
Abstract
Enterococcus faecalis, a common constituent of human and animal gut microbiota, has emerged as a significant pathogen due to its antimicrobial resistance (AMR) capabilities. This study investigates AMR profiles, virulence factors, and genetic diversity of E. faecalis isolates from broiler farms in Ludhiana, Punjab, India, to explore transmission dynamics between poultry and handlers. A total of 240 samples were collected from 20 farms, comprising 200 broiler droppings, 34 hand swabs from poultry handlers, and 6 human stool samples. Selective media and standard microbiological techniques were employed to isolate and identify E. faecalis. Whole genome sequencing (WGS) was performed on representative isolates to uncover antimicrobial resistance genes (ARGs) and virulence factors. E. faecalis was isolated from 47% of broiler droppings and 26.47% of handler hand swabs, whereas no isolates were recovered from stool samples. High resistance was observed for erythromycin (96.11%), tetracycline (78.64%), ciprofloxacin (76.69%), streptomycin (76.69%), and linezolid (65.04%), with 83.49% of isolates exhibiting multidrug resistance (MDR). Vancomycin resistance genes (vanC1–34.95% and vanC2/C3–9.70%) were also detected. WGS analysis of four isolates identified ARGs such as dfrE, efrA, vanT gene in vanG cluster, vanY gene in vanB cluster, ermB, tet45, tetM, tetA, aac(6′)-Ie-aph(2″)-Ia, ant(4′)-Ib, aph(3′)-Ia, sul1, sul3, mphA, qnrS1, and catA8, as well as virulence factors associated with biofilm formation, adherence, capsule formation, and protease production. Sequence typing identified was ST-1866, ST-7317, and ST-403, with ST-7317 common between broiler droppings and handler swab isolates, indicating potential transmission. While these findings highlight poultry environments as reservoirs for MDR E. faecalis, the directionality of transmission (zoonotic vs. reverse zoonotic) remains unclear. This underscores the need for expanded surveillance and molecular studies to better understand transmission dynamics and mitigate risks to farm workers and public health.
Keywords
Introduction
The poultry industry is a vital source of income and nutrition globally (Kalam et al, 2022; Otiang et al, 2020; Samboko et al, 2018). However, rising demand, driven by population growth, has led to widespread unregulated antimicrobial use, especially in low- and middle-income countries (LMICs), contributing to the global rise of antimicrobial resistance (AMR) (de Mesquita Souza Saraiva et al, 2022; Masud et al, 2020; Van Boeckel et al, 2015). India, like many LMICs, lacks a comprehensive regulatory framework governing antimicrobial use in livestock, including poultry, exacerbating the risk of AMR (Van Boeckel et al, 2015).
Among resistant bacteria, Enterococcus species are particularly concerning. These commensal bacteria inhabit the gastrointestinal tracts of humans and animals, including poultry (Comerlato et al, 2020; Escobedo-Hinojosa and Pardo-López, 2017) but have emerged as nosocomial opportunistic pathogens, exhibiting increasing resistance to antimicrobial agents, notably vancomycin-resistant enterococci (VRE) (Cetinkaya et al, 2000; Jones et al, 2004). VRE have been detected in the feces of healthy farm animals, animal-derived food products, and sewage (Bates, 1997). In numerous countries, unregulated antibiotic use in livestock and poultry is believed to contribute to VRE colonization in these animals and pets, potentially facilitating human colonization through direct contact or the food chain (Bates, 1997). Beyond food-producing animals, antimicrobial-resistant enterococci have been identified in retail food items, including meat, vegetables, cheese, and milk (Al-Shammary, 2019; Fard et al, 2019; Jahansepas et al, 2020; Sabença et al, 2020), raising concerns regarding the dissemination of resistant organisms (Alonso et al, 2019). Transmission of enterococci from these sources may augment the human reservoir of antibiotic-resistant enterococci, particularly VRE (Hammerum et al, 2010).
