
Abstract
The question of when the Diné (or Navajo) became a distinct ethnic group has been the focus of research for over a century. Recent archaeological work in the Dinetah region of the American Southwest suggests that the Gobernador phase of Diné prehistory –roughly dated from A.D. 1650 to 1780– was a critical period for Diné ethnogenesis. Archaeological faunas from the Fruitland Data Recovery Project have been used to support this argument; these faunas seem to indicate a shift in subsistence from diets focused on small prey to those focused on larger animals between the Dinétah (A.D. 1500–1650) and Gobernador phases. A reanalysis of the Fruitland Project faunas, involving both refitting of bone and attention to taphonomic issues, does show an increase in representation of large prey during the Gobernador phase; however, this trend does not seem to be related to improved hunting efficiency or encounter rates. Rather, the increase may reflect changes in Diné subsistence and mobility in response to climate change.
Introduction
The cultural landscape of the late prehistoric and early historic American Southwest was diverse both in ethnolinguistic groups and subsistence adaptations (Jorgensen 1983; Opler 1972; Spielmann 1991; Spielmann et al. 2009; Thompson and Tsosie 2008). One of the drivers of this diversity was the migration of Athabaskan-speaking peoples to the Southwest, and the eventual formation of new ethnic identities among these people, including the Diné, or Navajo. Sites distinct from those of the Pueblos, seemingly created by people engaged in more hunting and gathering than earlier groups, began appearing in northwestern New Mexico by the 16th century (Dykeman et al. 2002; Towner 2000; Towner and Heckman 2011). Most archaeologists attribute these sites to Athabaskan-speaking hunter-gatherers who eventually became the Diné. Their apparent subsistence strategy (i.e., more reliance on wild resources than their Puebloan counterparts) is a key part of discussions of early Diné archaeology; however, despite the importance of subsistence in understanding ethnic identity in the Southwest during the 16th and 17th centuries, relatively little zooarchaeological data from this time and region have been published (Gifford-Gonzalez 2011; Pavao-Zuckerman and Reitz 2006). Some paleoethnobotanical data (e.g., Brandt 2002) is available, but our knowledge of contact-era subsistence in the Southwest in general, and for the early Diné in particular, is incomplete.
As Gifford-Gonzalez (2011) points out, in recent years there has been an increase in the use of faunal data in discussions of the Southwestern cultural landscape between A.D. 1450 and 1700. For example, Wilshusen (2010) uses faunal data from the Fruitland Data Recovery Project (Hovezak and Schniebs 2002; Hovezak et al. 2002) to argue for a subsistence shift reflecting increased ethnic cohesion in post-1650 Diné-affiliated sites. Wilshusen (2010) notes that according to Hovezak and Schniebs (2002) smaller fauna dominate the earlier assemblages, while larger fauna dominate later ones, and he argues that this shift from small to large indicates culturally-fostered improved deer-hunting technology. Such a shift, if it is real, is an exciting find: it could reflect factors other than improved hunting efficiency, including an increase in larger mammals available on the landscape, or the introduction of Old World domestic fauna, such as sheep, goat, and cattle. However, the transition identified by Hovezak and Schniebs (2002) and explored by Wilshusen (2010) could also easily be an artifact of taphonomic or methodological processes (Grayson 1984; Lyman 1994, 2008).
To assess the validity of the transition, I re-analyzed the fauna from the Fruitland Data Recovery project, focusing on mechanical and taphonomic issues that could be responsible for the apparent subsistence shifts. In particular, I examined sample size, recovery method, and agent of accumulation. I discuss these results and then explore a number of potential drivers of changing faunal relative abundance in the archaeological record of the early Diné.
Historic Diné Subsistence
The Diné are an Athabaskan speaking indigenous group living in the Four Corners region of the Southwest. Like other Southwestern Athabaskan groups, archaeologists and historians see the Diné as relative latecomers to the area, arriving in northwestern New Mexico sometime after A.D. 1400. While the early archaeological record of the Diné is debated (see discussion later in this paper), the historic and ethnographic Diné have been extensively studied (for example, Amsden 1932; Griffin-Pierce 1992; Iverson 2002; Kelley and Francis 1994; Parezo 1996). Our knowledge of more recent Diné subsistence practices is thus relatively robust.
Unlike their more agriculturally-inclined Puebloan neighbors, the Diné subsistence was remarkably variable through both space and time. Historically, sheep pastoralism, horticulture, and hunting and gathering were all part of the their subsistence regime. As environmental conditions changed, Diné households increased or decreased the resources allocated to each subsistence component in response. When rainfall was plentiful, for example, the size of sheep herds would be increased; in dry years, the extra sheep would be butchered (Roessel and Broderick 1974; Weisiger 2009). Richard White (1988) argues that the balance between these different components allowed the Diné to buffer the risk associated with living in a highly variable environment.
Diné subsistence thus incorporated many “non-traditional” aspects. While the initial Athabaskan settlers were almost certainly exclusively hunter-gatherers (Haskell 1987), they appear to have adopted horticulture from the Puebloans quite early (Sesler and Hovezak 2002). The importance of hunting may have decreased as sheep pastoralism became widespread in the late 18th and/or early 19th centuries (Weisiger 2009; White 1988). When Western commodity foods became more available in the late 19th century, these replaced dryland farming in the “subsistence switching” system (White 1988).
Because change through time is a critical component of Diné subsistence, both transitions in resource use and adoption of new resources are obviously topics of importance to archaeologists interested in early Diné archaeology and, more broadly, Southwestern environment and human subsistence. In addition, the relevance of these questions extends far beyond the Southwest, as the “subsistence switching” strategy was not unique to the Diné, but was present among a number of horticulture-practicing North American groups (White 1988). Colonialism provided opportunities for the expansion of such systems, but as White shows, also carried hazards for their collapse.
