
Abstract
It is often assumed that the promise of a monetary bonus improves cognitive control. We show that in fact appetitive motivation can also impair cognitive control, depending on baseline levels of dopamine-synthesis capacity in the striatum. These data not only demonstrate that appetitive motivation can have paradoxical detrimental effects for cognitive control but also provide a mechanistic account of these effects.
Cognitive performance is generally thought to benefit from a promised bonus (Pessoa & Engelmann, 2010). In folk terms, being motivated implies being goal driven; that is, motivation is presumed to have purely beneficial consequences for people’s ability to direct their behavior to accomplish cognitive goals. In line with this intuition, studies have shown that appetitive motivation improves a wide range of cognitive-control functions (Jimura, Locke, & Braver, 2010; Krawczyk, Gazzaley, & D’Esposito, 2007; Pessoa & Engelmann, 2010), suggesting that appetitive motivation has general enhancing effects on cognition. Nevertheless, inconsistent results have been obtained in studies investigating the effects of reward on cognitive-control functions such as focused attention (Braem, Verguts, Roggeman, & Notebaert, 2012; Padmala & Pessoa, 2011; Sasaki, Nanez, & Watanabe, 2010; van Steenbergen, Band, & Hommel, 2009). It has been speculated that individual differences in levels of the neurotransmitter dopamine play an important role in the effects of motivation on cognitive control (Mobbs et al., 2009; Pessoa & Engelmann, 2010; van Steenbergen et al., 2009). Dopamine has long been known to play an important role in reward and motivation (Berridge & Robinson, 1998; Robbins & Everitt, 1992). However, none of the studies just cited have actually accounted for dopamine processing, which might explain the inconsistencies observed.
In the current study, participants underwent positron-emission tomography (PET) so that we could assess 6-[18F]fluoro-
Method
Participants
We invited 33 participants in a previous PET-FMT study to participate in this behavioral study. Sixteen participants accepted, but 2 participants were excluded because of an inability to perform the task (error rates: 34% and 40%; overall mean = 18%, SD = 9%). Thus, our final sample consisted of 14 neurologically and psychologically healthy right-handed people (6 men, 8 women; mean age = 28 years, age range = 24–34 years, SD = 2.7). The time between the PET scans and behavioral measurements was an average of 2.3 years (SD = 1.1, range = 1.0–4.2 years), similar to the time gap in previous studies that have measured FMT uptake (Cools et al., 2009). All participants gave written informed consent and were paid for participation according to institutional guidelines of the local ethics committee (University of California, Berkeley, Committee for the Protection of Human Subjects).
PET data acquisition
PET imaging and FMT synthesis were performed at Lawrence Berkeley National Laboratory (Berkeley, CA). For more information about FMT synthesis, see VanBrocklin et al. (2004). FMT is comparable with [18F]fluorodopa, except that it is not a substrate for O-methylation in the periphery and therefore provides images with higher signal-to-noise ratios (Jordan et al., 1997). FMT is a substrate of the dopamine-synthesizing enzyme aromatic
PET scans were performed using an ECAT-HR PET camera (Siemens, Knoxville, TN), as described elsewhere (Landau, Lal, O’Neil, Baker, & Jagust, 2009). Approximately 2.5 mCi of high-activity FMT was injected as a bolus into an antecubital vein, and a dynamic acquisition sequence in three-dimensional mode was obtained for a total of 89 min.
Structural MRI
Two high-resolution anatomical images were acquired for each participant on a Siemens (Erlangen, Germany) 1.5-T Magnetom Avanto MRI system (magnetization-prepared rapid acquisition with gradient echo) with the use of a 12-channel head coil (echo time = 3.58 ms; repetition time = 2,120 ms; voxel size = 1.0 × 1.0 × 1.0 mm, 160 axial slices; field of view = 256 mm; scanning time ≈ 9 min). The two images were averaged to obtain one high-resolution structural image, which was used to generate individual striatal regions of interest (ROIs).
