
Abstract
There are three principal methods humans use to evoke avian vocalizations. This paper explores the two that are constrained by human language: onomatopoeia and warblish. In onomatopoeia, new words are created to mimic sounds within the constraints of a language's phonology. Warblish, named and thoroughly described for the first time in this paper, is the imitation of avian vocalizations using existing words in human language. None of the conventionalized ways humans imitate birdsong has yet been studied rigorously, least of all warblish, for which a term has not even existed. Combing the ethno-ornithological literature for instances of warblish shows patterns in its functions and semantics across cultures, which range from friendly messages to ribald mocking. Warblish may evoke perceived ties between bird calls and other natural phenomena and can even offer clues to historical contact between human communities. Investigation of methods of birdsong imitation may expose alternative folk taxonomies based on vocalization type rather than morphology. As artifacts of human creativity in response to nature, warblish and the other methods of birdsong imitation described here merit interdisciplinary inquiry.
Introduction
The human impulse to imitate birdsong may well predate language. Modern humans can evoke avian vocalizations either through pure sound or through combinations of sound and meaning. Mimicry may be fleeting and idiosyncratic, as when poets or composers draw inspiration from birdsong, or observers imitate birds in novel ways. But as with most aspects of language, human groups tend to conventionalize ways to reproduce birdsong. That is, imitation forms are standardized within a speech community.
Humans speak through manipulation of the lips, tongue, and larynx, with air flowing through the oral and nasal cavities. Birds vocalize by using the muscles around their syrinx to modulate airflow through it. Human speech and birdsong likewise differ acoustically, although some bird species can mimic humans, and gifted humans may imitate birdsong with such accuracy as to provoke avian responses.
There are three widespread ways that humans imitate bird calls, all meriting investigation. One strategy is fully unfettered by linguistic constraints: in non-verbal vocal mimicry, the human attempts to approximate avian chirps, squeaks, whistles, clicks, and other vocalizations as closely as possible, using every available physical means. The other two strategies are constrained to different degrees by human language. With onomatopoeia, birdsong is imitated using nonce words comprising speech sounds of a particular human language. Here, the mimicry is phonetically restricted in language-specific ways. In the third strategy, termed here warblish (see below), imitation takes the form of actual words in the human language. These may combine into phrases or full sentences that are grammatical for speakers. The meanings of warblish phrases may be humorously incongruous with the birds to which they are attributed or the warblish may fit with the bird's mythic character, its behavior, or its place in the ecosystem. Warblish is largely constrained by human linguistic structure, although it may diverge from ordinary language at the levels of syntax, semantics, or pragmatics.
The existence of the first two strategies is tacitly or explicitly acknowledged in the ethno-ornithological literature. Non-verbal vocal mimicry is termed simply “imitation” by Berlin (1992:243), but this is clearly imprecise, covering all three strategies. (Earlier, Berlin and O'Neill [1981:239] called it “literal mimicry.”) Onomatopoeia is an accepted descriptor for the second strategy, occurring in many ethno-ornithological studies (e.g., Forth 2010; McElhanon 1977). As a category, what I term warblish in this paper has been largely overlooked by researchers. At a special ethno-ornithology session of the Society of Ethnobiology in 2015, scholars acknowledged that this strategy lacked even a name. (Some of the fumbling ways people have described warblish are in the section on warblish, below.) I define warblish as:
This paper aims to establish conventionalized birdsong mimicry, in particular warblish, as an area of investigation in its own right. Supporting details are drawn from ethno-ornithological studies, from birdwatching aids, and from personal communications with ethnobiologists, linguists, and birdwatchers. The first section provides a short review of the ethnolinguistics of birdsong imitation. The second section outlines non-verbal mimicry. The third section deals with onomatopoeia. The fourth section introduces warblish and is followed by a conclusion. Research into how humans imitate birdsong is in its infancy. Suggestions for further investigatory directions accompany introductions of the imitation strategies.
The Ethnolinguistics of Birdsong Imitation
As in many other sub-fields of ethnobiology, ethno-ornithological studies tend toward preoccupation with the forms and etymologies of bird names. Hunn and Brown (2011:319) even define linguistic ethnobiology as “study of the naming of plants and animals.” It is standard for the descriptive component of an ethno-ornithological article to be structured as a list of bird names, with a translation for each name or speculation on its origin, and usually notes on cultural significance (Anonby 2006; Cooke 1884; Diamond and Bishop 1999; Forth 1996; Hunn 1991; Hunn and Thornton 2010; Loss et al. 2014). The distribution of binomials versus simplex terms, the presence or absence of generic names, and the mapping from indigenous to Linnaean names are of central interest. Bird names in particular have even been the basis for a much broader claim about the origin of words in indigenous societies, by Berlin and O'Neill (1981:259).
But if ethnobiology is defined in terms of “cultural knowledge” (Anderson 2011:1), then its overlap with linguistics need not be limited to nouns and their etymologies. Covert linguistic indications of what people know about their ecosystems may be found, not just in names, but also in other types of words and even grammatical structures. For instance, nominal classifiers are particles that accompany nouns and serve to demarcate their membership in one of a number of classes. These can reveal culturally salient ways of classifying the natural world, as Messineo and Cúneo (2011) show for Toba and Maká (South America). Cultural knowledge about plants and animals may also be encoded in the gender system of a language. In Dyirbal (Australia), there are four genders (Dixon 1972, 2015:21–42). Animals and plants are divided among the four genders in ways that accord with Dyirbal knowledge of the natural world. For example, most fish belong to Gender I, but three harmful fish belong to Gender II; similarly, most non-edible plants belong to Gender IV, but four with stinging hairs belong to Gender II. Insect-eating bats belong to a different gender than fruit-eating bats, who share this gender with other vegetarian mammals such as kangaroos and possums (Dixon 2015:40). As will be discussed below, cultural knowledge of a community's environment may be embodied in the conventional ways that people reproduce ambient sounds, including those of fauna. There are likely many other ways that language can reflect cultural knowledge of biology.
A bias toward the collection of names and comparison with Linnaean categories has led researchers to overlook alternative taxonomies based on vocalizations rather than morphology. The widespread existence of such taxonomies is logical; as Ng'weno (2010:107) writes for Kenyan societies, people may hear birds more regularly than they see them. One example of a classificatory category based on vocalization is the broad category of birds and other beings that “say po” among the Nage of Indonesia (Forth 1998, 2004:63). According to Feld (1982:72–82), the Kaluli (Papua New Guinea) divide birds into seven groups based on vocalization type. The groups are: birds that “say their names,” “make a lot of noise,” “only (produce) sound,” “speak the Bosavi language,” “whistle,” “weep,” and “sing gisalo song.” Two different call types may be produced by the same bird, as with the White-crowned koel (Cacomantis leucolophus) (Feld 1982:79–80). Vocalization types and patterns are of such importance to the Kaluli that they consider the New Guinea friarbird (Philemon novaeguineae) and Brown oriole (Oriolus szalayi), which have similar calls and call together at times, to be the dimorphic male and female of a single bird,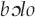 . As Feld writes, here “the criterion for sharing a taxon is aural” (1982:57).
