
Abstract
Direct evidence for the use of geophytes in the archaeological record of prehistoric foragers is typically lacking, even though they were often an important resource. Cymopterus bulbosus (springparsley or biscuitroot) is a geophyte that grows in fairly dense patches in areas of high archaeological site densities in the Green River Basin of Wyoming. Analysis of the patch size and distribution, density within patches, root weight, nutritional content, total calories available from the patches, and caloric return rates provides clues to the sustainability of roots of this plant for the prehistoric foragers of the area. This study demonstrates that the large desert pavement patches adjacent to the archaeological sites had the potential to furnish sufficient calories to have been an important influence on prehistoric forager selection of camp locations. It also adds to the growing corpus of information on the economic context of root resources and their use by prehistoric foragers.
Introduction
Geophytes (plants with underground storage organs) have served as a significant food resource and often as a staple for peoples throughout the world. Recent hypotheses concerning the evolution of Homo erectus suggest that geophytes played a major role in the human evolutionary process (O'Connell et al. 1999). Many early and modern foragers obtained a large portion of their carbohydrates from edible roots and tubers (Vincent 1985). A number of geophyte species in the Old and New Worlds were also domesticated, including at least 16 species in the American tropics (Alexander and Coursey 1969; Harris 1969; Hawkes 1989). In the northwestern United States and western Canada, geophytes provided a predictable, stable, and storable food resource, allowing prehistoric and ethnographic peoples to adopt a sedentary settlement pattern during at least some seasons (Ames and Marshall 1980; Peacock 1998; Prouty 1995; Thoms 1989). Despite this worldwide importance, geophytes are often overlooked in discussions of past plant utilization patterns, especially when compared to seeds (Bates 1985).
Lack of attention to geophytes is partly due to the typical absence of direct evidence for their use in the archaeological record. Though pieces of charred geophytes are occasionally recovered from excavated cooking or storage facilities (Prouty 1995:201–212; Thoms 1989:394), this is generally not the case in most contexts. Stone tools used in geophyte exploitation, such as large bifaces and groundstone, are preserved in archaeological contexts, but were also employed for other tasks, thereby reducing their usefulness as a marker for the procurement and processing of geophytes. The most important artifact associated with root and tuber digging, the digging stick, is only preserved under special conditions. In the absence of direct evidence, many archaeologists use the presence of pit ovens or storage facilities as a proxy for the importance of root and tuber collecting and processing (Francis 2000; Peacock 1998; Prouty 1995; Smith et al. 2001; Thoms 1989; Wandsnider 1999). Not all geophytes, however, were baked in pit ovens or stored, so in some cases the prehistoric use of these resources would not be evident in the archaeological record based on these proxies.
Considering other forms of evidence highlights the role that geophytes might have played in the subsistence behaviors of prehistoric foragers. The kinds of plant resources in the foraging area adjacent to an archaeological site may furnish clues, though the presence of an edible plant species in the vicinity of a site does not necessarily demonstrate its importance in foragers’ subsistence and settlement decisions. Researchers need to consider a number of factors when evaluating a resource's possible prehistoric use, including caloric return rates, abundance, seasonal availability, predictability, nutritional value, processing requirements, and patch structure (Todt and Hannon 1998; Wandsnider and Chung 2003).
Recently, a number of studies have taken new approaches to interpreting the significance of geophytes in the diet of prehistoric foragers. These approaches include determining the post-encounter return rates for some geophyte species through experimental and ethnographic studies (Couture et al. 1986; Hunsaker and O'Connell 2002; O'Connell et al. 1983; Simms 1984; Smith et al. 2001; Thoms 1989; Vincent 1985); determining complex carbohydrate composition and heat-treatment requirements (Peacock 1998:149–159; Smith et al. 2001; Thoms 1989:157–162; Wandsnider 1997); pit oven experiments (Peacock 1998:149–159; Smith et al. 2001); and studies of patch density and size (Hazlett 1991; Moe and Todd 1983; Prouty 1995; Wandsnider and Chung 2003). These studies have shown that caloric return rates for different geophyte taxa range widely depending on the ease of collection, size of the root or bulb, plant densities, and post-collection processing requirements, though they are all generally lower than for large and small mammals. Despite the often low return rates, a number of taxa probably provided a major portion of the diet and influenced land use patterns, at least during certain seasons, while other species contributed little to the diet, partly because of their low density or small patch size. More information is needed to fully understand the importance of geophytes to prehistoric foragers, especially the size and structure of patches and their relationship to the prehistoric campsites.
This study examines patch size, density within patches, root weight, nutritional content, total calories available from patches, and caloric return rates of Cymopterus bulbosus A. Nels. (springparsley or biscuitroot). Together with Allium textile Nels. & Macbr. (prairie onion), C. bulbosus is the most common geophyte observed during April and May in the open patches adjacent to dunal areas that frequently contain abundant archaeological sites in the Green River Basin of Wyoming (Figure 1). For comparison, the nutritional content and caloric return rates were also analyzed for A. textile, the other major geophyte present in patches near the archaeological sites.