Currently, the majority of nosocomial enterococcal infections are attributed to either E. faecalis or E. faecium, with E. faecalis being more prevalent (Hammerum et al, 2010). In addition to hospital settings, VRE colonization has been reported in community settings, suggesting the existence of community-based reservoir (Bates et al, 1994; Goel et al, 2016; Wegener, 2003). Antibiotic-resistant enterococci from animal sources, such as poultry and animal-derived foods, could play a crucial role in human colonization and infection outside health care settings (Bates et al, 1994; Mathur and Singh, 2013). These resistant organisms may subsequently disseminate from the community to hospital environments. Managing enterococcal infections is challenging due to resistance to antibiotics such as beta-lactams, cephalosporins, macrolides (erythromycin), glycopeptides (vancomycin), high-level aminoglycosides (e.g., streptomycin), and oxazolidinones (linezolid) (Arias and Murray, 2012; Kaye and Kaye, 2000; Torres et al, 2018). This has necessitated inclusion of Enterococcus species in global surveillance (Rogers et al, 2021). While European and North American regions have established AMR monitoring systems in animals (European Food Safety Authority, European Centre for Disease Prevention and Control, 2018), LMICs often lack effective surveillance in the animal sector, impeding global harmonization of AMR surveillance (Van Boeckel et al, 2019). This is particularly concerning given the potential role of animals, such as poultry, in facilitating the spread of resistant bacteria.
Whole genome sequencing (WGS) is transforming AMR surveillance by providing detailed insights into antimicrobial resistance genes (ARGs), virulence factors, and phylogenetic relationships (Hammerum, 2012; Pinholt et al, 2015). Despite extensive studies on E. faecalis in human infections, zoonotic transmission between poultry and humans remains underexplored. Understanding AMR profiles, virulence factors, and clonal relationships of E. faecalis from poultry and their handlers is essential for assessing risks of transmission for potential community colonization. This study investigated AMR profiles of E. faecalis in broiler farms in Punjab, India, using WGS, to explore potential transmission risks between broilers and handlers.
Materials and Methods
Sampling design
This study surveyed 20 broiler farms in Ludhiana, Punjab, India, from November 2021 and March 2023, collecting 240 samples: 200 broiler droppings, 34 handler hand swabs, and 6 handler stool samples (Supplementary Table S1). Droppings were randomly collected in sterile containers, and hand swabs were collected using sterile cotton swabs moistened with sterile buffered peptone water. Hand swabs and stool samples were collected from consenting poultry handlers who were routinely involved in poultry farm management. All samples were transported on ice and processed immediately. Metadata from farm owners at the sampled poultry farms was gathered through a structured questionnaire (Supplementary Table S2). Before administering the questionnaire, the study’s objectives were explained to all participants. All were informed about the voluntary nature of participation, data confidentiality, and use of data for research purposes. Explicit consent was obtained for the collection of hand swabs and stool samples from handlers, with participants briefed on the procedures, their voluntary participation, and data confidentiality.
Sample processing, bacterial isolation, and identification
In this study, fecal samples were processed with slight modifications to the method by Isichei-Ukah et al (2024), whereas hand swabs followed the method outlined by Willey et al (1999). Broiler droppings and stool samples (1 g each) were inoculated into 9 mL of tryptone soy broth with 6.5% NaCl and incubated at 37°C for 24 h. Hand swabs were enriched in brain heart infusion broth under same conditions. After enrichment, cultures were streaked onto bile esculin agar (BEA), where black colonies were presumptively identified as Enterococcus species (Isichei-Ukah et al, 2024). Species confirmation of E. faecalis was performed using matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS), with scores >2.3 indicating highly probable species identification, 2.0–2.3 secure genus and probable species, 1.7–2.0 probable genus, and <1.7 unreliable identification (Stępień-Pyśniak et al, 2018; Tracz et al, 2016). To further validate E. faecalis identification, species-specific polymerase chain reaction (PCR) was performed on MALDI-TOF MS-confirmed isolates, following the method described by Dutka-Malen et al (1995).
Phenotypic and genotypic AMR characterization
E. faecalis isolates were tested for AMR using the Kirby–Bauer disk diffusion method for 10 antibiotics and broth microdilution for vancomycin, following Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2020). Antibiotics tested included erythromycin (15 µg), ciprofloxacin (5 µg), levofloxacin (5 µg), vancomycin (960 µg/mg), gentamicin (10 µg), streptomycin (10 µg), ampicillin (10 µg), penicillin G (10 µg), fosfomycin (200 µg), linezolid (30 µg), and tetracycline (30 µg). Vancomycin resistance was assessed by broth microdilution, with a minimum inhibitory concentration (MIC) of ≤4 µg/mL indicating sensitivity, 8–16 µg/mL intermediate, and ≥32 µg/mL resistant (CLSI, 2020). Isolates resistant to at least one antibiotic in three or more classes were classified as multidrug resistance (MDR; Magiorakos et al, 2012). DNA extraction from isolates utilized the snap-chill protocol and vancomycin resistance genes (vanA, vanB, vanC1, and vanC2/C3) were screened as per the protocol by Kariyama et al (2000).