. As Feld writes, here “the criterion for sharing a taxon is aural” (1982:57).
Even where researchers are attuned to taxonomies based on vocalization type, they do not necessarily examine conventionalized human imitations of these vocalizations. Feld's (1982) study of Kaluli taxonomy of avian vocalization type focuses on perception, not reproduction. He gives examples of onomatopoeic call renditions in relation to the “say their names” category (Feld 1982:72–73), but otherwise strategies for imitation must be inferred from his descriptions or are simply absent. That is, “speak the Bosavi language” calls are apparently imitated with warblish and “whistled” calls perhaps by non-verbal mimicry, but there are no clues as to how Kaluli people evoke the remaining four call types described by Feld. In comparatively standard studies that follow the name-etymology mold, birdsong imitations are mentioned sporadically, usually subsumed in the descriptive texts following bird names (except in Agnihotri and Si [2012] and Berlin and O'Neill [1981]). Onomatopoeic renderings are most likely to be noted when they are also the sources of bird names.
Linguistics is more than the study of noun etymologies. Linguistic ethnobiology should examine all ways in which language reflects cultural knowledge of the natural world, not just names. Among these are conventions for the description and mimicry of natural sounds, including birdsong.
Linguistic ethnobiological research must always take into account the degree to which an aspect of language is conventional, that is, shared in the same form by members of a speech community. Individuals may choose to innovate unconventional birdsong imitations using any of the three strategies, creating their own unique onomatopoeia, warblish, or non-verbal mimicry. Scholarship that mentions birdsong imitations tends to lack discussion of the degree to which imitations take a set form within a community. Researchers largely take conventionality for granted, disregarding both potential individual creativity on the part of informants and the possibility that more than one conventional imitation are recognized by one community. In their list of onomatopoeic bird call reproductions, Berlin and O'Neill (1981:260, n 1) acknowledge:
We have not addressed the problem of informant variation in this paper. We do not know to what extent the onomatopoeic bird vocalizations recorded from the four Aguaruna and Huambisa informants…are shared for [sic] the population as a whole.
Despite this acknowledgment, Berlin and O'Neill are silent on a related point of methodology: the way they selected the single Huambisa onomatopoeic rendition they give for each bird on their list. Since three Huambisa men provided data, one might suppose that the single imitation given was determined by consensus, but the authors do not discuss this, nor the extent to which the men's individual imitations differed, if at all.
Hypothetically, any of the three birdsong imitation methods described here could be used to mimic any bird call, but actual usage depends on function, conventionality, and context. Non-verbal mimicry is likely the preferred choice when attempting to actually communicate with birds. When evoking the bird call in conversation with other humans, convention and context may determine the strategy used. When asked for it, Berlin and O'Neill's (1981:239) Aguaruna and Huambisa respondents produced non-verbal mimicry for “all birds that call”; when asked for onomatopoeia, the respondents also produced onomatopoeic imitations for most calls. Berlin and O'Neill give no information on whether the non-verbal mimicry was conventionalized, that is, would be produced in roughly the same way by all knowledgeable community members. Based on recurrent similarities between the Aguaruna and Huambisa imitations, and between these and those of other Jivaroan languages, Berlin and O'Neill (1981:260, n 1) speculate that the onomatopoeic imitations were indeed conventionalized in communities throughout the region.
It is too early to propose that any of the three birdsong imitation strategies is universally conventionalized. Thus far, no community has been identified in which conventionalized non-verbal vocal mimicry and onomatopoeia but not warblish are used. For instance, Berlin and O'Neill (1981) do not mention Aguaruna or Huambisa warblish, but Aguaruna speakers alerted linguist Simon Overall (personal communication, 2016) to at least one warblish example in their language. The following three sections introduce what we know about the birdsong imitation methods in more depth.
Non-Verbal Vocal Mimicry
Non-verbal vocal mimicry of bird calls may be more utilitarian than onomatopoeia or warblish. In Anglophone North America, non-verbal birdsong mimicry is practiced by hunters and birdwatchers to elicit avian reactions (and at least one California high school holds an annual birdsong mimicry contest; Nicole Sault, personal communication, 2016). As Clancy (1996:55) puts it, “calling is the fun part of turkey hunting…. learning when to yelp, cluck, purr or cutt… this is the art of spring turkey hunting.” As with other areas of folk performance, practitioners may hone their non-verbal birdsong imitations over years.
Since non-verbal vocal mimicry aims to approximate the sound of birdsongs as closely as possible, the degree to which this mimicry is conventionalized—produced in a particular way by all members of a community—may be difficult to establish. It could be surmised that this type of mimicry is not bound by the particularities of individual communities, and depends purely on individual talent. But it is likely that non-verbal vocal mimicry involves a learned, community-specific component, so that members of a single community consistently produce the same combinations of “whistling, humming, grunts, hissing, smacking, or clicking” (Berlin and O'Neill 1981:239) for the same birdsong.
Non-verbal birdsong mimicry across cultures can be supplemented by use of the hands or artificial devices. Berlin and O'Neill (1981:239) write that Aguaruna and Huambisa non-verbal mimicry “may be accomplished at times with the aid of double-cupped hands, appropriately held leaves, or by animal bone whistles.” North American turkey (Meleagris gallopavo) hunters can purchase or craft artificial calling devices; Clancy (1996:55) writes that “some hand-made wooden box calls [sound-making devices] were so good they were passed down through many generations.”
Areas for further investigation of non-verbal birdsong mimicry across cultures include: the contexts in which it is learned; the degree to which it varies within a community and between communities; and the acoustic and auditory relationships between birdsong and its non-verbal mimicry. In comparing this imitation method with the other two, it may be hypothesized that non-verbal vocal mimicry could be preferred when actually eliciting responses from birds, and onomatopoeia or warblish when evoking a bird call in conversation with other humans, but this is yet to be examined for any culture.
Onomatopoeia
The strategy of onomatopoeic imitation is the most commonly mentioned in ethno-ornithological studies—largely because onomatopoeic call imitations often give rise to bird names. In this strategy, people form nonsense words that may combine into set expressions evoking bird calls according to the grammars of their own languages. These are revealing of phonology and acoustic perception and sometimes also help expose ethno-taxonomical affiliations, as with the Nage category of birds and other beings that “say po” (Forth 1998, 2004:63).