Foraging Theory and Subsistence Transitions
The proposed transition in Diné subsistence at A.D. 1650 is intriguing for a number of reasons. Wilshusen (2010) sees the subsistence shift as a line of evidence that can tie Diné oral history to chronological time, and therefore a source of information about the origins of a distinct Diné identity – in other words, Diné ethnogenesis. However, an increase in the relative abundance of large game in human diets could also indicate a change in availability of prey. The prey choice model of optimal foraging theory (Stephens and Krebs 1986) provides a theoretical framework to link transitions in prey choice to changes in landscape abundance (e.g., Broughton 1994; Lupo 2007). This model predicts which resources, given certain assumptions, a forager will pursue once a prey item is encountered. Potential prey types are placed in a rank-ordered set, with the ranking based on their post-encounter energetic return rates –that is, how much energy (or some other currency) is returned from prey of a certain type given pursuit and handling costs. In archaeological contexts, where post-encounter return rates cannot be measured directly, faunal resources are typically ranked according to size, with larger mammals (up to a certain size) assumed to have a higher post-encounter return rate than smaller ones. Resources will be added to, or dropped from, a forager's optimal set according to efficiency rank order. If encounter rates with higher-ranked resources decline sufficiently, a wider and wider array of increasingly lower-ranked prey types will be taken.
In the context of foraging theory, diets dominated by lower-ranked resources –generally small prey items– may indicate a decline in available higher-ranked resources (often larger prey, such as deer) on the landscape, while diets rich in higher-ranked resources may suggest a plenitude of available higher-ranked prey. If indeed there was a transition from small-game dominant diets to larger-game dominant ones among the Diné around A.D.1650, this could indicate increasing encounter rates with larger prey items.
Increased encounter rates can be caused by a variety of factors. Improved hunting technology (as proposed by Wilshusen 2010) is one; decreasing transport costs, as might be expected with the introduction of horses, is another. But increasing encounter rates with higher-ranked prey might also simply indicate reduced pressure on higher-ranked resources due to decreased hunting. Hunting decreases are known to have happened in many regions of North America following indigenous population declines. Though the impacts of European diseases on the Native Americans of New Mexico continue to be debated (Kulishek 2010; O'Fallon and Fehren-Schmitz 2011; Ramenofsky 1996), shifts in settlement in conjunction with the Pueblo Revolt (1680–1692) are well documented (Liebmann 2011; Liebmann and Preucel 2007; Preucel 2011). Changes in the frequency of hunting in particular environments –whether due to changing settlement patterns or to a reduction in the number of people hunting– might have allowed New Mexican environments to recover from previous depletion. Such environmental recovery has been documented, both in the zooarchaeological record and in other datasets, in a number of other regions (e.g., Butler 2000; Davies et al. 2004; Denevan 1992; Endfield and O'Hara 1999; Kittinger et al. 2011). A documented landscape rebound in 17th-century New Mexico would be quite significant, as this would provide an independent line of evidence for demographic change, something that is sorely needed (Kulishek 2010). However, at this point such possibilities are merely speculation. A number of other factors need to be considered to verify that the apparent transition did in fact take place and to interpret what it represents.
Zooarchaeological patterns are not only driven by changes in the cultural and/or environmental landscape; taphonomic issues, such as variable rates of bone fragmentation, differences in collection method, and changing contributions of non-human foragers can also cause an apparent shift in relative abundances (Grayson 1984; Lyman 1994, 2008). Before cultural and environmental changes can be explored as potential causes for this shift, mechanical issues must be ruled out. In addition, optimal foraging models were developed to explain changes in foraging behavior, but by the 18th century and possibly earlier, the Diné increasingly moved from foraging toward food production (Carrillo 1992; Thompson 2009; Weisiger 2004). In such a situation, optimal foraging logic must be applied with caution.
Once such factors are addressed, possible causes of the transition can be explored using measures of diversity: richness, evenness, and nestedness. If indeed the increase in representation of large mammals reflects increased encounter rates with higher-ranked prey, then, using the logic of foraging theory, the diversity of the faunal data set should decrease through time, as lower-ranked prey are dropped from the diet (Jones 2004). Richness, or in a zooarchaeological context, the number of taxa represented (NTAXA), can be used as a measure of diet breadth (e.g., Grayson and Delpech 1998; Lyman 2008; Nagaoka 2002). In a case of increasing encounter rates with higher-ranked prey, diet breadth, and thus NTAXA, would be expected to decrease through time.
Dietary evenness, or the degree to which taxa are equally represented in an assemblage, measures a different aspect of diversity and is less plagued by issues of time averaging than NTAXA, and thus works well alongside NTAXA (Jones 2004; Lyman 2008; Magurran 1988). In a case of increasing encounter rates with higher-ranked prey, evenness would also be expected to decline through time, as diets increasingly focus on those higher-ranked types (Jones 2004).
Finally, foraging theory suggests that if encounter rates with higher-ranked, large prey types were increasing, later faunal assemblages should be subsets of earlier ones. Nestedness measures calculate the degree to which one assemblage is a subset of another (Atmar and Patterson 1993). In a zooarchaeological context, if there are fewer taxa from one chronological unit than another, but all the taxa from that first chronological unit are contained in the second, then the first unit is a subset of the second and the assemblages would be perfectly nested. This situation, where lower-ranked types are dropped from the diet as encounter rates increase, is exactly what would be expected in a foraging theory situation. Nestedness measures are thus an excellent tool for assessing the possible faunal transition discussed here (Jones 2004).
Diné Archaeological Chronology
Archaeological research on Diné origins suggests that Athabaskan hunter-gatherers entered the San Juan Basin area by the 16th century (Towner and Dean 1996; but see Schaafsma 2002 for an alternate interpretation). The first documentary reference to the early Diné dates to the 17th century (Zárate Salmerón 1966). The development of the Diné subsistence system from their initial settlement up to the complex mixed horticulture and pastoralist adaptation documented in the 19th century, and the concurrent and inter-related development of a distinct Diné ethnic identity, is one of the major research questions in Diné archaeology today (Hovezak et al. 2002). Hester (1962) established a basic two-phase chronological system for Diné prehistory: the initial Dinetah phase, and the later Gobernador phase.