Data analysis
Statistical analysis was performed using Epitools to calculate odds ratios with 95% confidence intervals and corresponding p-values (Table 1).
Influence of Farm Variables on the Occurrence of Enterococcus faecalis

CI, confidence interval; R, reference; p < 0.05.
WGS of E. faecalis isolates
Four E. faecalis isolates, selected based on vancomycin resistance and MDR profiles, underwent WGS. Genomic DNA was extracted using the HiPurA Bacterial Genomic DNA Isolation Kit (HiMedia, India), with quality assessed through agarose gel electrophoresis, NanoDrop spectrophotometry (ThermoFisher, USA), and Qubit assay (ThermoFisher, USA). Samples with intact bands, 50 ng/μL concentration, and A260/280 ≥1.80 were selected for library preparation. Libraries were prepared using the KAPA HyperPlus kit and sequenced on an Illumina NovaSeq 6000 platform (2 × 150 bp reads). Reads were quality-filtered with Trimmomatic (v0.36) (Bolger et al, 2014) and aligned to the E. faecalis EnGen0336 reference genome using Burrows–Wheeler Aligner (v0.7.12). Consensus sequences were generated using samtools (v1.17) (Li et al, 2009), annotated using Prokka (v1.14.6) (Seemann, 2014), and coverage/depth assessed via BEDtools (v2.30.0) (Quinlan and Hall, 2010). WGS metrics are detailed in Supplementary Table S3.
In silico analysis
Assembled sequences were screened for ARGs using the Resistance Gene Identifier (RGI 6.0.1, https://card.mcmaster.ca/analyze/rgi). Plasmid replicon types were determined using the PlasmidFinder 2.1 (https://cge.cbs.dtu.dk/services/PlasmidFinder/). Mobile genetic elements (MGEs) and integrons were detected using MobileElementFinder (https://cge.food.dtu.dk/services/MobileElementFinder/) and IntegronFinder (Galaxy@Pasteur). Virulence factors were analyzed using VFanalyzer (http://www.mgc.ac.cn/cgi-bin/VFs/v5/main.cgi). Multilocus sequence typing (MLST) was performed using the PubMLST platform (https://pubmlst.org/). Phylogenetic relationships were determined through core gene analysis. Core genes were extracted using GET_HOMOLOGUES (Contreras-Moreira and Vinuesa, 2013) and aligned with MAFFT v7 (https://mafft.cbrc.jp/alignment/server/phylogeny.html) (Katoh et al, 2002). A neighbor-joining tree was constructed and visualized using FigTree v1.4.3 (http://tree.bio.ed.ac.uk/software/Fig.tree/). Average nucleotide identity (ANI) was calculated using pyani v0.2.10 and pairwise alignment scores from NUCmer tool from MUMmer v3.23 (Pritchard et al, 2016). Whole genome alignments were performed with the Progressive Mauve Program in Mauve v20150226.
Accession numbers of isolates
WGS data were submitted to National Center for Biotechnology Information under BioProject PRJNA943603 and GenBank accession numbers: CP120226, CP120227, CP120228, and CP120229.
Results
Isolation and identification of E. faecalis
Out of 240 enriched samples streaked on BEA, 132 samples (55%) showed black colonies of Enterococcus species. From these, 396 colonies (3 colonies/positive sample) were randomly selected, and 225 were confirmed as E. faecalis using MALDI-TOF MS. One confirmed isolate per positive sample was further analyzed, resulting in 103 E. faecalis isolates from 103 positive samples. Additionally, all MALDI-TOF MS-confirmed isolates were validated as E. faecalis through PCR assay. The occurrence of Enterococcus species was 59.5% (119/200) in broiler droppings, 38.23% (13/34) in handler hand swabs, and 0% in human stool samples. E. faecalis isolation rates were 42.9% (103/240), with 47% in broiler droppings and 26.47% in hand swabs. Farms employed a deep litter system, with all respondents being male (100%). Most farmers (90%) were graduates, and 85% stored antibiotics, including quinolones (75%), aminoglycosides (60%), tetracyclines (60%), and second-generation cephalosporins (10%). Veterinarians treated 75% of farms, whereas 25% sought nonveterinary care. No significant association was found between E. faecalis occurrence and farmer education, veterinarian treatment, or antibiotic storage (p > 0.05) (Table 1).