Onomatopoeic birdsong imitations generally belong to a special part of speech known as “expressives,” which also includes the English dog's bowwow, rooster's cock-a-doodle-doo, and cat's meow (Agnihotri and Si [2012:203] call these “ideophones” for Solega, but this is not an appropriate term in every language; see Voeltz and Kilian-Hatz [2001]). Often, birdsong imitations contain more than one prosodic word, as in the English onomatopoeia for the Bell's vireo (Vireo bellii), cheedle chee chee cheedle chew (Feith 2002), but since none of the components has independent meaning or function, such imitations merit only a single dictionary entry. Agnihotri and Si (2012:203) term onomatopoeic birdsong imitations “lexicalized bird calls.”
As observers of language have noted repeatedly (see sources in Bredin 1996), onomatopoeia are hardly straightforward acoustic reproductions. They are constrained by the sound system of a language and by convention. For instance, in English, a heavy object's collision with the ground could be described as making the sound thud (IPA ['θ∧]). In Nungon (Papua New Guinea), the same situation creates a sound onomatopoeically rendered as hundum (IPA ['hun.dum]; Sarvasy 2017). Although they both contain a voiced alveolar oral stop “d,” the English and Nungon words otherwise differ in consonant and vowel quality and in number of syllables. Two of the sounds in the English word would never occur in any Nungon word; Nungon does not employ interdental fricatives such as English “th” (IPA [θ]) for contrasts in meaning, nor does it systematically use an unrounded mid-back vowel as in thud (IPA [∧]). Further, while Nungon does have a voiced alveolar stop “d” as in English, Nungon words cannot end with this sound.
Onomatopoeic birdsong imitations are likewise limited by sound systems of particular human languages. In the Solega language of southern India, several consonants are produced as retroflexes, with the tongue tip curled backward in the mouth. The Solega term for “steep slope” is 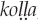 , with retroflex “l” (Si and Agnihotri 2014:390). Solega also includes contrastive vowel nasalization (English has nasalized vowels before nasal consonants, but because their distribution is predictable, English speakers need no awareness of them). Accordingly, the Solega onomatopoeia for the Red-whiskered bulbul (Pycnonotus jocosus) call is
, with retroflex “l” (Si and Agnihotri 2014:390). Solega also includes contrastive vowel nasalization (English has nasalized vowels before nasal consonants, but because their distribution is predictable, English speakers need no awareness of them). Accordingly, the Solega onomatopoeia for the Red-whiskered bulbul (Pycnonotus jocosus) call is 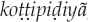 , with retroflex “t” and “d” and nasalized “a” (Agnihotri and Si 2012:203). In Kaluli, the bird named bas is said to call “bas bas bas bas” (Feld 1982:72); in Nungon, “s” cannot occur at the end of any word, including onomatopoeia for birdsongs (Sarvasy 2017). Unlike English, Nungon allows a velar nasal, written “ng,” at the beginning of words, so that the Nungon onomatopoeia for the Rufous woodcock (Scolopax saturata) call is biuin ngok ngok ngok ngok. English speakers do learn to produce this sound, as at the end of bring, but not at the beginning of words; English birdsong onomatopoeia as compiled in Feith (2002) and elsewhere never begin with a velar nasal.
, with retroflex “t” and “d” and nasalized “a” (Agnihotri and Si 2012:203). In Kaluli, the bird named bas is said to call “bas bas bas bas” (Feld 1982:72); in Nungon, “s” cannot occur at the end of any word, including onomatopoeia for birdsongs (Sarvasy 2017). Unlike English, Nungon allows a velar nasal, written “ng,” at the beginning of words, so that the Nungon onomatopoeia for the Rufous woodcock (Scolopax saturata) call is biuin ngok ngok ngok ngok. English speakers do learn to produce this sound, as at the end of bring, but not at the beginning of words; English birdsong onomatopoeia as compiled in Feith (2002) and elsewhere never begin with a velar nasal.
Although onomatopoeic birdsong imitations largely fit phonological systems, this does not preclude the possibility that birdsong imitations and other expressives may feature special systematic uses of sound that are not permitted in other types of words. For instance, in Nungon, a word-initial consonant cluster “kr” or “br” only occurs in expressives such as kruk kruk “rustling” and onomatopoeic birdsong imitations such as fit kum brit! for the call of the Chestnut-breasted cuckoo (Cacomantis castaneiventris).
Across languages, onomatopoeic and other renderings of birdsongs differ significantly beyond the dictates of sound systems. For example, the usual conventional imitation in English for the call of the Common poorwill (Phalaenoptilus nuttallii) is warblish: poor Will! poor Will! poor Will! The Sahaptin (Columbia River basin) onomatopoeia for the same call, wawiyúk'k, differs in many sounds from the English warblish (Hunn 1991:141). Different sound makeups of imitations across languages do not mean, however, that the consonants and vowels in a particular imitation are entirely arbitrary.