The Dinetah phase is defined as the earliest period of Athabaskan occupation in the San Juan Basin. Sites from this period typically contain some combination of brush structures and/or forked-pole hogans, Dinetah Gray pottery, ceramic and lithic scatters, and hearths. Although the Dinetah phase peoples are generally characterized as mobile hunter-gatherers (Hester 1962), there is evidence from the Fruitland Project and other sites to indicate that the ancestral Diné of this phase were practicing at least some maize agriculture (Brandt 2002; Hovezak et al. 2002). Likewise, there appears to have been significant interaction between Dinetah phase ancestral Diné and their Puebloan neighbors (Hancock and Moore 1988; Hester and Shiner 1963; Kearns 1988; Mills 1989; Wharton et al. 1996), even though such interaction has more often been considered a hallmark of the Gobernador phase.
Hovezak et al. (2002) consider Dinetah phase sites as dating between ca. A.D. 1500 and 1650, and in the work presented here I follow this convention as well. It should be noted, however, that A.D. 1690 is more commonly cited as the end date for Dinetah phase sites (e.g., Wharton et al. 1996). Hovezak et al.'s reasons for starting the Gobernador phase early are discussed below.
The Gobernador phase has traditionally been defined as beginning after the Pueblo Revolt (Hester 1962), but based on data from the Fruitland Data Recovery and associated projects, Hovezak et al. (2002:67) see “the formation of a cohesive tribal identity” among the Diné as early as 1650, and thus define the Gobernador phase as beginning then. The Gobernador phase ends with the transition to the Cabezon phase in the mid-to-late 18th century, when the Diné abandoned Dinétah and moved to the west.
Hovezak et al.'s decision to push the start of the Gobernador phase back in time is logical, given that the Gobernador is often considered to be the time period when a distinct Diné ethnic identity solidified (Brugge 1963; Preucel 2011; Wilshusen 2010). Settlement patterns are a major part of this argument: during the Gobernador, Diné settlement seems to have changed, with the development of “residential clusters” of family groups (Wilshusen et al. 2000). Economic diversification, as evidenced by faunal and botanical data, is also sometimes cited as evidence for Diné ethnogenesis in the Gobernador (Sesler and Hovezak 2002).