AMR profiling
High resistance was observed to erythromycin (96.11%), tetracycline (78.64%), ciprofloxacin (76.69%), streptomycin (76.69%), and levofloxacin (72.81%). Resistance was lower for penicillin G (28.15%), gentamicin (15.53%), ampicillin (3.88%), and fosfomycin (1.94%; Supplementary Table S4). Vancomycin resistance was detected in 23.30% of isolates, with 83.49% identified as MDR. The vanC1 and vanC2/C3 genes were present in 34.95% and 9.70% of isolates, respectively, whereas vanA and vanB were absent.
Chromosomal and plasmid ARGs
WGS revealed chromosomal ARGs conferring resistance to macrolides (efrA), fluoroquinolones (efrA), rifamycins (efrA), glycopeptides (vanT), and diaminopyrimidines (dfrE; Table 2). Additionally, several plasmid-borne ARGs were identified in the isolates (Table 2).
Antimicrobial Susceptibility Results of Selected Enterococcus faecalis Isolates

Antibiotics: E, erythromycin; CIP, ciprofloxacin; LE, levofloxacin; GEN, gentamicin; S, streptomycin; P, penicillin G; LZ, linezolid; TE, tetracycline; MDR, multidrug resistance.
ARG, antimicrobial resistance genes; NCBI, National Center for Biotechnology Information.
Virulence genes
Isolates contained virulence genes for adherence (ace, ebpA, ebpB, ebpC, srtC, and efaA), capsule formation (cpsA/uppS and cpsB/cdsA), biofilm formation (bopD, fsrA, fsrB, and fsrC), gelatinase production (gelE), and SprE enzyme production (sprE; Table 3).
Summary of MLST Profiles, Virulence Factors, Plasmid Types, and Plasmid Insertion Sequences in Enterococcus faecalis Isolates

MLST, multilocus sequence typing; NA, not available.
MGEs and MLST
Plasmid replicon types repUS43, rep9a, rep7a, and IncFII(K) were found in E. faecalis isolates with identical plasmid types (repUS43 and rep9a) in broiler (11P7) and handler isolates (11PHS) from the same farm (Table 3). No genomic integrons or transposons were found, with only one isolate (14P4) contained plasmid insertion sequences (Table 3). MLST identified ST-1866, ST-7317, and ST-403, with ST-7317 shared between broiler (11P7) and handler samples (11PHS).
Phylogenetic analysis
Core genome analysis utilizing 2028 core genes revealed that all study isolates clustered into a single clade. They shared a lineage with cattle fecal isolates from Switzerland and India, cattle handler hand isolates from India, and poultry fecal isolate from Canada (Fig. 1).
