
Abstract
Transhumant pastoralism has played a central role in shaping and managing Mediterranean mountain landscapes. Together with specialized agricultural and sylvicultural practices it has played a pivotal role in transforming the ecology of environmental systems over time: from plant and animal populations, up to the physical-chemical properties and structural characteristics of the soils. This contribution aims to identify and characterize agro-sylvo-pastoral practices and related environmental effects at Monte Mongioie (SW Alps, Italy), a summer pastureland historically affected by transhumant livestock from the Tyrrhenian coast and the Po valley. High-resolution, multi-proxy research is presented, combining pedoanthracology, palynology, archaeobotany, archaeoentomology, malacology, biomolecular analysis, soil micromorphology, and chemistry with radiocarbon dating and historical ecology observations. This evidence has been cross-checked with documentary and iconographic sources (i.e. historical texts and maps) to unravel the complex historical practices related to the use and management of grazing resources. The integrated analysis revealed a well-preserved Roman to Medieval paleosol with high organic content, rapidly buried by a Late Medieval colluvial event that sealed the underlying features. The analyzed assemblages document persistent open grassland landscapes from the Roman period onward, maintained through sheep/goat grazing, temporary cultivation, and controlled fire use for vegetation management within a commons regime system. Progressive woodland encroachment from the 19th–20th centuries reflects the gradual abandonment of historical agro-sylvo-pastoral practices. This research highlights the high potential of dry off-sites as archeological and environmental archives when analyzed through a multi-proxy approach and a regressive, high-resolution analysis of all the different evidence unearthed.
Keywords
Introduction
Transhumance has played a pivotal role in shaping Mediterranean rural landscapes, serving as a key factor that profoundly influenced the ecology of environmental systems: from plant and animal populations to the physical and chemical composition of the soil (Bunce et al., 2004; Costello and Svensson, 2018; Saatkamp et al., 2021; Tzortzis and Delestre, 2010). The seasonal movement of livestock has also allowed for the occupation and creation of new settled spaces, fostering the exchange and circulation of knowledge and practices at local, regional, and supra-regional levels. In doing so, transhumance contributed to the formation of complex social networks and relationships. In the last few decades, landscape archeology and mountain archeology have been investigating the legacy of these environmental and historical processes involving pastoralism across the Mediterranean (Galop, 1998; Gassiot Ballbè et al., 2017; Gassiot and Jiménez, 2006; Obea Gómez, 2024; Palet et al., 2017; Pescini et al., 2024; Rendu, 2003; Rendu et al., 2016).
In this sense, the Alps has been a key region for studying the complex processes of human-environment interactions (Angelucci and Carrer, 2015; Burri et al., 2018; Carrer, 2013; Carrer et al., 2013; Gilck and Poschlod, 2019; Schwörer et al., 2014; Segard, 2009; Walsh et al., 2007, 2014, 2023; Walsh and Giguet-Covex, 2020). Many areas, however, remain largely unexplored, such as the SW Italian Alps, and more specifically, the Ligurian Alps. Indeed, only limited information is currently available for this mountain region. Ethnographic and ethnoarchaeological studies in the Maudagna Valley (Cuneo) have explored the spatial organization of pastoral huts (Carrer, 2017) and challenged linear interpretations of mountain land-use as stable and solely adaptive (Carrer et al., 2020). Historical ecology and geography have also been applied to investigate foodways and cheese-making practices in the same area (Cevasco, 2010; Cevasco et al., 1999). Paleoecological and palynological analyses of peat bog sequences in the Ellero and Carnino valleys have shed light on postglacial vegetation histories in Piedmont (De Beaulieu, 1977; Ortu et al., 2003, 2008). Further south, in the upper Argentina Valley (Foresta Demaniale di Gerbonte), dendrochronological and historical-ecological studies have examined environmental change in larch wood pastures (Cevasco and Gabellieri, 2022; Parola, 2012). Extensive archival research (drawing on medieval charters, by-laws, notarial acts, and cartographic sources) has revealed the complex governance of commons and agro-sylvo-pastoral practices (Lasalle and Palmero, 2010; Palmero, 2009; Palmero et al., 2007). Despite this growing body of research, the absence of environmental archeology investigations in the Italian sector of the SW Alps remains a significant gap in the research, especially when compared to the more advanced investigations conducted on the SW French side (Battentier, 2022; Brisset et al., 2012; Gamba et al., 2024; Mocci et al., 2005, 2008; Palet et al., 2003; Segard et al., 2003; Walsh et al., 2010; Walsh and Mocci, 2016).