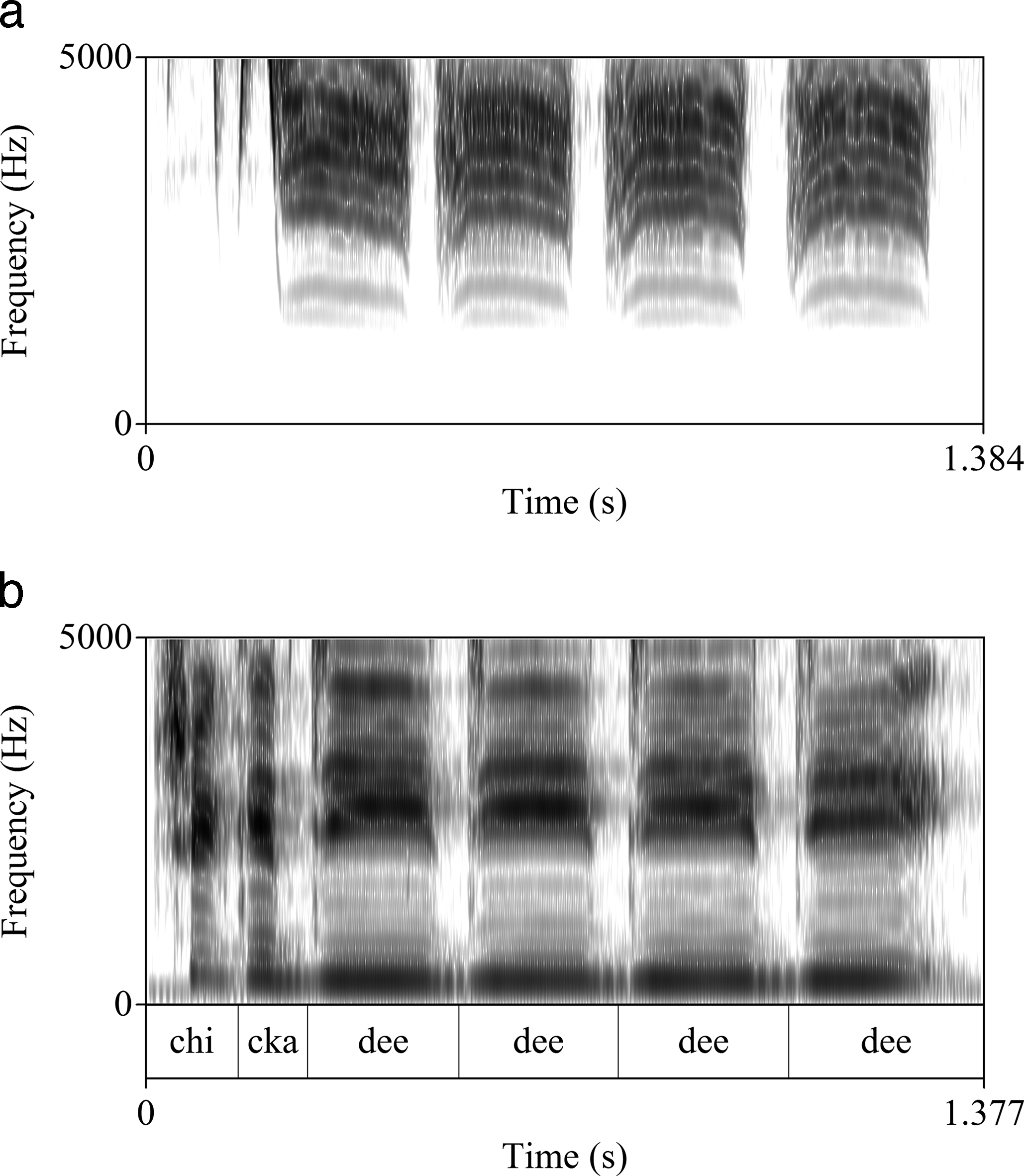
Indeed, acoustics of birdsongs may influence humans' choices of consonants and vowels in onomatopoeic renditions, although other characteristics of sound not usually represented in writing, such as tempo and pitch, may be crucial to effective imitations. Figure 1a is a spectrogram of one of the Black-capped chickadee's calls (Poecile atricapillus). Figure 1b is a spectrogram of a female human rendition of the English onomatopoeia for this call, chickadee dee dee dee. Spectrograms represent sound visually, with time on the x axis and frequencies in Hertz on the y axis. Concentrations of acoustic energy show up as dark bands. All spectrograms here were created using the software PRAAT (Boersma and Weenink 2013).
Black-capped chickadee call (audio available at: http://www.xeno-canto.org/334996).
In Figure 1b, the recurring dark areas in the lowest frequencies absent from Figure 1a are due to vibration of the human larynx. In the birdsong in Figure 1a, the initial quick bursts of energy in the higher frequencies are similar to those of human fricative consonants as in sssh! This is approximated with the first consonant of the onomatopoeic rendering in Figure 1b. The vowel repeated four times in chickadee-dee-dee-dee, IPA [i:], is the English vowel that most closely approximates the avian song, with bands of energy (formants) close together, and concentrated in the higher frequencies. Spectrograms of other English vowels show larger gaps between formants and formants in the lower frequencies. Here, there seems to be acoustic motivation for the choice of two consonant and vowel sounds in the English onomatopoeia. But the other sounds may be relatively arbitrary.
The most important aspects of onomatopoeic imitations are likely relative timing, tempo, intonation, and voice quality, though these are not consistently noted in ethno-ornithological studies. For instance, spectrograms 1a and 1b also show that the relative timing of the components of the human imitation closely mimic those of the birdsong. The first two bursts of energy are short in duration compared to the following four in both Figure 1a and Figure 1b. This may be more critical to a successful onomatopoeic rendition than choices of vowels and consonants.
It is common around the world for bird names to derive from onomatopoeic birdsong imitations. Often, an alternative folk taxonomy based on vocalization type distinguishes such birds as “saying their names,” as with the Kaluli and Nungon speakers in Papua New Guinea (Feld 1982; Sarvasy 2017) and apparently among the Samia of Kenya (Ng'weno 2010:107, 111). The onomatopoeic imitation is frequently longer than the derived name (Berlin and O'Neill 1981; Feld 1982:72–73). The imitation sometimes includes several repetitions of a unit repeated only once or twice in the name, as with the quail named 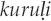 by the Solega, for which the onomatopoeic call imitation is
by the Solega, for which the onomatopoeic call imitation is 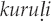
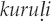
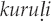 (Agnihotri and Si 2012:192). Alternatively, the imitation may include additional nonce onomatopoeic material missing from the name, as with the Huambisa onomatopoeic imitation kurukupipipip ɨkantsamkantsam for the Grayish mourner (Rhytipterna simplex), whose Huambisa name, ukúrpip, derives from part of the imitation (Berlin and O'Neill 1981:252).
(Agnihotri and Si 2012:192). Alternatively, the imitation may include additional nonce onomatopoeic material missing from the name, as with the Huambisa onomatopoeic imitation kurukupipipip ɨkantsamkantsam for the Grayish mourner (Rhytipterna simplex), whose Huambisa name, ukúrpip, derives from part of the imitation (Berlin and O'Neill 1981:252).
A bird name listed as “onomatopoeic” in the literature may alternatively derive from onomatopoeic renderings of other sounds associated with the bird, such as the beating of its wings, as in Sahaptin (Hunn 1991:141) and Tlingit (Alaska; Hunn and Thornton 2010:188). An onomatopoeic imitation of wingbeats may coexist with onomatopoeic imitations of calls for the same bird; this is the case for the Stephanie's bird-of-paradise (Astrapia stephaniae) among the Kalam (Papua New Guinea; Majnep and Bulmer 1977:60–61). This bird is named ksks for “the sound of the swish of the wings as it flies,” while two onomatopoeic imitations also exist for its calls: ŋe ŋe ŋe and ss ss ssw ssw.
Berlin and O'Neill's speculation that “non-literate peoples” may use more fauna names derived from onomatopoeic call imitation than those with “complex literate traditions” (1981:259) is as yet unproven. In existing studies, percentages of bird names acknowledged to derive from onomatopoeic call or wingbeat imitations range from just over 7% for the Southern Luri-speaking nomadic Mamasani community of Iran (Anonby 2006:10–18) to 49% for Tzeltal Maya (Berlin and O'Neill 1981:259).