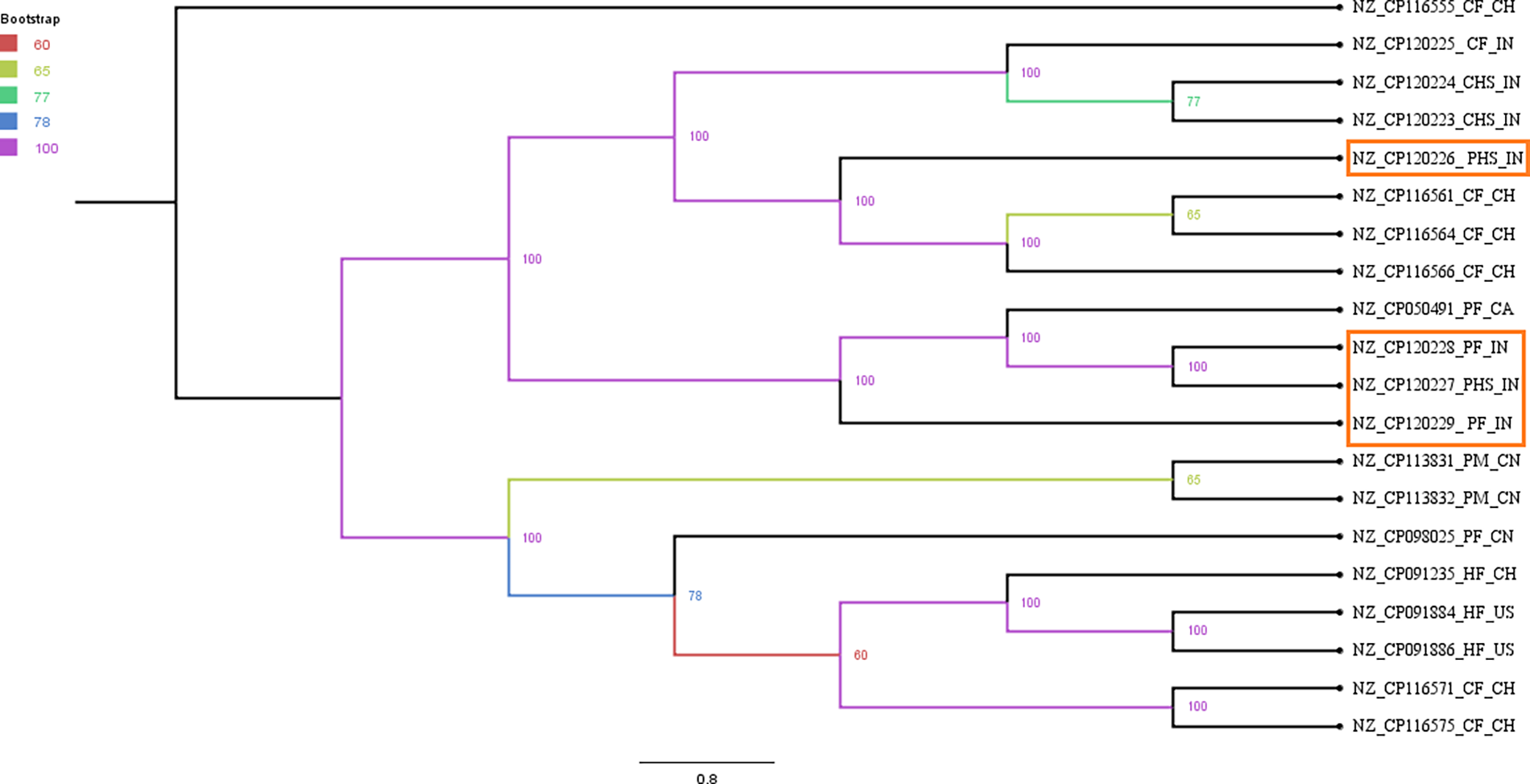
Phylogenetic relationship among Enterococcus faecalis isolates based on their core genomes. Branch coloring has been done as per bootstrap values with 100 replicates. Bootstrap values of nodes have also been shown. The E. faecalis isolates from current study have been highlighted with red color outline. Letters for the isolates after accession numbers indicate the host/isolation source of different E. faecalis isolates: CF, cattle feces; CHS, cattle handler hand isolate; PF, poultry feces; PHS, poultry handler hand swabs; PM, poultry manure; HF, human feces. The last two letters indicate the geographical locations: CH, Switzerland; IN, India; CA, Canada; CN, China; US, USA.
ANI and comparative genomics
ANI revealed that all study isolates shared >98% nucleotide identity with alignment coverage ranging from 81.6% to 100%. These isolates closely resembled E. faecalis strains from cattle and poultry feces, human feces, and cattle handler hand swabs from Switzerland, China, the USA, Canada, and India (Fig. 2).Whole genome alignment of study isolates with the reference genome E. faecalis EnGen0336 revealed local collinear blocks in blue and yellow, indicating conserved DNA regions (Fig. 3). Green lines indicated rearrangements such as inversions or translocations. Well-aligned blocks represented conserved sequences, whereas gaps or misaligned blocks marked structural variations. Red bars highlighted sequence differences (insertions and deletions).