Whether or not the Gobernador was the key period in the formation of a distinct Diné identity, there are a number of historical events and changes in material culture that date to this time period. Researchers typically characterize the Gobernador phase as a period of increased Diné-Puebloan interaction, evidenced by increasing frequency of Puebloan ceramic types. This increasing interaction may possibly –though not necessarily– have been due to refugees following the Pueblo Revolt and Spanish reconquest in the late 17th century (Hogan 1991; Liebmann and Preucel 2007; Preucel 2011; Towner 2003). The Gobernador phase was also a time of increasing conflict with raiding groups such as the Ute and Comanche, as such raids became an increasing problem by the early 18th century (Towner 2003). Other characteristics of Gobernador phase sites include the presence of locally-made Gobernador Polychrome in addition to the Puebloan ceramic types; specialized lithic tools, bone and shell ornaments, and European trade goods; a variety of architectural and feature types in addition to brush structures and forked-stick hogans; and possibly increasing dependence on sheep (Bailey and Bailey 1986; Hester 1962; Hill 1940; Hovezak et al. 2002; Madrid et al. 1996; Weisiger 2004).
Methodology
Wilshusen's (2010) argument is based on the faunal data from the Fruitland Data Recovery Project (Hovezak and Schniebs 2002), an investigation by La Plata Archaeological Consultants between 1991 and 1994 of more than 40 sites in the Fruitland coal gas development area in northwestern New Mexico (Figure 1). Though the Fruitland project included excavation of sites dating from the Archaic through the historic periods, the Diné-affiliated sites from the late prehistoric and early historic are the focus of this study. The Fruitland project area is in the heart of Dinétah, the traditional Diné homeland and the location of some of the earliest known Diné sites in New Mexico.
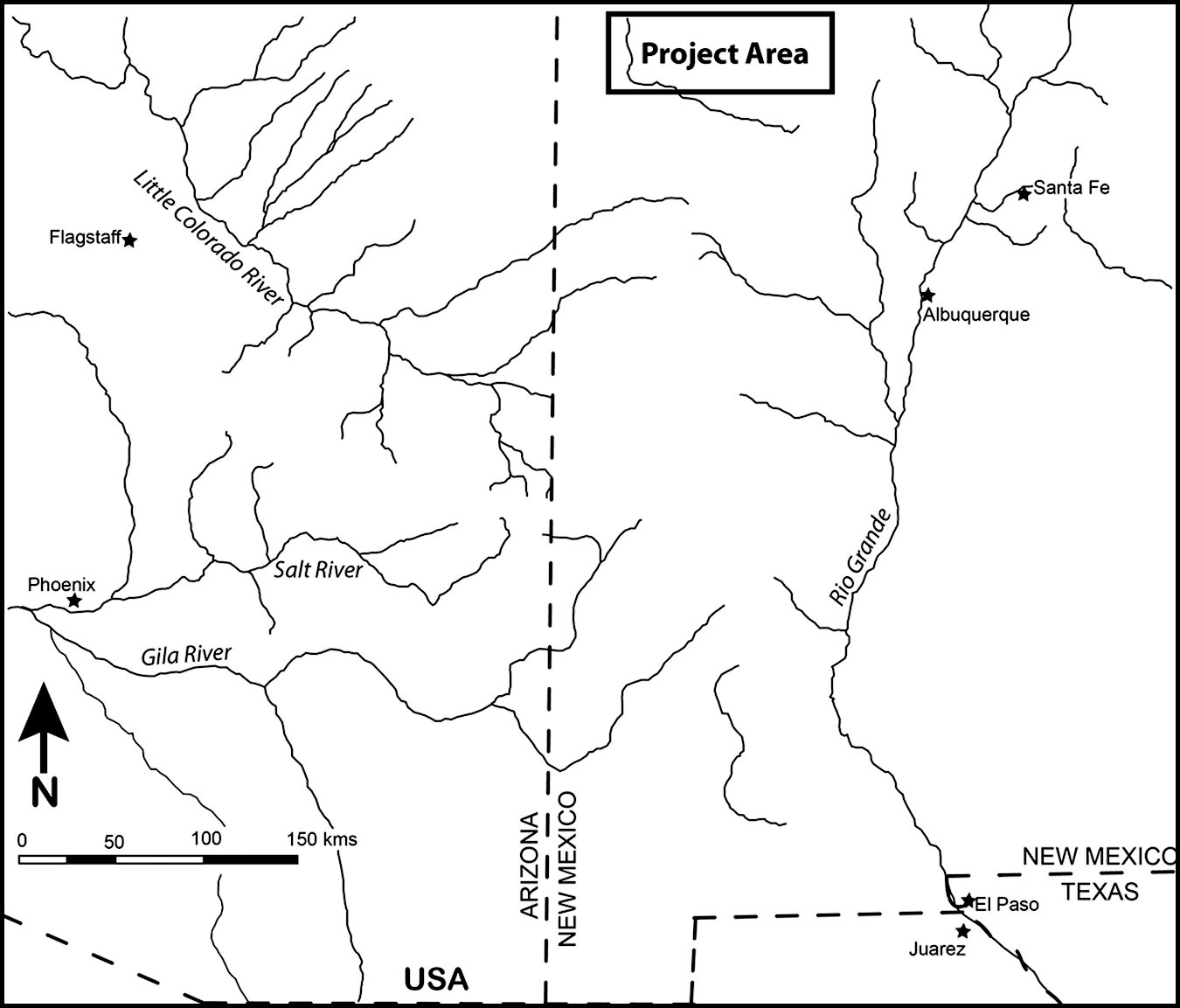
Map of the Fruitland Data Recovery Project area.
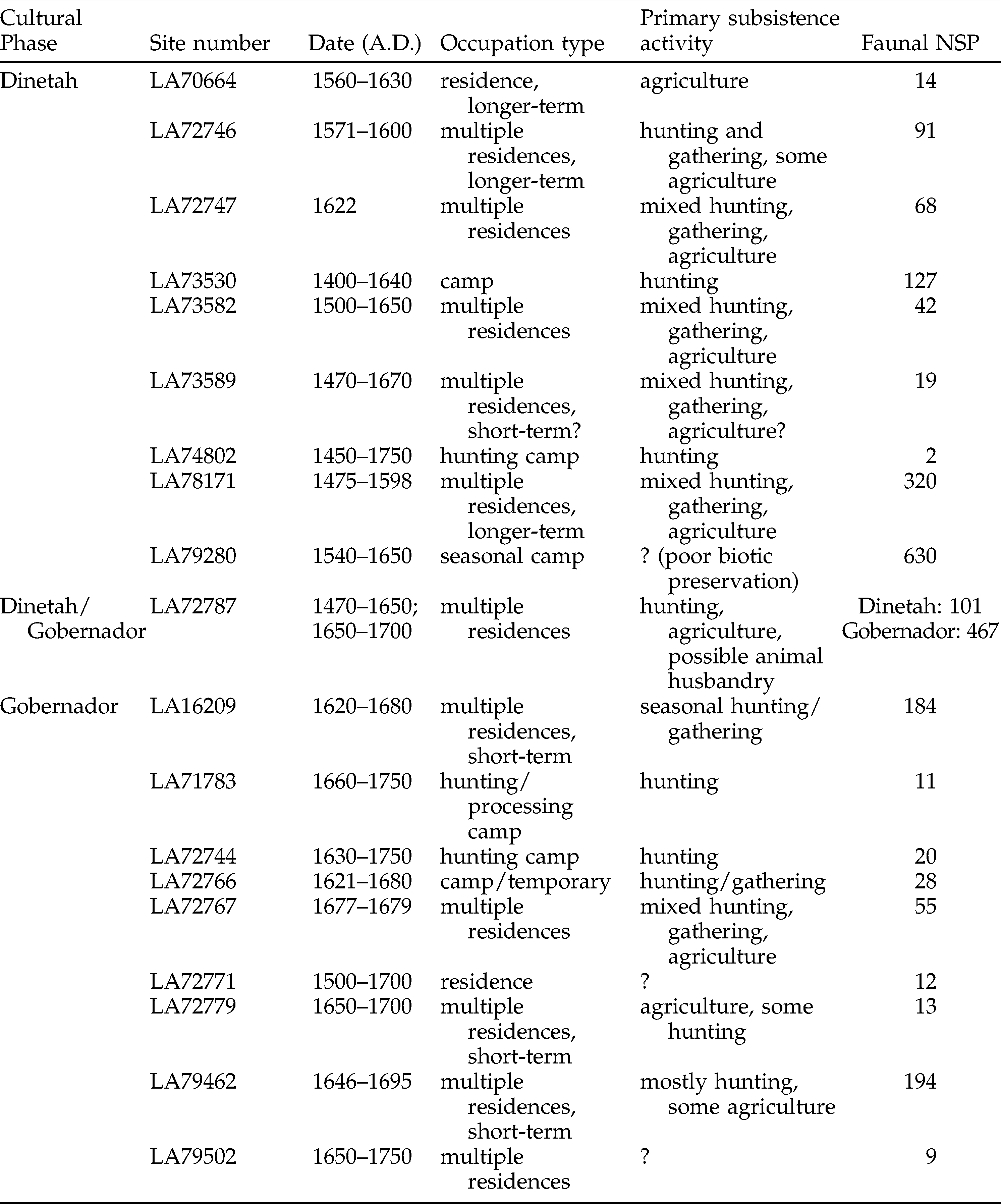
Early Diné subsistence and settlement was a major component of the research design for the Fruitland Project. Recovery methods were selected to maximize the potential information gained from faunal and botanical remains; in some (but not all) contexts, 1/8” screens or smaller were used. The resulting faunal dataset was extremely robust. Diné-affiliated sites from the project containing faunal remains are listed in Table 1, along with the initial numbers of specimens (NSPs) of fauna reported by Hovezak and Schniebs (2002). The Fruitland Data Recovery Project has had a significant impact on our understanding of the early Diné archaeological record. The data generated by this project has not only increased knowledge of early Diné subsistence and settlement, it has contributed to overall Diné archaeological chronology.