To date, onomatopoeic birdsong imitations have only been systematically examined in the service of claims about the origins of names. But they merit rigorous investigation in their own right. The extent to which they vary formally within and across speech communities and the process through which they are learned are two areas for descriptive research. The question of how much phonology and existing onomatopoeic conventions within one language determine new onomatopoeic forms begs an intriguing experiment that could have global scale. First, present unfamiliar bird calls to individuals from a single speech community to elicit nonce onomatopoeic imitations, then compare these. Then present the same bird calls to people in other speech communities around the world and study how nonce onomatopoeic imitations differ within a community and across communities worldwide. Finally, it is as yet unknown how cultural conventions for wordless singing, such as English la la la, dee dee dee, or dum dum dum relate to onomatopoeic birdsong imitation conventions. Onomatopoeia lack further semantic and pragmatic grounding because they have no intrinsic meaning beyond evocation of a sound. It is the third strategy for birdsong mimicry—warblish—which is revealing of culture beyond the level of sound.
Warblish
There has been to date no established term for the phenomenon of humans setting existing words to birdsong. Those noting it have either fumbled for terminology or used a term that does not apply across cultures. In describing warblish in Arandic languages (Australia), Turpin et al. (2013:19, 26) write of a call “saying the thing it signifies” and being “anthropomorphized into the local language as saying something specific.” North American English-speaking birdwatchers often call warblish “mnemonics” (e.g., Feith 2002). But in only a minority of cultures around the world does it seem that warblish is just an aid for learning to recognize calls. The most accurate description of warblish, “verbal imitations of birdsong,” is attested in a presentation by Donegan and Stampe (1977; Patricia Donegan, personal communication, 2016), the full content of which is lost to the annals of pre-digital age conferences. North American English warblish includes those given in Table 1.
North American English warblish examples.

As with onomatopoeia, the consonants and vowels of warblish may be particularly suited to imitate the bird call. Figure 2a shows a spectrogram of the Whip-poor-will call (Antrostomus vociferus), while Figure 2b is a spectrogram of a female human rendition of the English warblish for its call, whip poor Will! In Figure 2b, the transition from [w] to [
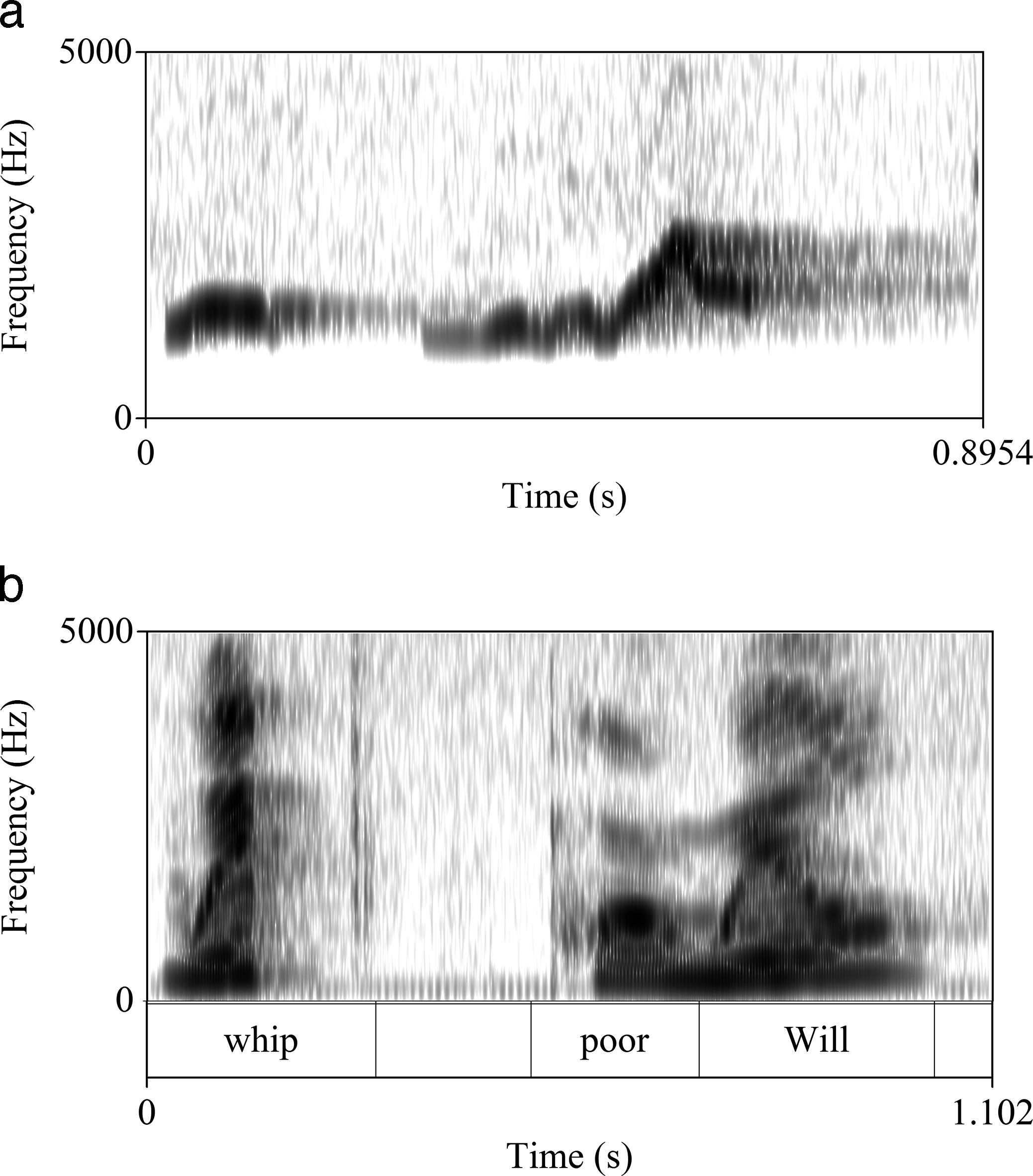
Whip-poor-will call (audio available at http://www.xeno-canto.org/325433).
The warblish for the White-throated sparrow (Zonotrichia albicollis) varies in North American English from poor Sam Peabody, Peabody, Peabody to oh sweet Canada, Canada, Canada. Apart from the diphthong [oυ] in the first syllable of both, these two variants differ in every vowel; [æ] in Sam versus [i:] in sweet and the sequence [i:  i] in Peabody versus [æ ə Λ] in Canada. The avian song is in Figure 3a, and the two English warblish variants are rendered by a female speaker in Figures 3b and 3c.
i] in Peabody versus [æ ə Λ] in Canada. The avian song is in Figure 3a, and the two English warblish variants are rendered by a female speaker in Figures 3b and 3c.