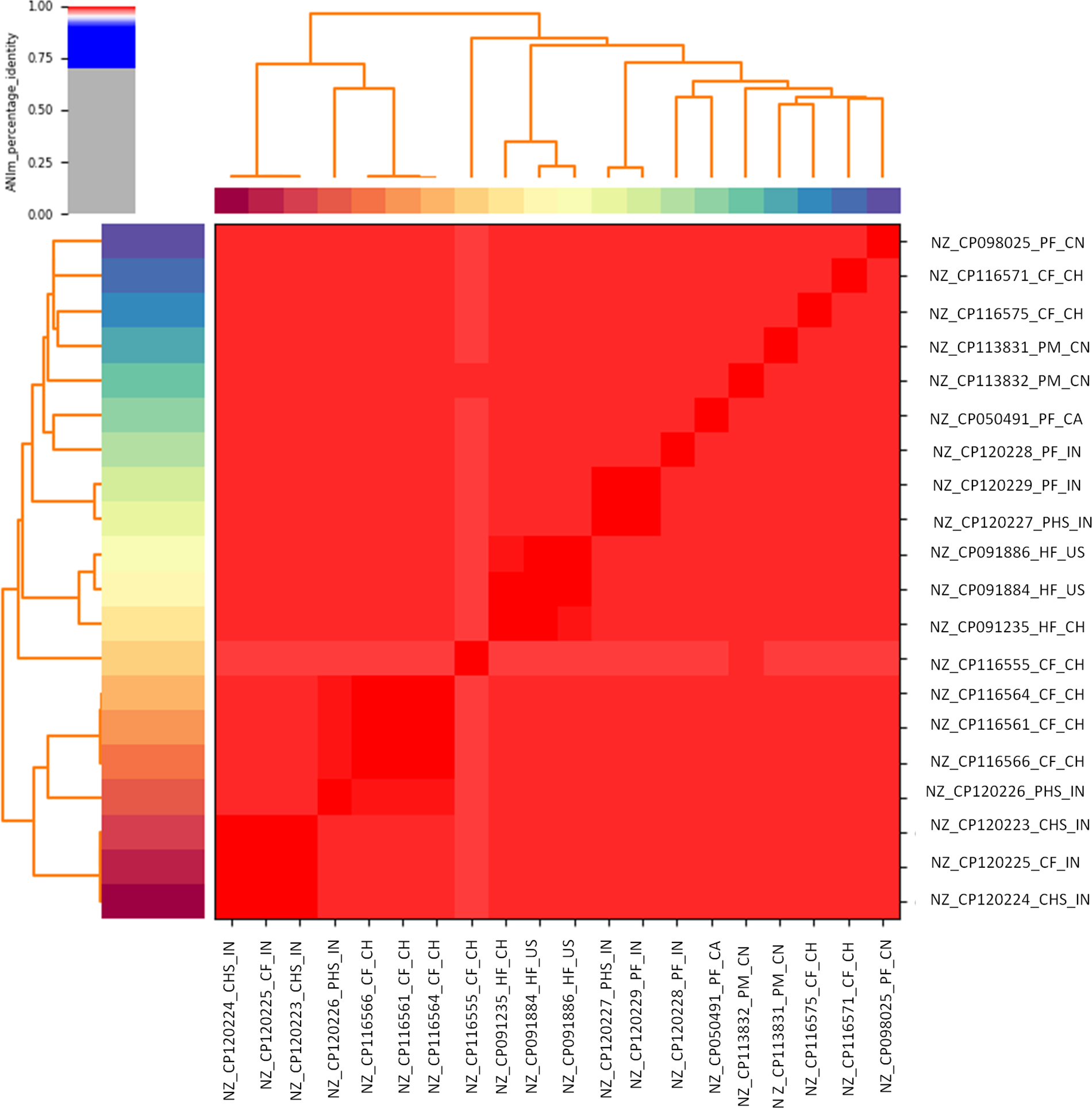
Average nucleotide sequence identity of complete genome of Enterococcus faecalis isolates of the present study (NZ_CP120226_PHS_IN, NZ_CP120227_PHS_IN, NZ_CP120228_PF_IN, and NZ_CP120229_PF_IN) with complete genomes of E. faecalis taken from NCBI database.

Synteny map generated by the Mauve genome alignment tool showing the alignment of genomes of Enterococcus faecalis strains (NZ_CP120226, NZ_CP120227, NZ_CP120228, and NZ_CP120229), represented by the FASTA files listed on the left, aligned against a reference genome (E. faecalis EnGen0336.fasta).