Location of the study area within the Green River Basin, Wyoming.
The Study Area
The Green River Basin is one of several intermountain basins comprising the Wyoming Basin within the Middle Rocky Mountain physiographic province (Fenneman 1931). The climate is semiarid, with long cold, windy winters, cool short summers, and short growing seasons (Martner 1986). The vegetation is primarily sagebrush steppe, with broad areas of mixed desert shrubland growing on dunal deposits and associated alkaline upland surfaces. Riparian vegetation extends along the perennial watercourses (Knight 1994).
Numerous prehistoric campsites cover the dunal areas of large portions of the Green River Basin. Surrounding these dunes are open areas, often expansive, consisting of alluvial terraces and residual desert pavement that support only sparse vegetation, though patches of several plants with edible geophytes grow in these areas during the spring. The sites within the dunes date throughout the Holocene, but occupations were most common 8,500–8,000, 6,000–3,600, and 1,800–1,000 years ago. Archaeological remains typically include basin and pit features with charcoal-stained sediment (probably mostly functioning as general cooking hearths), heat-altered rock, flaked stone tools and debitage, small quantities of animal bones, and occasionally housepits. Generally, each occupation appears to have been of short duration, but the site locations were important as evidenced by their repeated reuse over periods of centuries or even millennia (Smith 2003; Smith and McNees 1999).
Over the past several decades, much archaeological research has been conducted throughout the area in an attempt to more clearly understand the settlement and subsistence patterns of the prehistoric inhabitants and why such extensive, repeated occupations occur in the relatively low carrying-capacity environment of the Green River Basin. Among the important food resources that might have been available to prehistoric peoples were C. bulbosus roots and A. textile bulbs growing in open areas adjacent to the dunes. These patches may have provided a spatially and temporally predictable resource that may have periodically drawn prehistoric foragers to the site locations in the spring, when these geophytes would have been detectable on the surface. However, direct evidence for collecting and processing of these geophytes is usually lacking in the archaeological record from the area, though charred root material has occasionally been recovered from sites in the Wyoming Basin (Francis 2000; Sender et al. 1982).
Plant Description and Ethnographic Background
Cymopterus bulbosus, a member of the Apiaceae family, is a perennial herb with an elongated and thick taproot and a short stem (Figure 2) (Cronquist et al. 1997:392–394; Harrington 1967). Phylogenetic analyses of molecular and morphological data indicate that the genus Cymopterus is polyphyletic and inextricably linked with other Apiaceae genera such as Lomatium, another genus containing species with thick and elongated taproots (Downie et al. 2002; Gilmartin and Simmons 1987). C. bulbosus leaves form near the base and are fernlike (dissect). Flowers occur in clusters or umbels. They grow at elevations between 1500 and 2000 m asl on dry open areas, are common in sandy or clayey soil, and are tolerant of alkali. Their range includes southwest Wyoming, the Uinta Basin of Utah, western Colorado, northeastern Arizona, New Mexico, and west Texas. The aboveground part of these plants is visible on the landscape generally only in April and May, when they are in flower. In southwest Wyoming, C. bulbosus is common on dry, open, alluvial silt terraces and desert pavement areas. In 2003, they were observed in flower on May 6 (though they were probably visible from early April). They had mostly gone to seed by the first week in June, at which time the aboveground portion began to dry up and disappear, leaving no visible evidence of the plant.

Examples of collected C. bulbosus plants showing elongated and thick taproots.
C. bulbosus is one of several species in the Cymopterus genus common in the Rocky Mountain area that has edible roots. Various Cymopterus species are listed in several ethnobotanical and ethnohistoric sources as a food resource for the ethnographic groups of the Rocky Mountains and intermountain areas (Castetter 1935; Chamberlin 1911; Fowler 1986; French 1971; Havard 1895; Morton 1963). These sources note that roots, leaves, and seeds were used as food, depending on the species. Roots were eaten raw, baked, or boiled and often ground into flour and made into cakes. Smith (1974:271) records that the Ute in the Uinta Basin of Utah would bake biscuitroot (species not identified, could be a Cymopterus or Lomatium) overnight in earth ovens.
Additional information on the possible importance of Cymopterus roots comes from ethnographic data on the use of Lomatium (also known as biscuitroot), a genus closely related to Cymopterus that is available at the same time of the year. Lomatium provided a major source of food for ethnographic groups throughout the western United States, especially in the Northwest where several highly prized species grow (French 1971; Hunn and French 1981). Among the most important species that served as plant staples for many groups were L. canbyi Coult. & Rose and L. cous (S. Wats.) Coult. & Rose. These roots were collected in the spring and were boiled or dried whole or ground and made into cakes. They contributed a significant portion of the caloric consumption of the ethnographically known peoples of the Columbia Plateau (Hunn 1981), allowing many groups to become sedentary at least during some seasons (Ames and Marshall 1980).