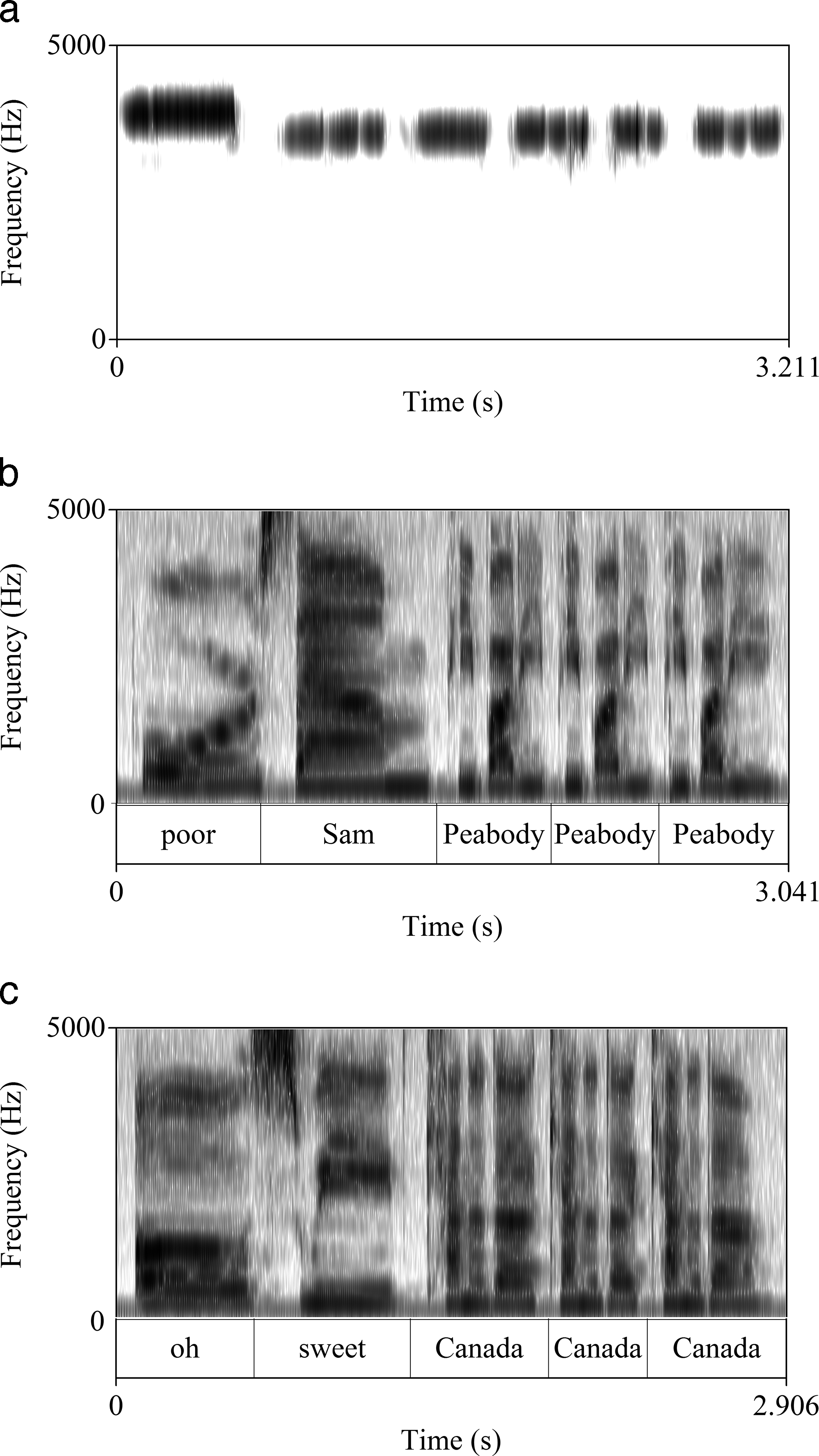
White-throated sparrow call (audio available at: http://www.xeno-canto.org/329835).
Of note in Figure 3a are the descending pitch, the number of notes, and their timing. The White-throated sparrow call lacks swoops in Hertz like those in the Whip-poor-will call of Figure 2a. Thus, consonant and vowel qualities in any imitation are less crucial to fit than number of syllables and their timing and descending pitch. Hence, the thrice-repeated trisyllabic word in the warblish of both Figure 3b and Figure 3c, which would seem to equally resemble Figure 3a, despite largely differing from each other.
Just as warblish can vary within a community, the sound composition and meaning of warblish in different languages for the same call can vary greatly. The English warblish for the Great horned owl (Bubo virginianus) call is who's awake? me, too, while the Sahaptin warblish for the same call is the ominous pátkwatana tanínšin, “Arrowhead has eaten,” followed by a person's name (Hunn 1991:140). The Western meadowlark (Sturnella neglecta) song lacks either conventionalized warblish or onomatopoeic imitation in English (Feith 2002), but Sahaptin and Secwepemc (British Columbia) speakers attribute warblish to the bird; it issues “taunting phrases” in Sahaptin (Hunn 1991:143) and advises on planting biscuit root in Secwepemc (Ignace and Ignace 2015). Variation in warblish around the world reveals differences in the ways communities have framed their relationship with birds. Examples in the rest of this section show that birds can figure in this relationship as heralds or transformed mythical humans. Alternatively, warblish depicts birds as chattering about their avian routines. Facetiously irrelevant and incongruous warblish, like the Northern waterthrush's nice old ladies don't chew in Table 1, may be more typical of industrialized societies and those with a history of colonization.
Some birds are known to call on the approach of a person or animal and these calls may generate appropriate warblish. Eastern Central Arrernte speakers (Australia) give the warblish for the Striated pardalote (Pardalotus striatus) as ipenye-petyeme “stranger coming” (Turpin et al. 2013:19). Similar warblish is attributed to the Red-browed pardalote (Pardalotus rubricatus) and Grey fantail (Rhipidura albiscapa) elsewhere in Australia (Tidemann and Whiteside 2010:170; Turpin et al. 2013:26). Warblish may indicate whether the approaching human is harmful; two Kaluli warblish for Chanting scrubwren (Crateroscelis murina) calls are kalu yab∊, “a person is coming,” and sei yab∊, “a witch is coming” (Feld 1982:79).
If the bird's call heralds the approach of an animal, its warblish may be directed at a hunter or the animal's owner. The Kalam warblish for two of three calls of the kosp (Mountain mouse-warbler, Crateroscelis robusta) are tip-offs for hunters: ñ-o py-ok-ey py-ok-ey and variants translate as, “son, something is close by” and tell a hunter that game is near, ñ-o yj pls gp-ey, ñ-o pls gp-ey, “son, now something is caught,” alerts the hunter that an animal is caught in a trap nearby (Majnep and Bulmer 1977:95). The Black-throated warbler (Gerygone palpebrosa) may help Kaluli locate stray pigs with warblish announcements of their whereabouts (Feld 1982:79).