The present study aims to begin filling this gap by applying an off-site multi-proxy approach to an historically important transhumance area located on the southern slope of Monte Mongioie and denominated Pian Rosso (Figure 1). Combining pedoanthracology, palynology, archaeobotany, archaeoentomology, malacology, biomolecular analysis, soil micromorphology and chemistry with radiocarbon dating, historical ecology observations and historical documents (i.e. historical texts and maps) we will unravel the complex methods of pastoral resource management. This will also include historical variations in vegetal cover and the resultant effects on soils and sediments that have contributed to the historical formation and transformation of the current mountain landscape.
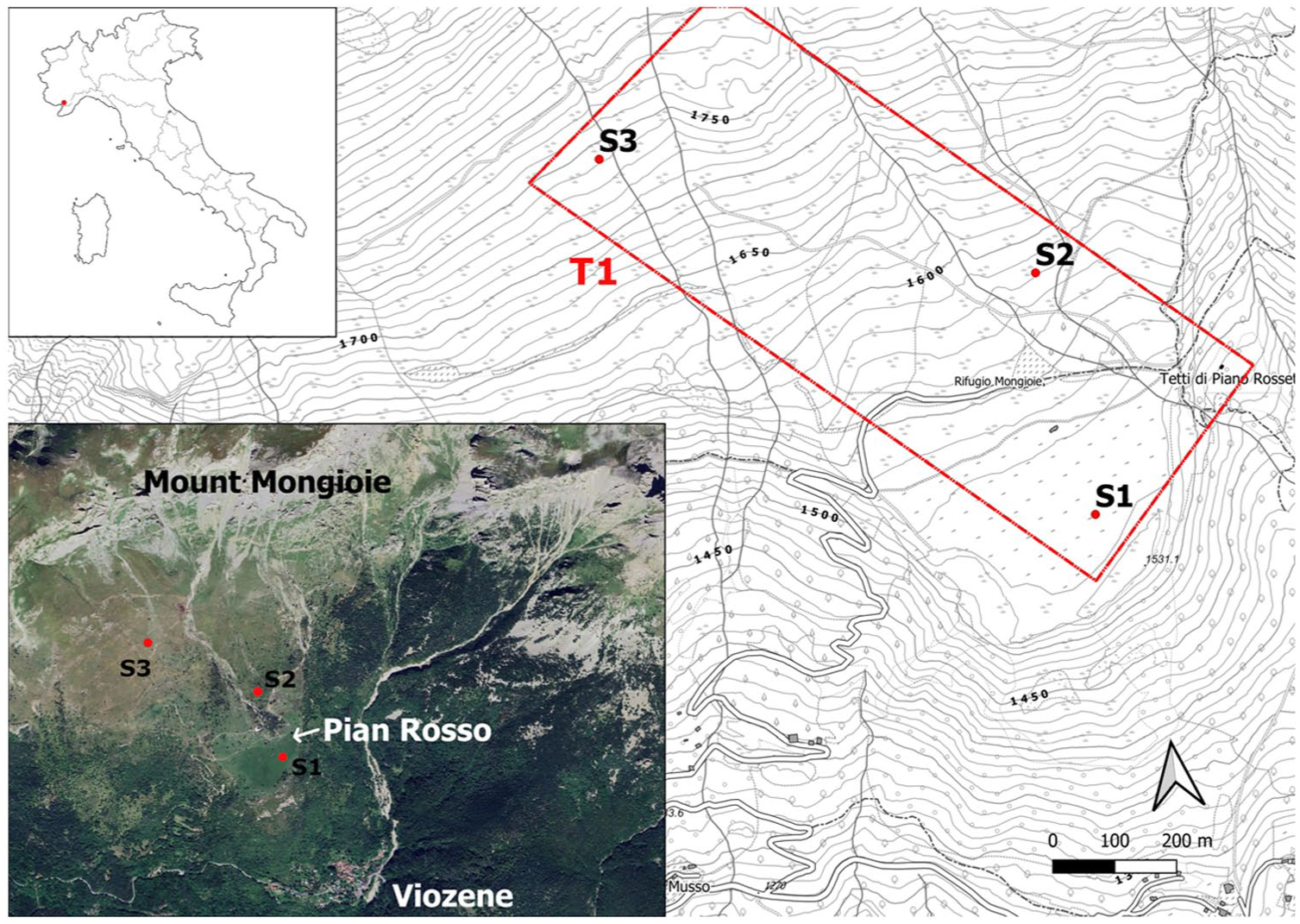
Study area location and orthophoto of Pian Rosso site. T1 refers to the altitudinal transect where the test pits (S1, S2, and S3) were excavated.