Ethnographically recorded groups whose territory crossed the Green River Basin of southwest Wyoming included the Eastern Shoshone and Ute (Murphy and Murphy 1960; Shimkin 1947). By the time ethnographic studies were conducted, their subsistence and mobility were greatly influenced by their possession of the horse. Ethnographic information concerning pre-horse use patterns of the area is limited. Some analogies can be drawn from the ethnographic record of the Great Basin, except that southwest Wyoming lacks an important food resource, pinyon pine (Pinus monophylla Torr. & Frem.). The Shoshone of the northern Great Basin were foragers who moved throughout their territory according to the availability and density of animal and plant resources (Fowler 1986; Steward 1938). Though the entire seasonal round is not well known, the archaeological record from the Green River Basin indicates similar patterns during the later prehistoric period, with subsistence focusing on a wide variety of animals and plants including bison, antelope, rabbits, ground squirrels, voles, sage grouse, tubers, fruits, and seeds (Creasman and Thompson 1997). The men probably hunted large game on an encounter basis while the women concentrated their efforts on collecting plant resources near their camps, as in the Great Basin (Zeanah 2004). Geophytes such as C. bulbosus would have been one of the earliest plant resources available in the spring and would have been especially valuable as a source of essential carbohydrates following the nutritionally stressful late winter period (Speth and Spielmann 1983).
Past Environment and Human Influences
C. bulbosus patches may have been fairly stable throughout much of the Holocene, despite fluctuations in effective moisture during this time. Eckerle and Taddie (2002), using an aridity index which models effective moisture through time with soil distribution maps, have predicted that Cymopterus would have been available throughout the Holocene in the study area. Compared to periods before and after, the middle Holocene in the Green River Basin appears to have been drier, resulting in a reduction of ecological diversity, decreased grass cover, and a decline of up to 95% in grazing potential within dunal areas and a decline of up to 50% in non-dunal areas (Eckerle 1997). This drier period, however, probably did not significantly affect the size and density of C. bulbosus patches. Thus, patches of C. bulbosus would probably have remained available in the Green River Basin even with a reduction of grasses and grazing potential during the drier periods, although these conditions may have affected the timing and length of the growing season and when roots could be harvested (Housley 1994:564–567).
Because of the presence of underground storage organs, geophytes are well adapted to variable climatic conditions, arid environments, and changes in temperatures (Harris 1969). The C. bulbosus patches occur on stable desert pavement surfaces that have persisted relatively unchanged throughout the Holocene. The adjacent sand dune areas have also been fairly stable as attested by the presence of intact archaeological remains within them, some of which date to over 8,500 years ago (e.g., Highland n.d.; Smith et al. 2003). The clayey, cobble-mantled thin soils of the desert pavement areas retain water well and are ideal for the development and growth of underground storage organs during the early spring growing seasons. This water retention allows geophytes to endure even during prolonged dry periods, as Prouty (1995:102–105) found for species of Lomatium in southeast Oregon that are similar to C. bulbosus.
Human activities both past and present probably have affected the densities of C. bulbosus. Plant densities may have been greater for much of the Holocene than at present. Past forager digging and collecting techniques actually may have encouraged the growth of the plants and increased their density. These practices may have included selectively harvesting plants based on size, rotating patches to prevent overexploitation, and repeated digging that would have had the effect of mixing and aerating the soil and providing a seed bed (Anderson 1997; Peacock 1998). Additional studies need to be completed to determine whether these practices were important to the prehistoric foragers of the Green River Basin and to what extent they influenced productivity. Present-day cessation of these traditional manipulations and the introduction of cattle and sheep grazing have probably had a negative impact on geophyte patches.
Investigated Root Patches
Four patches of C. bulbosus and A. textile (Patches 1–4) were examined as part of this study. They are adjacent to sand dunes containing abundant and widely distributed archaeological remains dominated by charcoal-stained basin hearth features, heat-altered rock, flaked stone artifacts, and large and small mammal bone. Patch 1 (location of collection trials and patch size and density studies) is on an area of desert pavement adjacent to an extensive dunal site complex designated as Site 48SW6965. It is at an elevation of 1950 m asl (Figure 3a). The other dunal areas adjacent to the large desert pavement Patch 1 also contain the remains of prehistoric campsites, especially heat-altered rock clusters and scatters. C. bulbosus was present across the entire length and breadth of the desert pavement basin. It occurred on most desert pavement surfaces with looser residual/colluvial deposits and on the shallow silt alluvium in ephemeral wash channels, but was not observed on areas eroded to hardpan or covered with aeolian deposits. Dense areas of A. textile also appeared throughout the basin, and Cymopterus acaulis, another edible taxon with roots smaller than C. bulbosus, was sparsely scattered throughout the basin. The sparse vegetation on the desert pavement also included Erigeron sp. (fleabane), Oenothera caespitosa Nutt. (evening primrose), Sphaeralces coccinea (Nutt.) Rydb. (globe mallow), and Kochia americana Wats. (greenmolly summercypress) (Figure 4).