ROIs
Striatal subregions and the cerebellum were drawn manually on each participant’s anatomical MRI scan using FSLView (Smith et al., 2004; http://fsl.fmrib.ox.ac.uk/fsl/fslview/). The striatal ROIs were drawn according to guidelines described previously (Mawlawi et al., 2001). Both inter- and intrarater reliability were greater than 95% (ratings made by two laboratory members). We used the cerebellum as the reference region for calculating FMT signal intensity. To avoid contamination of FMT signal from the dopaminergic nuclei, we included only the posterior three fourths of the gray matter in the cerebellar reference region. After coregistration to PET-FMT space, to ensure a high probability that each voxel contained gray matter, we included only those voxels with a greater-than-50% chance of lying in a certain ROI. This procedure resulted in six participant-specific striatal ROIs: left and right ventral striatum, left and right dorsal caudate nucleus, and left and right dorsal putamen (Fig. 1a).
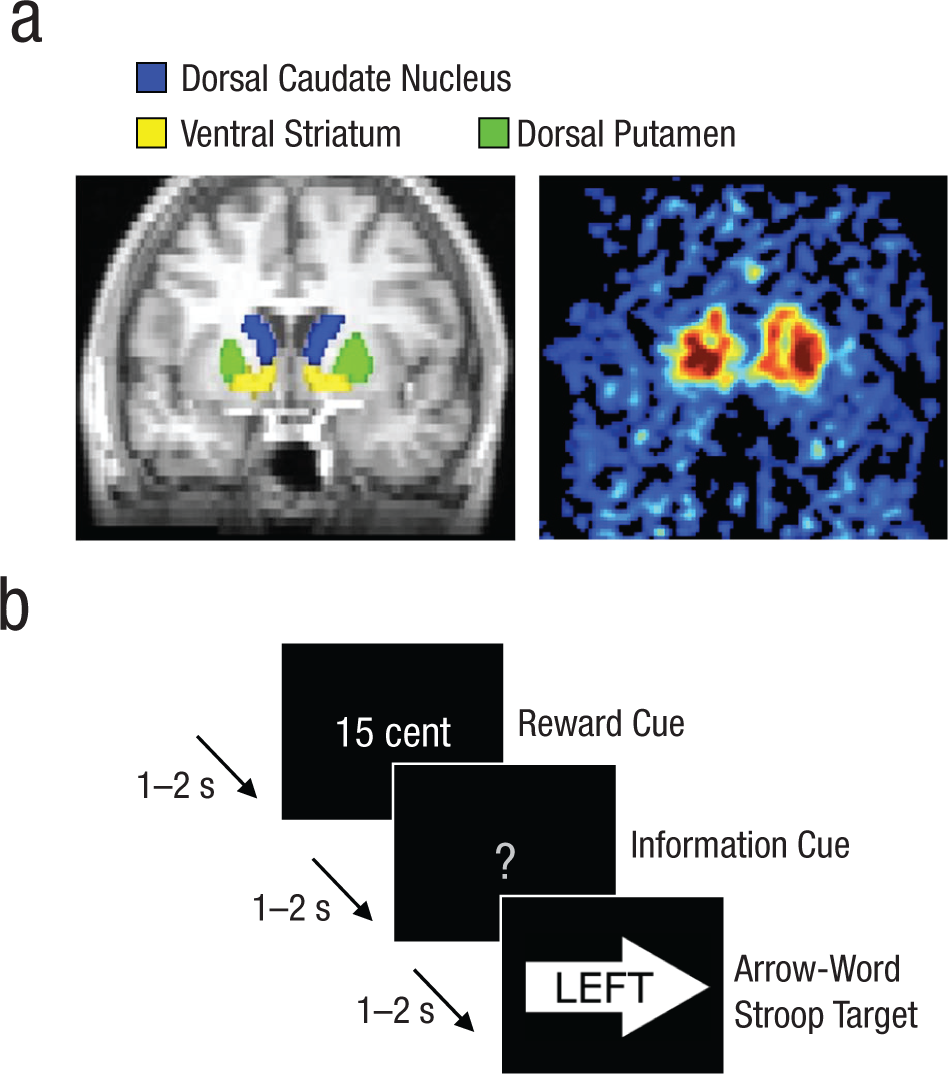
Illustration of the experimental method. Six striatal regions of interest (ROIs) were drawn on an anatomical magnetic resonance (MR) image: left and right dorsal caudate nucleus, left and right ventral striatum, and left and right dorsal putamen (a, left). Each MR image was subsequently coregistered to a positron-emission tomography image (a, right), and binding-potential values were extracted from the ROIs. The images here are examples chosen from among those used in the study. Sometime later, participants performed a rewarded Stroop task, responding to each target word by pressing the response button (i.e., left or right) corresponding to the meaning of the word. They were instructed to ignore the direction of the arrow on which the target word was superimposed. Each target was preceded by a reward cue and an information cue. In the example trial illustrated here (b), the participant could earn a high reward (i.e., U.S. $0.15), and the information cue provided no information about the congruency of the upcoming target, which was incongruent with the direction of the arrow.