Seasonal changes in birdsong mean that warblish often heralds planting and harvesting times. Meru and Gikuyu are closely related Bantu languages of Kenya. According to Muiruri and Maundu (2010:287), Meru speakers cite the warblish of the Red-chested cuckoo (Cuculus solitarius) as mbaikia ku, “Help me put the load of firewood onto my back [to take home],” while Gikuyu speakers cite it as ngwikia ku, “Where do I sow the seed?” Both of these relate to the cultural function of this call to alert people to sow seeds and to stockpile firewood for the rainy season. The Kalam warblish for the mature Schlegel's whistler call (Pachycephala schlegelii) is tw pc pc, “axe chop chop,” which Majnep and Bulmer (1977:84) report alerts people to the season of clearing new farm plots.
The North American English warblish for the breeding song of the Bobolink (Dolichonyx oryzivorus) originally had a similar function in folk agriculture, but the warblish appears to have been forgotten with a decline in small-scale agriculture. Hill (2000:116) writes of his 1930s childhood in rural Vermont, “we all knew that the only good time to plant corn was the week that the bobolinks returned and sang their permission.” This permission was issued as warblish, Bobby Lincoln-Lincoln time to plant corn, if you plant any this year, this year, this year (Goodrich 1849:183). The Bobolink's warblish has vanished from twenty-first century birdwatchers' repertoires; Feith (2002) only describes the Bobolink call as “long and bubbling,” and internet mentions of the Bobolink call similarly shy from either warblish or onomatopoeia.
Warblish sometimes refers to observable avian behavior. The Kalam warblish for the calls of the Black fantail (Rhipidura atra) and Dimorphic fantail (Rhipidura brachyrhyncha) relates to the birds' habits: warblish joley-boley is translated by Majnep and Bulmer (1977:85) as, “light in one place, alight in another, call and call again.” The Kaluli say that their warblish for the White-crowned koel (Cacomantis leucolophus), n∊ f∊s  “my back hurts,” stems from “the pain it feels from not sitting down, just flying around constantly” (Feld 1982:79). Among the Kalam, ttmñ, the Friendly fantail (Rhipidura albolimbata), is said to dance. Men chant its warblish before dancing (Majnep and Bulmer 1977:85).
“my back hurts,” stems from “the pain it feels from not sitting down, just flying around constantly” (Feld 1982:79). Among the Kalam, ttmñ, the Friendly fantail (Rhipidura albolimbata), is said to dance. Men chant its warblish before dancing (Majnep and Bulmer 1977:85).
Warblish frequently references a traditional story. The story is often about a person who turns into a particular bird. The content of that bird's warblish is then usually a lament about the circumstances that led to the shape-shift. This is the case for the Black cuckoo call (Cuculus clamosus) among the Samia (Ng'weno 2010:107). Similarly, in Tariana (Brazil), one legend relates that a group of children whose mother scolded them turned into a particular type of guan. To this day, these birds are said to call in the neighboring East Tucanoan language, Guanano, “Our mother scolded us!” (Aikhenvald 2003:660). The Kaluli tell a story in which children who had gone to fetch water were killed by enemies (Feld 1982:76–77). Their voices were then taken by the bird 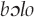 , noted earlier to encompass both the New Guinea friarbird and Brown oriole because of their vocalizations. Among the Kaluli warblish for
, noted earlier to encompass both the New Guinea friarbird and Brown oriole because of their vocalizations. Among the Kaluli warblish for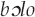 calls is the mournful dowo,
calls is the mournful dowo, 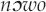 “father, mother” that relates to this story (unrelated, ribald warblish are also attributed to the
“father, mother” that relates to this story (unrelated, ribald warblish are also attributed to the 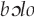 ; see below).
; see below).
At times the story and warblish may be better known than the bird's name. Agnihotri and Si (2012) write that all their local consultants on Solega ethno-ornithology agreed on the warblish of the Puff-throated babbler (Pellorneum ruficeps), but many could not give a name for the bird. The warblish nanna ku:so!, “my child!”, is related to “a universally-known folk tale in Solega that explains why this bird came to lament a lost child” (Agnihotri and Si 2012).
It is usually unclear whether the legend into which a warblish phrase fits preceded the warblish or vice versa. A myth could hypothetically arise to explain warblish; perhaps, in time, North American English speakers will tell the legend of the California quail yearning for its hometown in Illinois. In single-authored literature, on the other hand, the direction of development is usually plain. Robert Merry's Museum, a nineteenth-century U.S. children's periodical, ran fanciful and factual tales about fauna. In one story, the Song sparrow (Melospiza melodia) character is constructed around its English warblish:
The cheerful Tea-kettle Birds…were a strange family, always wanting tea. Hear them when you would, old or young, they were constantly calling out, “Maid, maid, put on the tea-kettle, kettle, kettle, 'tle, 'tle, 'tle.” (Goodrich 1849:183)
In multilingual societies in Papua New Guinea and Amazonia, warblish may feature foreign languages. Sometimes this is because the bird is associated with the region where the other language is spoken, as with the Kalam warblish for the bird called ñyolelegp, which means “a man is dying” in the Kobon language, spoken where the bird is mainly found (Majnep and Bulmer 1977:103). Two out of four known instances of warblish in Nungon include material from Nukna, the language of Nungon speakers' former trading partners. Either using a foreign language has magical or other significance, or Nungon speakers borrowed Nukna warblish, perhaps as an avian community expanded into the Nungon area. Tariana speakers pride themselves on never speaking in other Amazonian languages, even if they understand them. But in legends and warblish, birds, animals, and evil spirits often speak these other languages (Alexandra Aikhenvald, personal communication, 2015).
Some warblish is like the result of homophonic translation between human languages in that it seems to be an irrelevant string of words. Homophonic translation (see Dembeck 2015) disregards semantics in service of auditory resemblance to the original. This semantic disconnect has a humorous effect. The famous French homophonic translation of English poem Humpty Dumpty is usually attributed to van Rooten (1967); its first line, meant to approximate the sound of Humpty Dumpty sat on a wall, is often given as Un petit d'un petit s'etonne aux Halles, “a small one of a small one was astonished at the market.” Likewise, I have observed Mexican birthday celebrants serenading the guest of honor with a Spanish homophonic translation of the English birthday song, beginning Sapo verde eres tú, “a green toad are you.”
Just as homophonic translations are usually humorous, warblish can be humorously incongruous with the birds to which it is attributed. This is attested much more for North American and Australian English than for other languages referenced here. North American and Australian English warblish are rife with flippant warblish, such as several referring to “beer” in the United States (Feith 2002) and examples like shit a brick! (White-plumed honeyeater, Lichenostomus penicillatus) in Australia. Some North American English warblish is popularly attributed to individual field guide authors or ornithologists. Twentieth-century ornithologist Arthur Allen is credited with the English warblish for the Eastern warbling vireo (Vireo gilvus) song; this warblish is known in two variants with more or less veiled reference to male (human) genitalia.