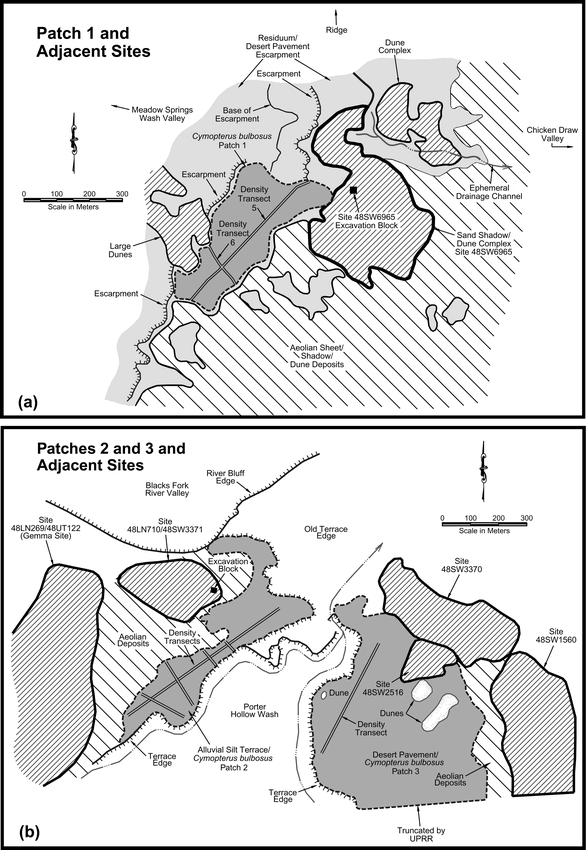
C. bulbosus patches and adjacent archaeological sites: a) Patch 1 near Site 48SW6965; and b) Patches 2 and 3 near Site 48LN710/48SW3371.

Overview of Patch 1 near site 48SW6965 showing desert pavement and sparse vegetation.
Patches 2 and 3 (location of patch size and density studies) are at an elevation of 1920 m asl approximately 19.5 km north of Patch 1. A complex of extensive prehistoric forager campsites in dunal deposits including localities recorded as Sites 48LN710/48SW3371, 48SW3370, 48SW1560, 48SW2516, and 48LN269/48UT122 surround the patches (Figure 3b). Patch 2 is situated on a silt alluvium terrace on the west side of Porter Hollow immediately adjacent to Site 48LN710/48SW3371. Only small, dispersed C. bulbosus plants grew on the terrace, and no A. textile was observed. Atriplex gardneri (Moq.) Dietr. (Gardner saltbush) dominated the other sparse vegetation. Patch 3 is on an extensive desert pavement area on the east side of Porter Hollow wash between dunes containing the Site 48SW3370, 48SW1560, and 48SW2516 complex. C. bulbosus was present throughout the patch and other vegetation was primarily limited to Atriplex gardneri.
Patch 4 (location of a collection trial) is an alluvial terrace patch adjacent to sand deposits containing Site 48SW6962 approximately 2.8 km northwest of Patch 1. The surface of the patch consists of loose silt alluvium in a slight playa basin with scattered desert pavement pebbles. In addition to C. bulbosus, other sparse vegetation included Atriplex gardneri and Erigeron sp. Data recovery excavations have been conducted at Site 48SW6965 (Highland and Batterman 2004; Highland and Byers n.d.) adjacent to Patch 1, Sites 48LN710/48SW3371 (Highland n.d.), 48SW1560 (Hoefer et al. 1996), and 48LN269/48UT122 (Jess 1990; Pastor 1998) adjacent to Patches 2 and 3, and Site 48SW6962 (Highland and Batterman 2004; Pahr n.d.) adjacent to Patch 4. The results of the data recovery investigations indicate that these site localities were repetitively occupied short-term hunter-gatherer camps where general domestic activities occurred. Radiocarbon age estimates span the period from 8,500 to 1,100 years ago. Basin hearth features, heat-altered rock, flaked stone artifacts, and large and small mammal bone dominate the archaeological remains. Little groundstone was recovered, and none of the excavations yielded pit ovens such as would have been used for long-term heat-treatment of geophytes rich in inulin. A charred root fragment was recovered from a basin hearth feature dating to 1,520 years ago at Site 48SW6965, providing possible direct evidence of geophyte use (Bach n.d.).