PET data analysis
FMT images were reconstructed with an ordered-subset expectation-maximization algorithm that was weighted by attenuation, corrected for scatter, and smoothed with a 4-mm full-width/half-maximum kernel. To correct for motion during scanning, we used SPM8 (Ashburner et al., 2010; http://www.fil.ion.ucl.ac.uk/spm/) to realign all FMT frames to the middle (12th) frame. We used FSL-FLIRT (Version 4.1.2; http://www.fmrib.ox.ac.uk/fsl/) to coregister the anatomical MRI scan to the mean image of all realigned frames in the FMT scan. We used an in-house graphical-analysis program implementing Patlak plotting (Logan, 2000; Patlak & Blasberg, 1985) to create images representing the amount of tracer accumulated in the ROIs relative to the cerebellar reference region. We extracted average binding-potential (Ki) values from the six ROIs and computed associations between regional FMT uptake (Ki values) and the measures on the task described in the next section.
Rewarded Stroop paradigm
Behavioral responses were assessed with a Stroop-like conflict task with high-reward ($0.15) and low-reward ($0.01) conditions. The task resembled a Stroop paradigm used previously (Aarts et al., 2008), except that each trial began with a reward cue, and the interstimulus intervals were smaller (1–2 s). On each trial, the word “LEFT” or “RIGHT” (relevant dimension) was presented positioned in a left- or right-pointing arrow (irrelevant dimension). Participants responded to the target word using a button box, pressing the left button with the right index finger or the right button with the right middle finger in order to indicate the meaning of the word. The direction denoted by the word was either congruent or incongruent with the direction indicated by the arrow (Fig. 1b). All trials began with a cue predicting high or low reward for correct performance. After the reward cue, an information cue (100% valid) was provided. The information cue either informed participants about the congruency of the upcoming Stroop target (informative cue) or did not (uninformative cue; Aarts et al., 2008). Informative cues were either a red cross (incongruent target) or a green circle (congruent target); the uninformative cue was a gray question mark. Reward cues, information cues, and target congruency were equally distributed across the 240 trials (duration of ~30 min).
Three practice blocks (~15 min in total) preceded the actual experiment: one to familiarize the participants with the use of the information cues (12 trials; no reward cues), one to familiarize the participants with the use of the reward cues (32 trials with immediate feedback), and one to set the initial response window (48 trials), which was the average response time (RT) for trials responded to correctly per trial type.
In the main experiment, reward was obtained only when an answer was correct and occurred within a response window determined individually for each participant. The initial response windows, based on the final practice block, were adapted throughout the main experiment: After a correct response that was on time, 25 ms was subtracted from the response window for that trial type, and after a response that was too late, 25 ms was added to the response window for that trial type. Hence, frequency of reward receipt did not vary with difficulty and was similar across participants. Participants received no reward ($0) for either errors (button presses that were incorrect) or misses (button presses that were too late). After every 30 trials, participants were informed about the amount of reward obtained in the previous block, the amount of reward that could have been obtained, the total reward earned at that point, and the number of misses and errors made in the previous block. The reward money (M = U.S. $9.33, SD = $0.67) was added to the participants’ compensation.
Behavioral data analyses
We analyzed RTs and error rates using IBM SPSS Statistics (Version 19; IBM, Armonk, NY). RTs of all trials responded to correctly were analyzed, even if the response was too late for the reward to be earned. First, we ran six different analyses of variance (ANOVAs) with within-subjects factors of reward (high, low), congruency (congruent, incongruent), and information (informed, uninformed), and one covariate of interest: FMT uptake in the left or right ventral striatum, left or right caudate nucleus, or left or right putamen. We corrected for multiple comparisons (Bonferroni-corrected α = .0042). When we found significant effects, we assessed the effects of motivation (high-reward cues, as opposed to low-reward cues) on Stroop performance (greater interference on incongruent than on congruent trials) separately for trials with informative and uninformative cues. Finally, we assessed the effects of motivation on performance on congruent and incongruent trials separately. Post hoc simple Pearson’s correlations were calculated between FMT uptake and these behavioral measures.