Discussion
The isolation rate of E. faecalis from broiler farms in this study was 42.91% (103/240), aligning with reported rates in Rajasthan, India, ranging from 34% (Sharma et al, 2022) to 50% (Kanchan Jangir et al, 2015). Globally, prevalence varies, with isolation rates of 0%, 11.5%, and 33.62% reported in poultry cloacal swabs from Egypt (Osman et al, 2019), Ethiopia (Bekele and Ashenafi, 2010), and Turkey (Ünal et al, 2017), respectively, and 37.9% in poultry droppings from Zambia (Mwikuma et al, 2023). These studies also identified other Enterococcus species, including E. faecium and E. gallinarum. However, the focus of the current study was on E. faecalis due to its higher pathogenic potential (Georges et al, 2022; Archambaud et al, 2024). Differences in isolation rates can be attributed to factors, including geographical variations, laboratory methodologies, sample types and numbers, and farm biosecurity practices (Hayes et al, 2003).
The World Health Organization (WHO) classifies antimicrobials based on their importance in human medicine, categorizing them as critically important (highest and high priority), highly important, and important (World Health Organization, 2019). This classification aids in responsible antibiotic use and prioritizes antibiotics for inclusion in AMR surveillance programs. In this study, E. faecalis isolates showed significant resistance to WHO-classified antibiotics. Among highest priority, erythromycin had the highest resistance (96.11%), followed by ciprofloxacin (76.69%), levofloxacin (72.81%), and vancomycin (23.30%). Resistance to high priority antibiotics included streptomycin (76.69%), linezolid (65.04%), penicillin (28.15%), gentamicin (15.53%), ampicillin (3.88%), and fosfomycin (1.94%). Resistance to tetracycline (highly important) was 78.64%. These resistance patterns are concerning as they narrow treatment options and increase the risk of horizontal gene transfer (HGT), exacerbating AMR in health care and communities.
The highest resistance to erythromycin and tetracycline in current study is similar to other studies (Hasan et al, 2018; Mwikuma et al, 2023). This high resistance to tetracyclines, fluoroquinolones, and macrolides is likely due to the overuse and inappropriate use of these antibiotics in poultry farming (Brower et al, 2017; Perumal et al, 2020). Tetracyclines and macrolides are commonly used in India due to their affordability and broad-spectrum efficacy, which has contributed to widespread resistance, with resistant isolates often found in poultry litter and farm soil (Perumal et al, 2020).
This study observed 23.30% vancomycin resistance in E. faecalis isolates, determined using MIC values. This resistance rate was higher than those reported in MIC-based studies, such as Rivera-Gomis et al (2021), who found 10.77% resistance in laying hen isolates, and Makarov et al (2022), who detected no resistance in chicken biomaterials. Hasan et al (2018) also reported complete vancomycin susceptibility in poultry isolates from Pakistan. Conversely, studies using disk diffusion method reported higher resistance rates in E. faecalis isolates from poultry droppings, including 32.8% resistance in Zambia (Mwikuma et al, 2023) and 35.3% intermediate resistance in Rajasthan, India (Sharma et al, 2022). Discrepancies between MIC and disk diffusion methods highlight differences in sensitivity and accuracy. While disk diffusion is practical and cost-effective, it may overestimate resistance or fail to distinguish strains with reduced susceptibility (Hageman et al, 2003). MIC provides more precise and reproducible results, especially for borderline resistance. A study in the United States has documented disk diffusion to be unreliable for vancomycin susceptibility testing in Enterococci (Hageman et al, 2003).
In this study, none of the isolates carried the vanA or vanB genes. However, the presence of vanC1 (specific to E. gallinarum) and vanC2/C3 genes (specific to E. casseliflavus) in E. faecalis strains suggests HGT as a likely mechanism. E. faecalis may acquire vanC genes from E. gallinarum and E. casseliflavus through HGT (Moura et al, 2013), as these genes are chromosomally located in their native hosts and found within transferable genetic elements such as transposons and integrons, which facilitate interspecies transfer (Depardieu et al, 2004; Fisher and Phillips, 2009; Navarro and Courvalin, 1994; Patel et al, 1997, 1998). Previous studies have reported vanC1 and vanC2/C3 in E. faecalis isolates from various sources. The vanC1 gene, first reported in E. faecalis strains from pig manure in Germany, was acquired from E. gallinarum (Schwaiger et al, 2012). Sagor et al (2022) identified vanC2 in 62.5% of vancomycin-resistant E. faecalis isolates from poultry cloacal swabs. El-Oraby et al. (2023) found vanC1 in 14.7% and vanC2/C3 in 5.9% of 34 vancomycin-resistant E. faecalis strains from poultry meat. The vanC gene has also been found in E. faecalis from pigeons, ducks, and broiler cloacal swabs (Moura et al, 2013; Osman et al, 2019).