Methods
Density and Size of Patches
C. bulbosus Patches 1–3 were mapped and their plant density recorded. The perimeter of each patch was recorded using a Trimble GeoExplorer3 GPS unit, which provided the outline and the area of the patch. Straight line transects were then walked through each patch. A point location was recorded with the GPS unit for each C. bulbosus plant encountered within a 1-m wide swath. Using this information, plant density per square meter and total density for each patch was figured.
Two transects (Transects 5 and 6) were walked on 26 May 2003 across Patch 1 (Figure 5a). The density of Patch 2 at Site 48LN710/48SW3371 was recorded on 30 May 2003 using five transects (Transects 1–5) across the silt alluvium terrace (Figure 5b). A single transect (Transect 1) was walked on 1 June 2003 across Patch 3 (see Figure 3b). Only the portion of the patch north of the railroad tracks was examined and recorded. The patch continued south beyond the railroad tracks for a considerable distance.

Examples of mapped perimeter and density transects of C. bulbosus patches: a) Patch 1; and b) Patch 2.
Root Weight
The roots collected from the three C. bulbosus trials in Patches 1 (Collection Samples [CS] 1 and 2) and 4 (CS 4) were weighed to obtain information on their variability in size and differences in root sizes and weights between patches. Immediately after collection, the roots were washed and all dirt adhering to their surface was removed prior to weighing.
Nutritional Composition
For the nutritional composition analyses, fresh, undried samples of C. bulbosus roots and A. textile bulbs were submitted to Medallion Labs, Minneapolis, Minnesota. Analyses included total carbohydrates, sugars, proteins, ash, inulin, total starch measured as percentage, and kcal per 100 g of fresh root. Methods for these calculations are detailed on the Medallion Labs web site (www.medlabs.com).
Total Calories Available from Patches
The total calories from each of the three C. bulbosus patches (Patches 1–3) were figured using the estimated number of plants in each patch multiplied by the average high and low weights for the plant (19.3 [high] and 9.0 [low] g) and then multiplied by the number of kcal per gram (1.18 kcal/g). The total time required to collect the entire patch was also calculated. The estimated number of plants for each patch was divided by the high and low number of plants that could be harvested in an hour based on the collection trials (140 [high] and 52 [low] plants/hr).
Finally, the length of time each patch would support a family of four was computed, assuming 2,000 kcal per person for daily caloric requirements. Obviously, the entire patch would not have been collected during any one season, and C. bulbosus roots would not have been the only food resource utilized during this period. Other food resources including small and large animals were also important as attested by the presence of their remains in the adjacent archaeological sites. However, the total time a patch would support a family was figured to serve as a baseline to evaluate whether C. bulbosus roots could have been a significant enough resource and provided enough kcal to have influenced prehistoric campsite selection.
Return Rates
In our experiments, the return rates for C. bulbosus and A. textile were figured using methods similar to those of Simms (1984). To obtain the quantity of a taxon that could be collected in a given amount of time, collecting trials were conducted. These experiments were completed on 15 and 19 May 2003 when the plants were in bloom at two patches (Patches 1 and 4) adjacent to large, excavated archaeological sites (Sites 48SW6965 and 48SW6962) within the study area.
Three 30-minute collection samples were taken for C. bulbosus. Two individuals each completed one collection trial (CS 1 and 2) at Patch 1 on desert pavement, and one individual conducted another collection (CS 4) at Patch 4 on an alluvial silt terrace. One person also completed a single 30-minute collection (CS 6) of A. textile at Patch 1 for comparison. The roots were dug with a shovel. An experienced collector with a digging stick could probably have obtained the bulbs faster than novices using shovels, so the estimates based on this study may be conservative. Additional collection trials using digging sticks would provide useful comparisons. The experimental collectors targeted medium to large plants and left the smallest plants. The roots from each trial were individually measured, cleaned of dirt, and weighed immediately after the trials. The number of roots dug in the 30-minute time span and hence the resulting return rate varied according to soil moisture. Following a wet snow, the underlying clay was wet and could be easily penetrated with a shovel, facilitating the collection of the entire root with a single direction of prying. As the clay became dry, it hardened, making removal of the roots extremely difficult. The soil contained intermediate moisture at the time of the collection trials, so middle range values for gathering times were likely obtained.
The total weight of the plants collected during each of the 30-minute trials (three C. bulbosus collection trials and the one A. textile trial) was doubled and then multiplied by the kcal per gram (1.18 kcal/g for C. bulbosus and 0.94 kcal/g for A. textile) to obtain the return rate. The return rates were expressed as kcal/hr and only included collection time. Any processing time is not considered in calculating the rates, which would reduce the net return rate, especially if the particular species requires extended baking in pit ovens.