In a supplementary analysis, the six covariates were put together in one model that assessed unique contributions of FMT uptake in each ROI while correcting for the effects in the other ROIs. Moreover, we performed a multiple regression analysis to demonstrate that the observed effects could not be attributed either to the time between PET and behavioral measurements or to participants’ age.
Results
Participants performed more poorly on incongruent than on congruent trials of the rewarded Stroop paradigm—RT: F(1, 13) = 66.47, p < .001; errors: F(1, 13) = 50.23, p < .001. They also performed more poorly when uninformed than when informed about the upcoming congruency—RT: F(1, 13) = 19.02, p = .001; errors: F(1, 13) = 3.95, p = .068). There was no main effect of reward—RT and errors: F(1, 13) < 1 (Table 1).
Mean Response Times and Error Rates in the Rewarded Stroop Paradigm
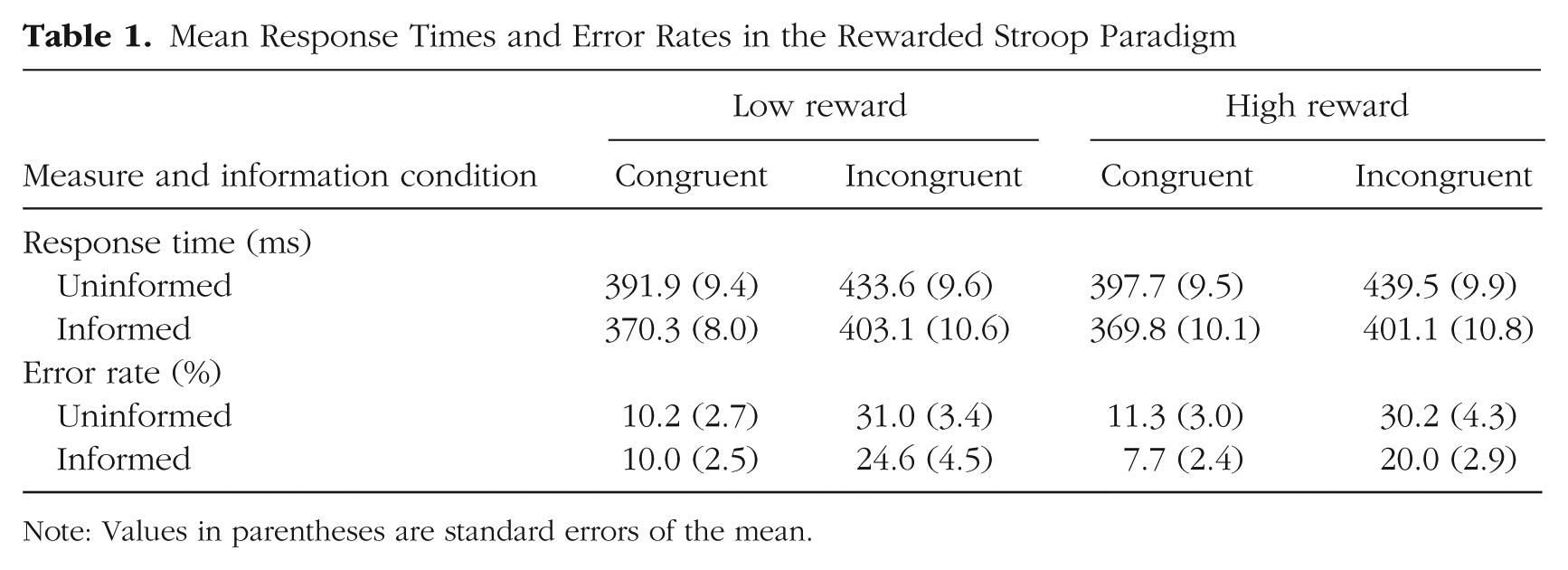
Note: Values in parentheses are standard errors of the mean.