Descriptive information for sites included in this analysis (after Hovezak and Schniebs 2002; Hovezak and Sesler 2002a, 2002b; Hovezak et al. 2002). NSP indicates number of specimens identified in Hovezak and Schniebs’ initial analysis (2002).
To explore the transition in Diné subsistence identified by Hovezak and Schniebs (2002) and referenced by Wilshusen (2010), I reanalyzed the faunal collections from the Fruitland Data Recovery Project, with two main objectives. First, I hoped to increase the number of specimens identifiable to genus or species level. Second, I wished to obtain additional taphonomic information to verify that this transition was not the result of a change in primary accumulator (for instance, a shift between carnivore-deposited faunas and human-collected ones), collection method (i.e., a change in screen size used by excavators), or similar processes.
The Navajo (Diné)-affiliated Fruitland fauna were extremely fragmented (Hovezak and Schniebs 2002), and through a systematic refit process I hoped to gain finer species resolution. All fragments were examined and identified using the comparative collections of the Utah State University Brigham City zooarchaeology lab as well as collections at University of New Mexico's Maxwell Museum of Anthropology and the Southwest Museum of Biology. Refits were attempted within the collection from each site.
If many of the small mammals found in Dinetah-phase aged sites were intrusive, while the larger prey (primarily artiodactyls, such as deer and pronghorn) found at the Gobernador phase sites were the result of human activity, then obviously the switch in dominance from small faunas to large would not indicate changing human subsistence, but would have a different meaning. So I attempted to identify fauna that were accumulated through non-cultural processes. I used three main lines of evidence to do this. First, I referenced the site-specific contextual information provided in the published data report (Hovezak and Sesler 2002a, 2002b). Second, I recorded surface modifications: toolmarks, weathering (I