WGS of selected E. faecalis isolates revealed resistance to macrolides, glycopeptides, quinolones, oxazolidinones, tetracyclines, aminoglycosides, and penicillins, supported by the identification of ARGs. However, the genetic basis for penicillin and linezolid resistance was not determined. Penicillin resistance may result from overexpressed penicillin-binding proteins (Duez et al, 2004; Murray, 1998), whereas linezolid resistance is linked to mutations in the 23S rRNA gene (Yadav et al, 2017). These findings align with previous WGS studies on Enterococcus species (Sharma et al, 2020; Tyson et al, 2018). The study also identified multiple plasmid replicon types, with rep9 being most prevalent. Plasmids promote HGT, facilitating the spread of resistance traits (Arredondo-Alonso et al, 2020). One isolate contained several insertion sequences, potentially enhancing ARG dissemination.
Our study identified E. faecalis virulence factors, including adherence genes (ace, ebpA-C, srtC, and efaA) for host attachment (Nallapareddy et al, 2008), cpsA and cpsB for antiphagocytosis (Opoku-Temeng et al, 2019), as well as biofilm (bopD and fsrA-C) and enzyme production (gelE, sprE) genes, which enhance bacterial pathogenicity (Oli et al, 2022; Strateva et al, 2016).
E. faecalis isolates were classified into ST-403, ST-1866, and ST-7317, with ST-403 previously linked to chicken meat in Korea (Choi and Woo, 2013). The detection of ST-7317 in both broilers and handlers suggests a potential transmission link. However, the study does not determine the directionality of transmission—whether zoonotic or reverse zoonotic. Both pathways are plausible, as E. faecalis is part of the gut microbiota in both humans and animals and can adapt to diverse ecological niches. Financial limitations constrained our WGS analysis to a few isolates and short-read sequencing technology. Despite these limitations, the data provided critical insights into the AMR profiles and virulence factors of E. faecalis isolates.
Conclusion
In this study, the high isolation rate, significant prevalence of MDR strains, and identification of vancomycin-resistant strains underscore substantial public health risks associated with poultry production. Findings on resistance genes, virulence factors, and potential intrafarm transmission emphasize the necessity for targeted interventions in poultry health management.
Authors’ Contributions
N.P.: Investigation, formal analysis, and writing—original draft. R.S.: Conceptualization, supervision, and writing—review and editing. A.T.: Formal analysis. S.K.: Resources. H.M.: Formal analysis. R.S.A.: Writing—review and editing. A.K.A.: Resources. J.P.S.G.: Conceptualization and writing—review and editing.
Footnotes
Author Disclosure Statement
The authors declare no conflict of interest. The authors have no relevant financial or nonfinancial interests to disclose.
Funding Information
This work was supported by the Indian Council of Agricultural research (ICAR, F.No.Edn.10(8)/2016-EP&HS), Rashtriya Krishi Vikas Yojana (RKVY, 12.4 2), and acknowledges Indian Council of Medical Research [ICMR, Reference No.: 3/1/3/JRF-2021/HRD-004 (1207299] for providing Junior Research Fellowship (JRF).
Ethics Statement
Ethical approval was not required for sample collection from broilers, as only freshly voided broiler droppings were collected, and no invasive techniques were applied. The entire survey and human sample collection activities were conducted in accordance with ethical standards. Approval for the study, including the survey questions and human sample collection, was obtained from the Institutional Review Board of Dayanand Medical College, Ludhiana, India (Reference no: DMCH/R&D/2021/7, Dated: 20th January, 2021). This approval ensured adherence to ethical guidelines for research involving human participants and survey administration.
Submission Declaration and Verification
The present work has not been published anywhere, and all authors are aware of the submission of present work.
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.