The effects of promised reward on Stroop performance depended on individual differences in dopamine-synthesis capacity, specifically in the left caudate nucleus, as indexed by the degree of FMT uptake. The interaction of FMT uptake in the left caudate nucleus, reward, congruency, and information had a significant effect on RT, F(1, 12) = 12.8, p = .004. This interaction was significant after Bonferroni correction for multiple striatal subregions and was driven by the trials with uninformative cues, which showed a significant interaction of reward, congruency, and FMT uptake in the left caudate nucleus, F(1, 12) = 15.3, p = .002. Thus, greater dopamine-synthesis capacity was associated with enhanced Stroop interference (more interference on incongruent than on congruent trials) when participants anticipated high rather than low reward (r = .75; Fig. 2): The correlation was still significant, r = .57, t(11) = 2.3, p = .04, when the participant with the least FMT uptake and the least detrimental effect of reward (lower left data point in the figure) was excluded from the analysis. This participant’s values for FMT uptake in the left caudate nucleus and the reaction time effect were not outliers according to the Grubbs test (Barnett & Lewis, 1994); that is, they did not differ from the mean by more than 2.51 SD. This effect was present only when participants were uninformed and could not prepare for the type of cognitive control (more focus vs. less focus) that was required by the target (incongruent vs. congruent).
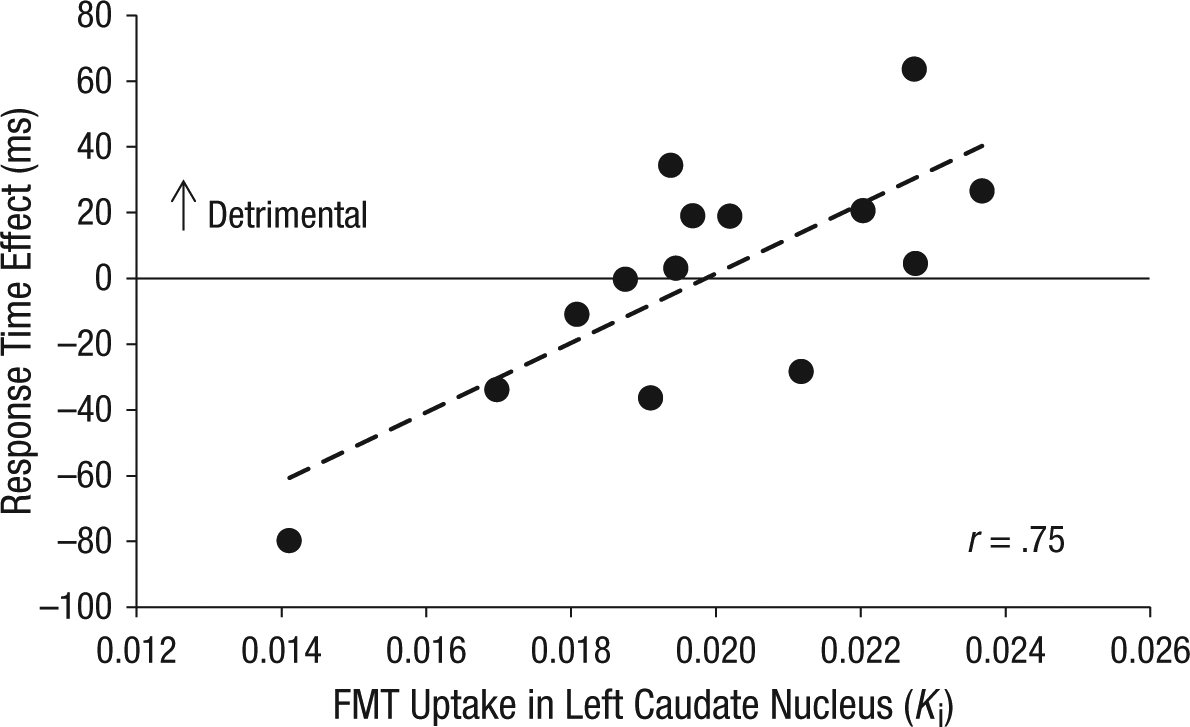
Detrimental dopamine-dependent effects of reward on cognitive control when participants were uninformed about the congruency of the target word. This scatter plot (with best-fitting regression line) shows the difference in Stroop interference between high- and low-reward trials as a function of dopamine-synthesis capacity in the left caudate nucleus. Stroop interference was measured by subtracting mean response time (RT) on congruent trials from mean RT on incongruent trials. Dopamine-synthesis capacity was measured as 6-[18F]fluoro-
The interaction of reward and FMT uptake in the left caudate nucleus had different effects on RT in the congruent and incongruent trials. Increased dopamine-synthesis capacity predicted beneficial effects of reward on congruent trials, F(1, 12) = 9.12, p = .011, r = −.66, whereas it tended to predict detrimental effects of reward on incongruent trials, F(1, 12) = 3.81, p = .075, r = .49.