used the scale in Behrensmeyer 1978 and made the assumption that intrusive rodents were likely to be less weathered than bone deposited by cultural activity), traces of carnivore activity, or burning. And third, I analyzed element completeness, as small mammal elements used by humans as food are generally less complete than those deposited through other means (Hockett and Haws 2002). Based on this analysis, I characterized each specimen's likely origin: intrusive, non-human predator, cultural, or unknown. Because the screen size used in excavation can also have a disproportionate effect on the recovery of small fauna (Cannon 1999; Schaffer and Sanchez 1994; Stahl 1996), I coded each specimen as to screen size used, and tested for a relationship between the proportion of fauna recovered using a 1/8” or smaller screen and the proportion of small fauna in the sample.
Results
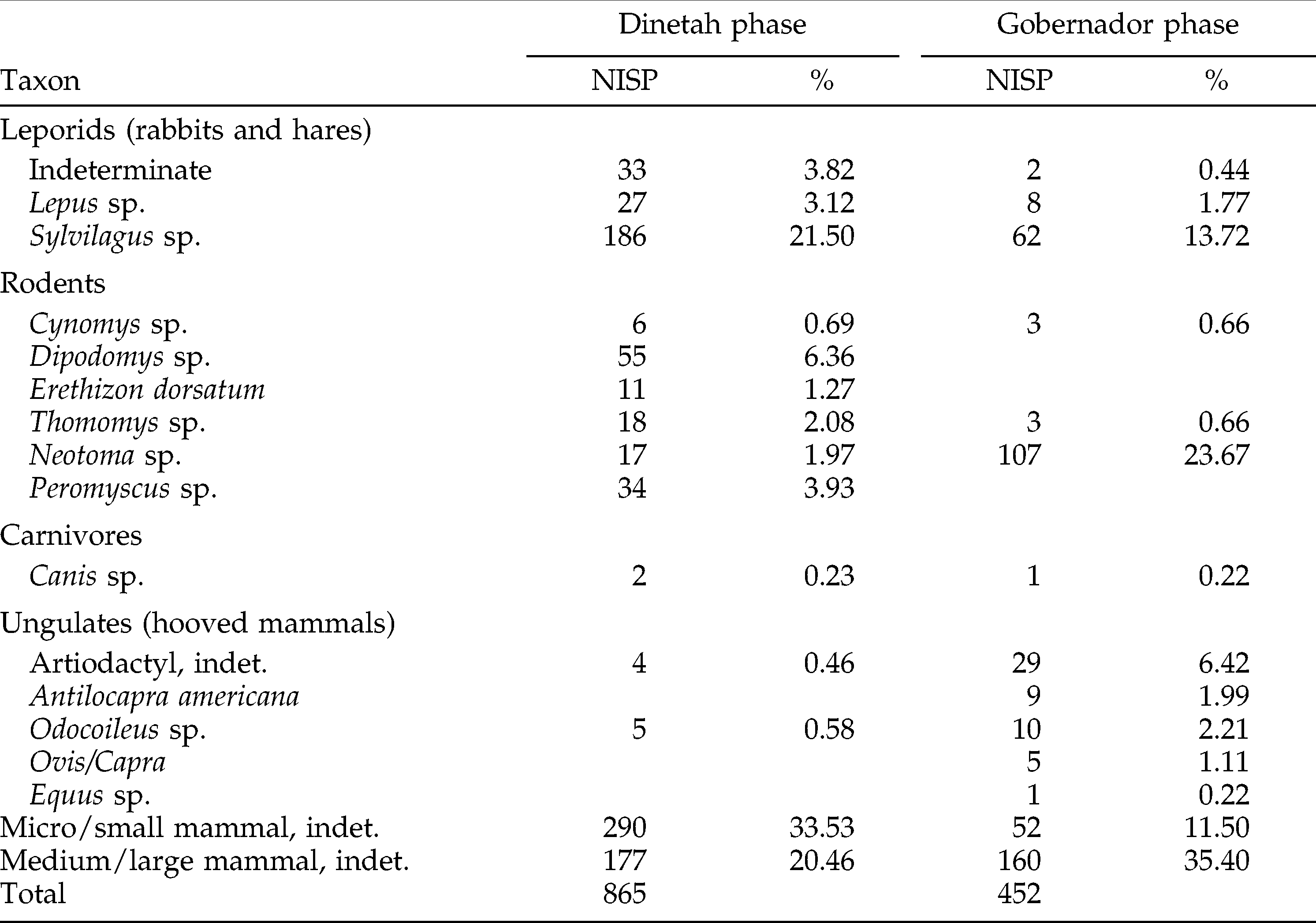
The reanalysis project resulted in a slight increase in species resolution among the artiodactyls (Table 2); refitting of bones allowed some specimens previously identifiable only to order to be identified as deer (Odocoileus sp.) and pronghorn (Antilocapra americana). I focused my analysis on the mammalian fauna, and included only those specimens that I could reliably identify to a particular taxonomic group, or to small/micro mammal (hare or smaller) or medium/large mammal size classes (pronghorn or larger). This resulted in some additional differences from Schniebs and Hovezak's analysis, notably a reduction in the overall number of specimens. However, the increase in relative abundance of large prey items in Gobernador phase sites identified in the original analysis remained striking (Figure 2). Sample size is only insignificantly correlated with the patterns observed in Figure 2 (r = +0.72, p = 0.4887); likewise, screen size does not appear to be correlated with the relative abundance of small mammals (r = −0.037, p = .976).
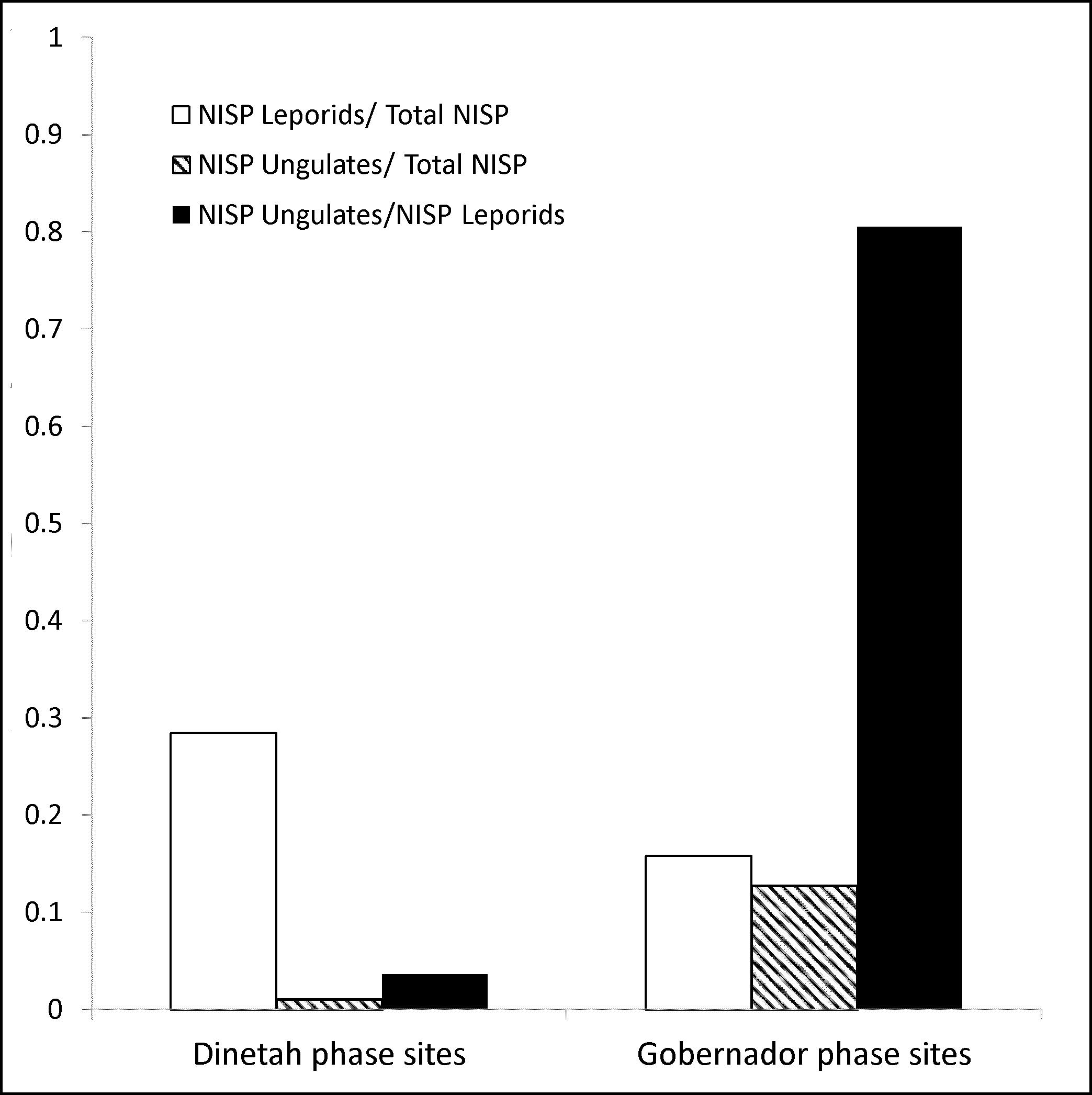
Relative abundance of different prey types in Dinetah and Gobernador phase sites.