Materials and methods
Study area
Monte Mongioie (2631 m a.s.l.) is located in the Ligurian Alps, in the province of Cuneo, between the upper Tanaro Valley and the Ellero Valley. It is primarily composed of sedimentary limestone and dolomitic rocks, dating back to the Jurassic-Cretaceous period (San Pietro dei Monti dolostones and Val Tanarello or Costa Losera limestones; Bertok et al., 2011; Cortesogno and Vanossi, 1984). Karst phenomena are a defining feature of the region, with the presence of sinkholes, swallow holes, and caves formed by the dissolving action of water on limestone. Among these features, Pian Rosso, located at the foot of Monte Mongioie, at 1500 m a.s.l., has been interpreted as a paleosinkhole, formed at the geological contact between the limestone-dolomitic rocks and metasedimentary rocks (Melogno porphyroids). This is namely the remnant of the collapse of a large underground karst cavity, whose original boundaries likely extended much further south than the current edge of Pian Rosso (Walter et al., 2004).
The elliptical shaped plan shows a maximum diameter of about 600 m and a minimum diameter of about 300 m. Toward the NNW this sub-horizontal surface gently joins the slope while on the other sides it is delimited by steeper slopes; in particular, a wide alluvial fan is built on its surface at the mouth of the small gorge. It consists of individual debris flow lobes composed of clast-supported material with scarce fine-sediment matrix at its apex, grading to finer sediments toward the distal fan areas on the Pian Rosso surface. This deposit is followed by continuous coarse-grained stratified-slope deposit (scree) which is the result of erosive cycles during late glacial stages. Terracing and embankments characterize the area surrounding the Pian Rosso plateau, located between c. 1550 and 1650 m a.s.l.
The site experiences a cold climate but with mitigating effects generated by proximity to the Mediterranean coast; average annual temperatures range from 3.0°C to 5.9°C and annual precipitation is between 1001 and 1500 mm (Agrillo and Bonati, 2013; Fratianni and Acquaotta, 2017).
The study area lacks detailed floristic surveys or agronomic studies at a local scale. In 2016, a vegetation map was produced for territorial forestry planning based on photointerpretation of the overall tree cover (see Camerano et al., 2008 for details on the ‘forest’ types described in the map).
According to this map, ‘forests’ dominated by upright hooked pine (Pinus uncinata Mill.) – typically in pure stands, though sometimes mixed with larch (Larix decidua Mill.) or Scots pine (Pinus sylvestris L.) – are found at altitudes above 1500 m a.s.l. Below this elevation, ‘forests’ are primarily composed of beeches (Fagus sylvatica L.) and chestnut (Castanea sativa Mill.), the latter originating from abandoned cultivation. These ‘forests’ are interspersed with colonizing tree populations consisting of various broadleaf species, such as whitebeam (Sorbus aria L.), as well as mixed stands dominated by mountain maple (Acer pseudoplatanus L.), common ash (Fraxinus excelsior L.), linden (Tilia cordata Mill.), and mountain elm (Ulmus glabra Huds.), particularly near watercourses. The slope descending from the base of Monte Mongioie to the Pian Rosso plateau consists mainly of open areas interspersed with secondary woodlands of hooked pine, whitebeam, and ash-colored broom (Genista cinerea (Vill.) DC). This area remains one of the last active pasturelands, used for cattle grazing from June to September for cheese production. In the 1960–1970s these grasslands of Mongioie, Marguareis and Antoroto located below the rock cliff were classified according to the principle of the ‘dynamic series’ of the Grenoble school, at the scale 1:100,000, as ‘subalpine plain grasslands on superficial calcareous soils’ (Barbero et al., 1973; Bono and Barbero, 1976). In the most recent phytosociological cartography – scale 1:500,000 – the Pian Rosso pastures are included in the ‘western Alpine geosigmetum of the primary vegetation at high altitude’ – alpine geographical sector (Blasi, 2010).