Density and Size of Patches
Plant density per square meter and total density for three investigated patches (Patches 1–3) were figured (Table 1). The average density of C. bulbosus at the two desert pavement patches (Patch 1 at Site 48SW6965 and Patch 3 at Sites 48SW3370, 48SW1560, and 48SW2516) was 0.49 and 0.86 plants/m2, respectively. The patch on the silt alluvium terrace (Patch 2 at Site 48LN710/48SW3371) had a lower density of only 0.06 plants/m2, suggesting that C. bulbosus favors desert pavement contexts over alluvial silt. However, much higher densities (up to 7.09 plants/m2) of small C. bulbosus plants were observed at Patch 4, the silt alluvial terrace patch adjacent to Site 48SW6962. The high density of C. bulbosus plants in Patch 4 is associated with an active veneer of looser deposits where an ephemeral wash crosses the terrace tread. Both the higher density and smaller plant size of the plants appears to be related to the more dynamic depositional context.
Patch size and plant density for the three C. bulbosus patches (Patches 1–3).

The density of C. bulbosus in the Green River Basin of southwest Wyoming is similar to those of Calochortus gunnisonii S. Wats. (0.03–0.71 plants/m2) and A. textile (0.05–0.42 plants/m2) recorded by Wandsnider and Chung (2003) from their 1-m square sample plots in the grasslands of northwestern Nebraska. Densities similar to those found during these studies were also observed for Cymopterus montanus (Nutt.) T. & G. and A. textile in sagebrush and grass/forb vegetation communities surrounding the Bates Hole site in central Wyoming (Moe and Todd 1983:174). The density of C. bulbosus also compares with the less than 1.00 plant/m2 for Cymopterus acaulis (Pursh) Raf. and C. montanus noted in undisturbed steppe of northeastern Colorado, though the maximum density of C. montanus in disturbed contexts was 39.00 plants/m2 (Hazlett 1991).
These densities, however, are lower than those recorded for geophytes in sagebrush steppe and juniper woodlands of the northern Great Basin of central Oregon. Using 1-m2 sampling quadrats, Prouty (1995:144) calculated densities of 4.37 and 13.00 plants/m2 for Lomatium canbyi, 11.00 plants/m2 for Allium acuminatum Hook., and 2.04 and 3.30 plants/m2 for Calochortus macrocarpus Dougl. Densities of Lomatium canbyi and L. cous have also been found up to 33.00 and 32.00 plants/m2, respectively, in the northwestern United States (Hunn and French 1981). Balsamorhiza sagittata (Pursh) Nutt. grow at densities of 45.00 to 80.00 plants/m2 on the Canadian Plateau (Peacock 1998:185). The contrast is even greater for geophytes that grow in wet meadows. For example, Thoms (1989:169) recorded densities up to 300.00 plants/m2 for wet meadow species such as Camassia quamash (Pursh) Green.
Root Weight
The weights of C. bulbosus roots were quite variable, ranging from as little as 2 g to as much as 45 g (Figure 6). This size grading probably represents differences in the ages of the plants. The roots collected from Patch 1 (CS 1 and 2) on desert pavement adjacent to Site 48SW6965 had a considerably higher average weight (19.3 and 17.2 g) than those collected from Patch 4 (CS 4) on an alluvial silt terrace adjacent to Site 48SW6962 (9.0 g). The average weight of the roots collected also affected the resulting return rate; the collection of smaller roots yielded lower overall return rates.

Comparisons of C. bulbosus root weight for the three collection trials: collection sample 1 (Patch 1); collection sample 2 (Patch 1); and collection sample 4 (Patch 4).
Nutritional Composition
Nutritional composition analyses of C. bulbosus roots and A. textile bulbs indicate many similarities (Table 2). The number of kcal per 100 g of fresh weight (of which at least 70% is water content) and the percentage of carbohydrates and total sugar contents are comparable. Caloric content compares closely with the 92 kcal/100 g for Calochortus nuttallii T. & G. (sego lily) (Smith et al. 2001), 108 kcal/100 g for Lomatium canbyi, and 127 kcal/100 g for L. cous (Hunn and French 1981). Total sugar content compares closely with the 2.8% total sugar obtained for Cymopterus acaulis, but lower than the 4.5% for Cymopterus montanus (Hazlett 1991). These results are also similar to those obtained for the pioneering study of Yanovsky and Kingsbury (1938).
Nutritional composition of Cymopterus bulbosus and Allium textile (Medallion Labs, Minneapolis, Minnesota).

The major difference between C. bulbosus and A. textile is in their complex carbohydrate composition. C. bulbosus contains mostly starch, while A. textile has inulin. Taxa that contain the storage polysaccharides fructan and inulin require long-term heat treatment to effectively hydrolyse these complex carbohydrates to simpler sugars more suitable for human digestion (Wandsnider 1997). Ethnographically, this processing was usually accomplished in pit ovens. Because of the lack of inulin in C. bulbosus roots, long-term heat treatment of the roots is most likely not required for human digestibility, though heat treatment may increase the human digestibility of other starches such as those in Calochortus nuttallii (Smith et al. 2001). Therefore, clues in the archaeological record for the processing of C. bulbosus by prehistoric foragers would necessitate some sort of evidence other than the presence of pit ovens adjacent to geophyte patches, the proxy often considered in archaeological studies of the prehistoric use of inulin-rich species including Camassia quamash (camas) and Balsamorhiza sagittata (arrowleaf balsam-root) (Peacock 1998; Thoms 1989).
