reduction of hepsin in both breast cancer cell lines, MDA-MB-231 and HER18, significantly reduced cell viability by 25% to 40% after 5-day culture. Colony formation assay further revealed that down-regulation of hepsin decreased the colony formation rate of breast cancer cells. Although the exact underlying mechanism remains elusive, these results underscore a tumor-promoting role for hepsin in breast cancer cells.
In summary, we reported that hepsin was aberrantly overexpressed in breast tumor tissues. This overexpression is significantly associated with lymph node metastasis, hormone receptor status, and tumor stage in patients with breast cancer, and contributes to tumor cell growth and invasion. Thus, inhibition of hepsin expression might have therapeutic potentials against breast carcinogenesis and metastasis.