More detailed information is found following the classifications adopted for evaluating the grazing capacity of these slopes (Cavallero et al., 2007). The composition of these pastures is classified into the following types, as located on the 1:10.000 map attached to the publication: a. Bromus erectus type – on moderately evolved soils (Pian Rosso, c.1500–1530 m); b. Rumex alpinus type (nitrophilic formations), 1500–1550 m (a patch); c. Brachypodium rupestre type – on evolved soils with xerophilous and mesoxerophilous facies (Pian Rosso, Arpetta, Martinelle, c. 1530–1850 m); d. Helictotrichon parlatorei type (xerophilous facies) 1850–2050 m; Sesleria varia (Jacq.) Wettst. and Carex humilis Leyss. (xerophilous facies) 1650–2050 m; Carex sempervirens Vill., Festuca gr. ovina, Sesleria varia (mesoxerophilous facies) 2050–2100 m; e. Plantago atrata Hoppe, Polygonum viviparum L., Festuca gr. rubra and Festuca gr. violacea type (mesophilous facies) 2050–2100 m (head of the Vastera stream).
In 2021, a floristic survey was conducted in the framework of this project on these open pasturelands along an altitudinal transect between 1530 and 1700 m a.s.l. Around the Pian Rosso plain the survey confirmed the presence of various herbaceous formations dominated by Gramineae (Carex sempervirens Vill., Brachypodium cfr caespitosum (Host) Roem. & Schult, Festuca ovina L., Bromus erectus Huds.) with species of mown and fertilized meadows (such as Trifolium pratense L., Trifolium repens L., Lathyrus pratensis L., Taraxacum officinale L., Tragopogon pratense L.). On narrow lynchets along the slope, nitrophilous vegetation with Chenopodium bonus-henricus L., Rumex alpinus L., Urtica dioica L., etc. Going up the slope an increase in species of pastures on limestone (such as Teucrium montanum L., Anthyllis vulneraria L., Gymnadenia conopsea L., Hieracium villosum Jacq., Phyteuma orbiculare L., Gentiana verna L., Myosotis alpestris F.W.Schmidt, Hieracium piloselloides Vill., Scutellaria alpina L.). The current coverage reflects a mosaic of xerophilous and mesophilous facies, confirming the types established by Cavallero et al. (2007).
A list of identified species at different sampling points is provided in the Supplementary Material 1, available online, along with their potential links to past land use and agro-sylvo-pastoral practices.
Historical background
Although the recovery of sporadic finds (Ravotto, 2004, 2006) near Pian Rosso suggest some form of human activity took place here during the Roman period, the lack of dedicated archeological investigations limits our understanding of this area in prehistory and Antiquity. By contrast, the medieval and post-medieval periods are better documented through written and cartographic sources, which provide detailed insights into the organization and use of these highland landscapes.
From at least the 13th to 19th century, this mountain region was part of the area known as Viozena, one of the most significant transhumant pastoral zones in the upper Tanaro Valley. Disputes over its control and use generated a substantial body of documentation (both textual and cartographic; Quaini, 1986), although this material has so far been examined primarily from jurisdictional and political perspectives (Costa, 1985; Palmero, 1999; Rosa, 2019a, 2019b). Despite ongoing armed conflicts throughout the post-medieval period, driven first by local seigniorial interests and later by community-level policies (Vester, 1997), an important shift occurred in the 18th century when the Viozena alps became the focus of an international dispute between the Kingdom of Sardinia and the Republic of Genoa, both claiming jurisdiction over these highland pastures. Information extracted from these sources enables a partial reconstruction of the systems for environmental resource use and the legal framework within which local communities operated. The most detailed documentary source concerning the Viozena’s pasturelands is the Capitula Viozene, a by-law specifically edited to clarify and regulate the management system and the rights of access to this environmental resource. This by-law was likely composed in the late 14th century but preserved only through later versions dating from the 16th to 18th centuries (Rosa, 2019a, 2019b). At that time, the Viozena area comprised seven alpine pastures, a common land jointly used by the upland communities of Ormea and Pieve di Teco. Among these, the Alpe di Pian Rosso is already mentioned in agreements from 1226, 1340, and 1429, which established shared rules for pasture allocation (Palmero, 1999). The Capitula set out the timing, conditions, and rights governing the use of pastoral resources and appointed a specific official (the gastaldo) responsible for