We observed no main effects of FMT uptake on speed or accuracy, and no two-way interactions between FMT uptake and congruency. Moreover, analyses of RT revealed no significant effects of FMT uptake in other striatal ROIs. The unique effect of FMT uptake in the left caudate nucleus was confirmed in a supplementary analysis including FMT uptake in all six ROIs as covariates (Table 2; Table 2 also reports the results of the ANOVA with FMT uptake as a single covariate). Analyses of error rates revealed no significant effects of FMT uptake in any ROI.
Interaction Effects of 6-[18F]fluoro-
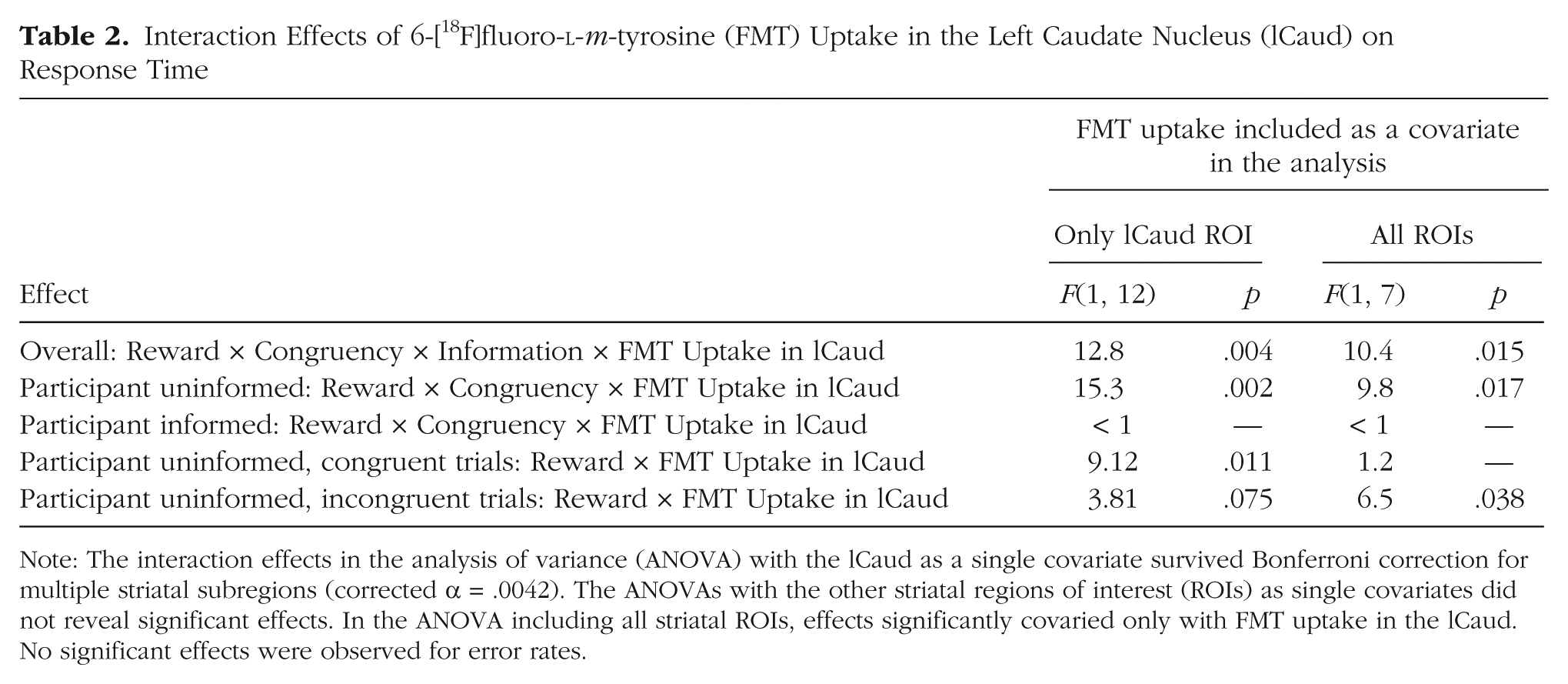
Note: The interaction effects in the analysis of variance (ANOVA) with the lCaud as a single covariate survived Bonferroni correction for multiple striatal subregions (corrected α = .0042). The ANOVAs with the other striatal regions of interest (ROIs) as single covariates did not reveal significant effects. In the ANOVA including all striatal ROIs, effects significantly covaried only with FMT uptake in the lCaud. No significant effects were observed for error rates.