Total Calories Available from Patches
Total food energy provided in a patch, total collection time for the entire patch, and total time the patch would support a family of four were calculated for C. bulbosus in Patches 1–3 (Table 3). We estimated that the desert pavement Patch 1 would have potentially provided between 474,417 to 931,143 kcal, more than enough for a family of four for about 2–4 months, and the desert pavement Patch 3 would have produced 2,667,978 to 5,721,330 kcal, enough for a family of four for up to a year or two. In contrast, Patch 2 on the alluvial terrace would have potentially yielded only 43,881 to 94,102 kcal, about 1–2 weeks supply for a family of four. Additional uninvestigated C. bulbosus patches are present in the vicinity of each of these sites, so the total potential C. bulbosus resource base at each site is even greater than the above figures indicate.
Total calories (kcal) available for the three C. bulbosus patches (Patches 1–3).

These calculations suggest that the C. bulbosus patches (particularly those on desert pavement), would have provided more than enough calories to support forager groups through at least the season of availability, especially considering that other resources such as small and large animals would also have contributed to their diet. Though the calories available from some of the patches would have supported a forager family for a considerable length of time, the entire patch would not have been harvested during any single season. A point of diminishing returns would have been reached when most of the larger, more densely occurring plants had been collected. As shown by the collection trials of the smaller plants, the return rates after a certain point would have been considerably reduced to a level where further collection would not have been worth the effort.
Return Rates
An indication of the role that roots such as C. bulbosus may have played in prehistoric forager subsistence can be obtained within a behavioral ecological framework. The heuristic diet breadth model derived from optimal foraging theory offers one avenue for exploring the potential use of resources. It suggests hypotheses that predict an optimal pattern of behavior based on the costs and benefits of alternatives (Broughton and O'Connell 1999; Kelly 1995; Zeanah and Simms 1999). Comparisons between predicted and observed patterns provide an opportunity to gain a clearer understanding of forager behavior. This model has been applied with success to the archaeological record of the Great Basin (Kelly 2001; Zeanah 2004) and to the study of modern hunter-gatherers, including investigations of sexual division of labor (Bliege Bird 1999; Hawkes 1996).
The diet breadth model predicts whether a forager will take or ignore a resource when it is encountered while foraging (MacArthur and Pianka 1966; Stephens and Krebs 1986). Two of the most important predictions of the diet breadth model are: (1) the highest-ranked resource are always taken upon encounter; and (2) the inclusion of lower-ranked resources in the diet depends, not on their abundance, but rather on encounter rates with higher ranked resources. For the diet breadth model to be of use in understanding forager subsistence, the resources need to be ranked from the most to the least efficient. One currency used to rank resources is the return rate.
Simms (1984:76–87) calculated return rates by using the number of calories that an organism provides divided by the handling costs expressed as time. Included in handling costs are pursuit time (or the time required to pursue or collect a particular prey) and processing time (or the time necessary to process the resource into a usable form). Excluded from the return rates are the costs associated with searching for and locating resources. Though return rates expressed as kcal per hour may suggest a false precision, they are used only to obtain a relative ranking of resources so that predictions can be made. Search and handling costs are kept separate so that the effects of changes in one can be evaluated independently of the other (Kelly 1995:80). Lower ranked resources usually require less search time but have higher handling costs than higher ranked resources. However, technological improvements may be introduced to reduce handling costs for lower ranked resources (Broughton and O'Connell 1999; Ugan et al. 2003).
Using the kcal per gram shown in Table 2 (1.18 kcal/g for C. bulbosus and 0.94 kcal/g for A. textile), return rates were calculated for the three 30-minute C. bulbosus collection trials (CS 1, 2 and 4) and the one A. textile trial (CS 6) (Table 4). The return rates calculated from these experiments are 1,054–1867 kcal/hr for C. bulbosus and 230 kcal/hr for A. textile. C. bulbosus is at the higher end of the range for North American geophytes based on studies from Great Basin and Columbia Plateau and A. textile is at the lower end of the range (Figure 7). A similar range of variation in return rates has also been obtained for geophytes from throughout the world (O'Connell et al. 1983, 1999; Vincent 1985).
Return Rates for Cymopterus bulbosus and Allium textile calculated from 30-minute collection trials at Patches 1 and 4.


Return rates for various geophytes compared to relative resource ranking for animals and plants (after Simms 1984).