Mammalian fauna from the Dinetah and Gobernador phase sites of the Fruitland Data Recovery Project identified as part of this project.
Not all the small fauna seem to have been deposited by human activity. Many of the small mammals, particularly the rodents, appear to have been intrusive, rather than subsistence items (though rodents were often consumed in the prehistoric Southwest; see Matthews 1994; Szuter 1991). In contrast to the rest of the collection, the rodent specimens tended to be complete and unmodified. No toolmarks were observed on them; only 4 of 154 specimens show any signs of burning; and most (99%) received the lowest possible weathering score. Hovezak and Schniebs (2002) noted several rodent specimens in contexts that they believed were cultural, but I was unable to identify these individuals. Because most of the rodents are probably not cultural, I needed to mitigate potential bias from intrusive rodents in the overall relative abundance of ungulates. I thus calculated the ratio of ungulates (hooved mammals) to leporids (rabbits and hares). As can be seen in Figure 2, the pattern of increase seen in the relative abundance of ungulates is also present in the ratio of ungulates to leporids (Dinetah: NISP Ungulates/NISP Leporids = .04; Gobernador: NISP Ungulates/NISP Leporids = .81), as it would not be if decreasing representation of non-cultural rodents were driving the change.
Since the shift appears to be real and not the result of taphonomic processes, I wanted to know if it was the result of a change in prey availability. If Diné foragers were becoming more specialized large-game hunters, whether due to environmental rebound or increased hunting efficiency, one would expect increased encounter rates with higher-ranked prey. NTAXA would be expected to decrease from the Dinetah to the Gobernador phase. In the Fruitland assemblages, however, NTAXA remains the same (NTAXA = 10). When rodents are excluded from the analysis, NTAXA actually increases, with a value of 4 in Dinetah phase sites and 7 in the Gobernador (Table 3). While the difficulties in using NTAXA as a measure of diet breadth (Grayson and Delpech 1998; Lyman 2008) could be responsible for this lack of trend, evenness also failed to identify any changes in diversity across the transition. I used the reciprocal of Simpson's index (1/D) to measure evenness (Jones 2004; Magurran 2004); in the Dinetah phase sites, 1/D was 1.35, while in the Gobernador, 1/D was 1.39.
Presence/absence matrix for the Fruitland Data Recovery Project fauna.

Finally, if Diné diets were becoming more specialized, we might expect later faunas to be a subset of earlier ones (Jones 2004); that is, we would expect these faunas to display a high degree of nestedness (Atmar and Patterson 1993). A presence/absence matrix of the species represented in the faunas from each phase (Table 3) suggests that this is not the case; if anything, Dinetah phase faunas are a subset of Gobernador phase ones. I used a measure of nestedness called the NODF (nested overlap and decreasing fill) index (Almeida-Neto et al. 2008), calculated with a freely available program called ANINHANDO (Guimarães and Guimarães 2006). This index quantifies the degree of nestedness from 0 (a perfectly nested distribution) to 100 (a perfectly random distribution). An “anti-nested” distribution could be expected to have a NODF value of 50 (Almeida-Neto et al. 2008). The Fruitland faunas are not nested, with a NODF value of 39.
Discussion
Though there does seem to be an increasing focus on large prey in the Gobernador phase faunas, the data presented here do not support the argument that this increase represents increased encounter rates with higher-ranked prey. However, some sort of subsistence transition does seem to have taken place during this time period. The pattern of increased representation of ungulates after A.D.1650 identified by Hovezak and Schniebs (2002) persists even when taphonomy is taken into account, but the changes in assemblage diversity that would be predicted by foraging theory are not present.
An obvious explanation for this involves the adoption of Old World domestic faunas. If the Diné adopted sheep (or other domestic faunas) in larger numbers during the Gobernador phase, then a relative increase in ungulates might make sense; and the move to a more pastoral economy might make the application of foraging theory problematic because foraging theory was designed for hunter-gatherer economies. The Diné-affiliated faunas from the Fruitland Data Recovery project do contain sheep/goat (Ovis/Capra) and horse/donkey (Equus sp.) specimens. However, Sesler and Hovezak argue against a large-scale adoption of domestic fauna at this time, as these specimens represent only a small part of the assemblage (Hovezak and Schniebs 2002; Sesler and Hovezak 2002; also see Table 2). A recent study of the faunas at two Puebloan 17th-century New Mexican sites found that, as at Fruitland, there was an increase in relative abundance of artiodactyls in the later 17th century, and again, as at Fruitland, the increase persisted even when domestic fauna were excluded from the analysis (Jones n.d.); the lack of Old World domesticates in all these faunal assemblages suggests that the adoption of these new taxa is not the cause of this trend.
Climate-driven landscape change can affect ungulate frequencies, and must also be considered as a potential cause for the transition. The 16th and early 17th centuries were marked by drought which likely impacted local environments; while drought persisted in some locations throughout the 17th century, other parts of the state appear to have had more precipitation after A.D. 1650 (Barrett 2012; Margolis et al. 2011; Salzer 2000; Stahle et al. 2009). The apparent spatial variability of the precipitation would likely result in a patchy mosaic of landscape rebound –increased grasslands in some areas, but not in others. This type of edgy patchiness often favors higher ungulate populations, and thus would have had a beneficial impact on hunters’ encounter rates with higher-ranked prey. Unpredictable spatial variability in precipitation is, however, not good for agriculture (unless one is along a major water source) because when precipitation is unpredictable, so is the success of a particular field.
Some environmental rebound is suggested by the micromammals from the Fruitland Project faunas, particularly by the relative frequencies of two small rodent taxa: Neotoma sp. and Dipodomys ordii. These taxa are both likely intrusive, as discussed earlier, and so are not good indicators of human subsistence; however, because they have distinct habitat preferences, they can tell us about changing environmental conditions. Dipodomys ordii (Ord's kangaroo rat) is ubiquitous in the project area today and has a strong preference for open environments (Schroder 1987). Neotoma sp., the woodrat, is represented in the project area by three species: Neotoma albigula (the white-throated woodrat), Neotoma mexicana (the Mexican woodrat), and Neotoma cinerea (the bushytailed woodrat). Though these three species have distinct food preferences and N. albigula and N. mexicana are more general in their subsistence than N. cinerea, all three have a preference for green shoots (Dial 1988).