A multiple regression analysis confirmed a significant interaction between FMT uptake in the left caudate nucleus and the effect of reward on Stroop interference during uninformative trials, β = 0.88, t(10) = 3.34, p = .008. Moreover, this analysis showed that the time between PET and behavioral measurements did not explain the observed effect of reward on Stroop interference, β = 0.22, t(10) = 0.76, p = .46, and neither did participants’ age, β = −0.24, t(10) = −1.07, p = .31.
Discussion
The present data not only demonstrate that appetitive motivation can have detrimental effects for cognitive control, depending on individual differences, but also provide a mechanistic account of these effects. Specifically, in people with a high baseline capacity for dopamine synthesis, a promised bonus might “overdose” the dopaminergic system, thereby impairing rather than improving cognitive control (Cools & D’Esposito, 2011). We speculate that this effect reflects motivation-induced increases in striatal dopamine that amplify the direct go pathway and inhibit the indirect no-go pathway in the striatum, resulting in an overall go bias and a “gating in” of cognitive representations (Hazy, Frank, & O’Reilly, 2006). Such gating in of multiple representations would lead to processing of both relevant and irrelevant features of a stimulus, thereby causing unselective attention and, in turn, impaired Stroop performance. In contrast, increased dopamine processing caused by anticipated reward would lead to more optimal cognitive control in people with low baseline dopamine levels, in accordance with the inverted-U shape of the function relating cognitive control to dopamine level (Cools & D’Esposito, 2011).
The dopamine-synthesis capacity of the left dorsal caudate nucleus uniquely predicted the observed effects (see Table 2), in line with previously observed interactions between motivation and cognition in the same region, which were moderated by variation in a dopaminergic gene (Aarts et al., 2010). One important implication of the present results is that greater motivation does not necessarily contribute to greater cognitive control.
Our results are remarkable in the context of the intuition that increased motivation should help people attain their goals. However, psychologists have long recognized that behavior is motivated not only by the goals that people set for themselves, but also by generalized drives that do not necessarily contribute to adaptive, optimized behavior (Dayan, Niv, Seymour, & Daw, 2006; Dickinson & Balleine, 2002). Stimuli that elicit motivation can modify behavior without accessing goal representations (i.e., in a manner that is not goal directed; Aarts, van Holstein, & Cools, 2011). This is illustrated most clearly by the role of reward-predictive stimuli in the (dopamine-mediated) compulsive seeking and taking of drugs of abuse or other targets of addiction, despite negative consequences (Everitt et al., 2008; Robinson & Berridge, 2008; Volkow et al., 2006). In the context of addiction, it is perhaps not surprising that individual differences in striatal dopamine-synthesis capacity are related to the large individual differences in the effects of appetitive motivation on cognition.
Footnotes
Acknowledgements
We thank Mieke van Holstein for pilot-testing the experiment.
Declaration of Conflicting Interests
R. Cools is a consultant for both Abbott Laboratories and Pfizer but is not an employee or stock shareholder. The other authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by the Niels Stensen foundation (a stipend to E. Aarts) and by the National Institutes of Health (Grant R01-DA20600 to M. D’Esposito, Grant F32-DA027684 to D. L. Wallace, and Grant AG044292 to W. J. Jagust). R. Cools is supported by a James McDonnell scholar award.