overseeing pasture and agricultural management. Specifically, the inhabitants of Ormea were granted rights to use the Viozena pastures during the winter months (from mid-October to mid-April), while the communities of Pieve di Teco had access in the summer. These regulations indicate that the mountain pastures were not permanently divided between cultivated and grazed areas. Instead, the grazing of various livestock species (cattle, pigs, goat but mostly sheep) was regulated through a multi-year cycle of the temporary sowing of small fields in the common Alpe property, which at the end of the concession (i.e. end of the agricultural cycle) were converted back into common pasture. Until the early 19th century, grazing took place across both collectively and temporarily private held lands, following clearly established seasonal calendars. Plots reserved for the cultivation of legumes, wheat and minor cereal varieties (rye, barley, oats) were relatively small, typically defined in medieval by-laws in terms of the area a man could hoe, or an ox could plow, in a day (Barelli et al., 1907). These cultivated parcels, known as sciorte (a vernacular adaptation of the Latin sortes), were leased for multi-year periods and used for temporary agriculture employing fire techniques (ronko-runko dial.) (Barelli et al., 1907; Moreno, 2018; Quaini, 1986). In the medieval and post-medieval periods, the Alpe della Viozena was managed through a multiple-use land system in which the same plot, depending on the season and access right, was used for grazing, cereal cultivation, and sylvicultural activities focused on both timber and fodder production. Until the late 18th century, this common land-use system was the primary driver of local environmental dynamics.
A major shift occurred in the 19th century, when permanent settlement began to develop at the foot of Monte Mongioie (corresponding to the present-day hamlet of Viozene), and the area transitioned to a predominantly agricultural regime based on permanent field cultivation. This system, however, began to collapse over the course of the 20th century due to the progressive depopulation of the area.
Field sampling/fieldwork
Soil and sediment sampling was conducted along an altitudinal transect (T1; Figure 1) on the southern slope of Monte Mongioie, at elevations ranging from 1742 to 1530 m a.s.l. The selection of test pit locations was based on geomorphological and historical criteria: (1) the presence of local plateaus, increasing the likelihood of finding well-preserved pedosedimentary archives; (2) a ground surface free of tree and shrub vegetation to prevent mixing effects caused by root activity; (3) historical evidence, including texts and maps from the 16th to 18th centuries, indicating varying land use along the altitudinal transect, making it valuable to assess the effectiveness of archeological-environmental analyses in detecting land-use changes. A total of three test pits (S1-S2-S3) of varying depths and dimensions (approximately 1.5 × 1.5 m) were excavated manually. Excavation continued until reaching either the rocky substrate (whether intact or weathered) or, in its absence, an equivalent C horizon. The GPS location of each pit was recorded, along with detailed notes and photographs documenting its immediate environment. Each of the three reference pits was described in terms of morphological features and sampled. The accurate and reliable descriptions with the analytical results enabled the full characterization of all the soil horizons and the classification of the soil type according to the World Reference Base (WRB) classification (FAO, 2022).
Various amounts of soil material were collected for multi-proxy analysis using methods from related disciplines.
A total of 14 samples were collected (3–5 kg minimum/sample) from each horizon for pedoanthracological analysis (Cunill et al., 2013); when the horizon was particularly thick, sampling was carried out approximately every 10 cm to develop a more precise representation of the entire profile. A total of 27 samples were collected every 5 cm (200/300 g minimum/sample) for palynological, Non-Pollen Palynomorphs (NPPs) and lipid analysis. Collection of all samples took place from the bottom to the top of the profile to avoid contaminating the underlying material.
A 1 kg soil sample was also collected from each horizon to undergo complete laboratory analysis to characterize the major physical and chemical features (see below). Moreover, eight large, oriented monoliths (15/20 × 10 × 5 cm) were collected at different depths for micromorphological analysis. The undisturbed samples were outlined by means of a trowel and knife and, before its removal, plaster wraps were applied in order to stabilize it.