It may be the case that facetious warblish is most prevalent in industrialized societies and those with shorter shared histories with avian communities due to recent colonization. Andrew Gosler (personal communication, 2015) surmises that flippant warblish in England was a by-product of the Protestant Reformation and concomitant movements toward rationalist, scientific thought, away from augury. Citing nonsensical rhymes attributed to birds in nineteenth-century England, he suggests that both North American and Australian English flippant warblish could stem from this tradition. This effect could have been compounded by the white American and Australians' lack of intimacy or historical associations with local wild birds. Humor and off-color warblish are not limited to these societies, however. Hunn (1991:141) implies that the Killdeer (Charadrius vociferus) calls tiit “fart” in Sahaptin. The Kaluli attribute ribald warblish to the male 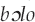 (New Guinea friarbird, Philemon novaeguineae); men heckled with these insults about their genitalia “shout back, with both laughter and scorn” (Feld 1982:78).
(New Guinea friarbird, Philemon novaeguineae); men heckled with these insults about their genitalia “shout back, with both laughter and scorn” (Feld 1982:78).
Warblish is not necessarily conventionalized within a community. A bird may be interpreted to say different things depending on the situation and the human interpreter. Nungon speaker Rosarin Ögate (personal communication, 2012) has her own warblish interpretations for the call of a bird she claims helps her keep time while she is in her remote farm plots, singing nain öklak! at 9:00 am, teri öklak! at 3:00 pm, and sis öklak! at 6:00 pm. Some birdwatchers in Washington State dispute the generally publicized English warblish for the Red-winged blackbird (Agelaius phoeniceus), konklaree! (Feith 2002). On the Whatcom Birds email list in Washington State, one birdwatcher wrote, “I think our blackbirds sing ma-ma-MEE-ah,” while another declared, “I always thought they were singing Mt. Ray-NEE-er. At least the Washington ones were!” (Sheila Sondik, personal communication, 2015).
Warblish of a bird with soothsaying powers can be expected to vary depending on portent. Forth (2004:99) writes of an investigation of a death among the Nage that when the dead party's mistress's name was uttered, a Helmeted friarbird (Philemon buceroides) “flew across the village calling iko ako ma ke ma ke ma ke, ‘iko ako, ‘that's it, that's it, that's it.'” Other versions of this account use other warblish, ta kéna de, ta kéna de, “that's the one, that's the one,” or ta bu'e, ta bu'e, “the mistress, the mistress” (Forth 2004:214, n 4). The Sahaptin warblish for the Great horned owl call noted above is followed by the name of a person who is doomed to die (Hunn 1991:140).
Conventionalized call-and-response exchanges between people and birds may involve warblish. This area of ethno-ornithology is rarely reported but could nevertheless be widespread. Humans sometimes address birds with human language to elicit non-vocal responses. Nungon-speaking children call to Raggiana birds-of-paradise (Paradisaea raggiana): mak! nok eppot! “Mother! I'm coming!”, to draw the birds into view and Tashelhit-speaking children (Morocco) taunt domesticated turkeys with tmut 3aisha Bibi!, “Aisha Turkey has died!”, to elicit gobbling and eventually charging. Alternatively, the bird may respond to formulaic addresses by humans with portentful warblish, as the Pied chat (Saxicola caprata) is said to do among the Kalam (Majnep and Bulmer 1977:100–102)
Warblish is a little-studied area of human creativity in response to nature. Its relevance to existing fields of inquiry is multifold. Warblish can include clues to cultural knowledge of avian behavior and can also reveal elements of human cultures unrelated to birds. For historians, the presence of words from other languages in warblish may support claims of historical connections between human groups.
Conclusion
People around the world imitate avian vocalizations in three main ways: non-verbal vocal mimicry, onomatopoeia, and warblish. This paper outlines the three birdsong imitation methods and provides a new term, warblish, for vocal imitation using existing words, which was heretofore unnamed. It suggests further investigatory directions for each method, within and across human speech communities worldwide. It proposes that conventions for imitating birdsong may encode cultural knowledge of birds that goes beyond taxonomies based on morphology.
This cultural knowledge may involve alternative folk taxonomies based on avian vocalization type, such as those of the Kaluli (Feld 1982) and Nage (one such category is described by Forth [1998, 2004]). Such taxonomies of sound could be widespread, albeit little-studied, since a bird's call can be more salient to humans than its appearance. Other types of cultural knowledge of birds can be encoded in warblish; these include correlations between birdsong and seasons, patterns of avian flight, and some birds' tendencies to announce other animals' movements. Warblish can also highlight traditional conceptions of birds as transmogrified humans. North American and Australian English speaking birdwatchers belong to societies that scoff at augury and no longer practice small-scale agriculture; their contemporary warblish encodes reference to cultural mores unrelated to ethnobiology. The facetious tone of much warblish in these cultures differs from the tone of warblish in small communities like the Kalam of Papua New Guinea or Samia of Kenya.
Most existing ethno-ornithological literature discusses birdsong imitations haphazardly. An emphasis on bird names and their mapping to Linnean equivalents has meant that other ways humans describe, evoke, and interact with birds are buried unsystematically in prose following a bird's name. Even where onomatopoeic birdsong imitations are systematically examined, as in Berlin and O'Neill (1981) and to a lesser extent in Agnihotri and Si (2012), the questions of degree of conventionalization and amount of variation within a community are not pursued.
The season is ripe for ethno-ornithologists to catalogue, experiment with, and analyze conventionalized birdsong imitations in societies around the world. To quote the Kalam warblish for the Schlegel's whistler call, “axe chop chop!” Let's begin clearing this fertile ground now.
Footnotes
Acknowledgments
Sincere thanks to two anonymous reviewers, Peter Reynolds, Alan Rumsey, Nicole Sault, and Journal of Ethnobiology editors Dana Lepofsky and Steve Wolverton for helpful comments on earlier versions. Thanks to Jared Diamond for sage advice and encouragement in coining the new term. Data and suggestions were contributed by: Alexandra Aikhenvald, George Appell, Wendy Baarda, Margit Bowler, R. M. W. Dixon, Nicholas Evans, Eleanor Glewwe, Greta Hawes, Bruce Hayes, Pam Munro, David Ögate, Irising Ögate, Rosarin Ögate, Simon Overall, Jane Simpson, Brian Smith, Sheila Sondik and audiences at the Society of Ethnobiology 2015 Meeting, the 2015 Western Folklore Society Meeting, a UCLA American Indian Seminar, and the Centre of Excellence for the Dynamics of Language at the Australian National University. Members of the National Puzzlers' League e-mailed inventive nomenclature ideas; it was Tahnan's passeringlish that inspired my warblish.