Dipodomys and Neotoma are the two most common rodent taxa in the Diné-affiliated faunal assemblages, but they are present in very different proportions in the Dinetah and Gobernador phase sites (Table 2). Dipodomys is dominant in the Dinetah phase and absent during the Gobernador phase; Neotoma, while present in both phases, is the dominant rodent in Gobernador phase faunas. Without finer species resolution and larger sample sizes, this difference should not be used as the basis for conclusions; but the decrease in Dipodomys relative to Neotoma could indicate an increase in green vegetation, and thus, perhaps some landscape recovery from earlier droughts.
If a simple landscape rebound caused increasing abundance of large mammals, one would expect the predictions from foraging theory to be met; for the hunter, this would mean increasing encounter rates with higher-ranked prey. However, foraging theory predictions must be adjusted if settlement patterns change, because this process can result in the same faunal pattern as would be expected in a case of changing encounter rates (Jones 2007). If an area is characterized by long-term settlements during one time period, and by short-term seasonal hunting camps in a following one, then it is likely that the zooarchaeological record would show an increase in the relative frequency of large mammals. This change could be due to a genuine subsistence shift –for example, when semi-sedentary horticulturalists become mobile hunter-gatherers. It could also, however, reflect a change in landscape use in a larger region (i.e., semi-sedentary horticulturalists who take short, seasonal hunting trips change the locations of both their semi-permanent and hunting camps).
The non-faunal archaeological data from the Fruitland sites –particularly the paleoethnobotanical remains– suggest that changes in settlement and in the importance of different subsistence activities took place between the Dinetah and Gobernador phase sites (Brandt 2002; Sesler and Hovezak 2002). The botanical remains and architecture from the Dinetah phase sites seem to indicate a focus on horticultural activities, with the fauna perhaps reflecting garden hunting; the Gobernador phase sites, on the other hand, appear to be more oriented towards hunting and (in one case) possibly pastoralism (Table 1). This difference could be an issue of changing use of the project area; perhaps the project area was home to agricultural sites early on, and hunting sites later. But it could also reflect a fundamental subsistence transition, where the Dinetah phase ancestral Diné focused on horticulture, while the people of the Gobernador phase were more oriented towards hunting (Sesler and Hovezak 2002; Wilshusen 2010). As discussed earlier, such switching between subsistence activities is a well-documented risk reduction strategy among the historic Diné and other peoples (Weisiger 2009; White 1988).
This returns us to the issue of landscape change because among people who practiced subsistence switching, changing between different modes of subsistence was often a response to environmental variability (White 1988). In the case of the late 17th century, increased (but spatially variable) precipitation would likely have increased large mammal populations due to rebounding grasslands and enlarged edge habitat. The response of the Gobernador-era Diné may have been to devote more resources to hunting. The spatial variability of precipitation likely made horticulture a riskier alternative during this time period. Thus, while a simple reaction to increasing precipitation is not supported by this data, a more complex subsistence-switching response to climate change –one known to have been present among the historic Diné– remains a possibility.
Conclusions
Wilshusen (2010) argues that the increasing relative abundance of large mammals in these faunas reflects the formation of a distinct Diné identity –that this trend indicates a culturally-fostered increase in deer-hunting efficiency among early Diné peoples. The transition from small-game dominant diets to large-game dominant ones initially identified by Hovezak and Schniebs (2002) and referenced by Wilshusen (2010) does appear to be real and not a taphonomic artifact. However, the results presented here do not support an argument for increased hunting efficiency in the Gobernador phase. Rather, they may reflect switching between subsistence types as an adaption to environmental change –a practice documented among the Diné of the 19th and 20th centuries (Sesler and Hovezak 2002; Weisiger 2009; White 1988), when pastoralism was one of several subsistence strategies employed depending on various environmental conditions. More research is needed to verify this interpretation, as a change in the location of different settlement types could also explain the pattern seen here. However, given both the extent of the Fruitland project area (Hovezak et al. 2002) and the known practice of subsistence switching among the historic Diné, a circa A.D. 1650 change in settlement and/or subsistence mode seems the most likely explanation.
What this means for Diné ethnogenesis at 1650 (as proposed by Wilshusen and Sesler and Hovezak) is, I believe, open to debate. Some might argue that the “switching” response is an indicator of an established Diné identity; others might counter that this was a tactic employed by many non-Diné peoples as well. Research from archaeological faunas in 17th-century Mexico suggests that the increase in large mammals seen in the Fruitland data is not limited to Diné-affiliated sites (Jones nd), although such increases are not apparent in all archaeological faunas from this time period (see, for instance, Spielmann et al. 2009). Additional research on faunas from the late prehistoric and early historic Southwest will hopefully further elucidate this situation. The work presented here shows that studies of archaeological subsistence can contribute to our understanding of shifting ethnic identity during the contact period. While the precise ways in which ethnic identity, landscape change, and subsistence practices combined to shape Southwestern archaeological faunas from this period are still unclear, the fact that these did influence each other is evident.
Footnotes
Acknowledgments
This paper was vastly improved by the comments of three anonymous reviewers, by feedback from editors Virginia Popper and Heather Trigg, and by conversations with Don Grayson, Ann Ramenofsky, and Ron Towner. The Navajo Nation and the New Mexico Museum of the American Indian/Laboratory of Anthropology graciously allowed me to examine the collections discussed here. Thanks also to Katie Conrad, Elaine Taylor, and Haylee Toland, who assisted in the initial stages of analysis, and to Cecilia Chávez Beltrán, for expert help with the Spanish abstract.