Laboratory analysis/protocols
Radiocarbon
A total of eight AMS radiocarbon dates were obtained at the Radiocarbon Dating Laboratory of the University of Lund (n = 3) and the Poznan Radiocarbon Laboratory (n = 5). This included five dates from T1S1, one date from T1S2 and two dates from T1S3. All samples were cleaned using an acid-base-acid (ABA) pre-treatment protocol. All results have been corrected for isotopic fractionation according to the conventions of Stuiver and Polach (1977), with δ13C values measured on prepared graphite using the AMS spectrometer. The 14C dates were calibrated in calendar years with OxCal Ver. 4.4.4 software (Bronk Ramsey, 2021) based on the IntCal 20 database (Reimer et al., 2020). Charcoal fragments measuring ⩾ 1 mm, collected directly from the test pit excavation and at different depths, were selected for dating. In the case of test pit S1, which exhibited considerable depth, additional charred material (i.e. charcoals and seeds) was extracted in the laboratory to provide a more comprehensive chronological framework for the entire profile. Results are shown in Table 1 together with taxa identification and further information on the dated charred material.
List of the radiocarbon dating (AMS 14C dating) with information on the type of the dated material and taxa identification.
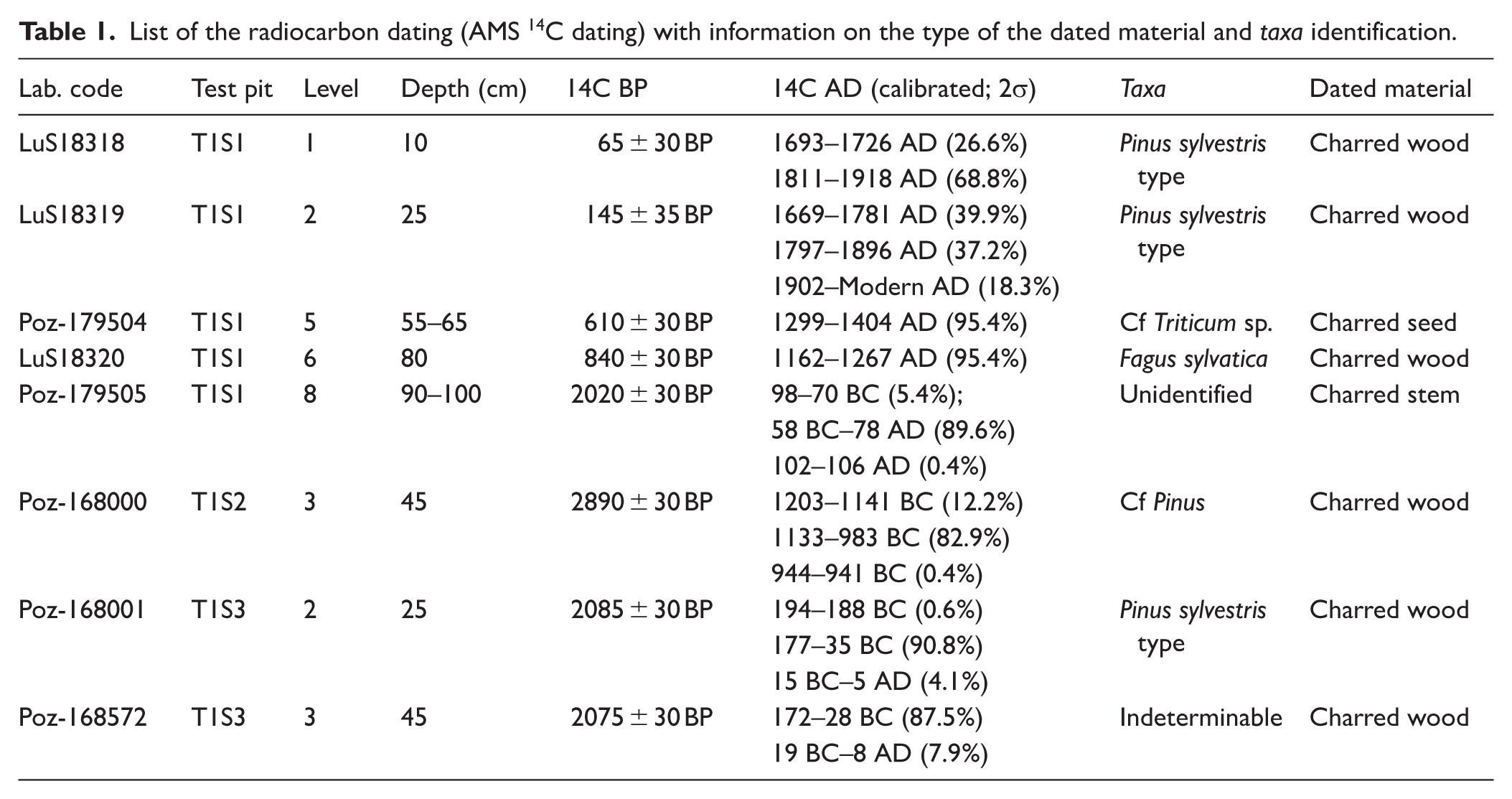
Despite ongoing debate surrounding the reliability of radiocarbon dating of charred remains (i.e. charcoals, seeds) in pedogenetic horizons (due to potential mechanical disturbances such as tree uprooting, bioturbation, or agro-sylvo-pastoral activities) this method has demonstrated sufficient robustness, as evidenced by multiple studies (Knapp et al., 2013; Nelle et al., 2013; Saulnier et al., 2020). Although a precise chronological attribution of the horizons in these types of contexts should always be approached with caution, the designation of each horizon was strengthened by the consistent integration of multiple indicators (i.e. pedological dynamics, pedo and bioturbation, bioarchaeological proxies’ distribution). Age-depth models have also been created to further assist chronological interpretation (See Supplemental Material 2, available online).