Because many geophytes provide approximately the same kcal per gram, the major reasons for such a variation in return rates among the different taxa appear to be their density within a patch and the size of the underground storage organ. Camassia quamash, for example, grows in extremely dense stands of up to 300 plants/m2 in wet meadows, allowing for easy and quick collection. As calculated by Thoms (1989:235–236) from ethnographic information, they may provide initial return rates of 5,479 kcal/hr (collection rate) and net return rates (which includes processing, transportation, and storage) of 2,042 kcal/hr. In contrast, Calochortus nuttallii has a return rate of only 207 kcal/hr because of its lower density patches and the small size of its bulb (Smith et al. 2001).
The high return rate for C. bulbosus (1,054 to 1,867 kcal/hr) compared to A. textile (230 kcal/hr) obtained during the current study is the result of the difference in the size of the root or bulb and the ease of collection. A. textile has a considerably smaller bulb, and its thin stem makes removing them from the soil matrix more difficult. The return rate for A. textile would be even lower if the additional processing, consisting of removal of a fibrous sheath and long-term heat treatment to hydrolyse the complex carbohydrate inulin, was figured into the calculation. It would probably be comparable to Calochortus nuttallii, another taxon requiring long-term heat treatment for best human digestibility (Smith et al. 2001).
Given these differing return rates and their similar periods of availability, prehistoric foragers would most likely have chosen C. bulbosus for procurement over A. textile, all else being equal. As recorded by ethnographic studies of the Northern Paiute in the northern Great Basin, root diggers selected particular species upon which to focus their effort and typically chose those that could be gathered most easily even though other species occurred at higher densities or nearby (Couture et al. 1986).
Conclusions
The determination of return rates, patch size, and density of C. bulbosus near the archaeological sites in the Green River Basin of Wyoming can help us assess whether these roots would have been a sufficiently viable resource to have influenced prehistoric campsite selection. These data are especially useful because evidence for the use of C. bulbosus is typically nonexistent in the archaeological record. This study demonstrates that large desert pavement patches containing even comparatively low densities of C. bulbosus had the potential to furnish a significant number of calories for the prehistoric foragers of the area. Therefore, patches of geophytes may have exerted considerable influence on settlement and subsistence decisions and may have been at least one reason—if not the primary reason—that the particular archaeological sites adjacent to these patches were repeatedly occupied. These locations would likely have been occupied during at least April and May when the plants were visible above ground, and the ground was moist enough to facilitate digging. The early spring was especially important because prehistoric foragers would have been critically in need of carbohydrates at the end of the lean winter season. The roots appear to have been collected for immediate consumption, as no evidence is present for intensive exploitation of the resource or long-term storage.
Our investigation also suggests that not all geophyte species growing in the area would have had equal importance in determining camp locations. The selection of one species over another probably was influenced by the general size of the root or bulb, the ease of collection, and plant density. Comparisons between the return rates of C. bulbosus and A. textile suggest that though A. textile is more abundant, prehistoric foragers would have first chosen and exploited C. bulbosus. C. bulbosus has a larger underground storage organ and is easier to dig up than A. textile. The net return rate for A. textile would have been even lower if the bulbs were processed using long-term heat treatment to hydrolyse the complex carbohydrate inulin to more simple sugars for best human digestion (Wandsnider 1997) and if their fibrous coverings were removed.
The variability in the size of the roots between patches and among the individual plants within a patch probably greatly influenced prehistoric foragers’ selection of patches to exploit from year to year and hence camp location. Ethnographic studies in the northwestern United States and western Canada have shown that forager groups generally chose the patches with the largest plants and moistest soil conditions for exploitation during any one year (Couture et al. 1986; Peacock 1998). Within a particular patch, only plants within a certain size range were collected; the smaller ones were left to grow big enough for future harvest. The patches were probably harvested on a rotating basis, allowing them to regenerate over a period of years, thus resulting in an episodic use of particular camp locations associated with the patches (Smith and McNees 1999). The actual intervals between use is unknown but was probably longer than the 4 or 5 years observed as average maturation rates for other geophyte species in highly productive areas of the northwestern United States (Thoms 1989:173). This alternating use of the patches would have necessitated a cyclic use of the landscape, with the repetitive occupation of camp locations over a period of decades and centuries.
Footnotes
Acknowledgments
This study was conducted as part of the archaeological data recovery excavations completed by TRC Mariah Associates Inc. for the Williams Pipeline Rockies Expansion Project, Muddy Creek Loop. We thank Williams for their interest and funding and especially James Bloemker, Williams archaeologist, for his help and coordination during the project. Ann Kelsey of the University of Utah Garrett Herbarium identified the plant specimens. We thank LuAnn Wandsnider, Donn Todt, Nan Hannon, and an anonymous reviewer for furnishing useful comments that improved the paper.