Soil analysis and micromorphology
The laboratory soil analyses were performed by the Regional Soil Analysis Laboratory in Sarzana (Spezia, Liguria; ISO 9001 certified). All soil samples from each horizon, were subject to the following routine laboratory procedures and analyses, performed according to the Ministero delle Politiche Agricole e Forestali (2000): the soil samples were air dried; particle size distribution analysis of the coarse fraction (>50 µm) was conducted via wet sieving; the composition of the fine fraction (<50 µm) was determined via the pipette procedure after dispersion of the sample with sodium hexametaphosphate, (NaPO3)6; the pH was measured by the potentiometric method in a 1:2.5 soil: water suspension; the total carbonate content was determined using the Dietrich Früling calcimeter; the active carbonate content was determined with ammonium oxalate; the cation exchange capacity (CEC) and exchangeable bases were determined with BaCl2-triethanolamine at a pH of 8.2; and the concentrations of extracted chemical elements, Total Organic Carbon (TOC) and N content were determined via flame atomic absorption spectrometry (FAAS) based on the Dumas methods (1831) by means of high-temperature (1000°C) combustion and the removal of inorganic carbon (carbonates) in the sample by pre-treatment (Kerven et al., 2000). Measurements of organic carbon can be used to estimate organic matter (OM) through the use of a conversion factor, which assumes that 58% of the OM was formed by carbon (Nelson et al., 1996).
The undisturbed samples for micromorphological analysis were dried and successively impregnated with epoxy resin under vacuum in the ‘Servizi per la Geologia’ laboratory located in Piombino (LI), Italy, following the guidelines detailed in Benyarku and Stoops (2005). Standard thin sections (55 × 85 mm) were obtained from the samples and were observed using a polarizing microscope. The thin sections were described using the terms and methods of Stoops (2020). The interpretation of micromorphological features was performed according to Stoops et al. (2018).
Palynology
From each of the palynological samples 2 cm3 of sediment was collected to be treated according to standard methods (Fægri et al., 1989; Moore et al., 1991). Testing with HCl 10% revealed a high content of calcium carbonate, particularly in the samples of the uppermost 70 cm. Lycopodium tablets were added to estimate pollen concentration (grains/cm3). Palynomorph counting was carried out operating at 400–1000×. Pollen and spores were identified referring to literature (Beug, 2004; Blackmore et al., 2003; Moore et al., 1991; Punt et al., 2003; Punt and Hoen, 2009; Reille, 1992; Van Leeuwen et al., 1988). For the identification of other palynomorphs, several papers and books were consulted (among others Ellis and Ellis, 1997, 1998; Nuñez Otaño et al., 2021; Shumilovskikh et al., 2015, 2021; Shumilovskikh and van Geel, 2020; Van Geel, 1978; van Geel and Aptroot, 2006). The nomenclature of the NPPs follows the criteria indicated in Miola (2012). Due to the very low concentration of pollen grains in the samples and their poor preservation, it was not possible to reach the standard threshold of at least 300 grains. The counts for each sample are shown in the diagram (11–128 pollen grains), and the curves of pollen types are indicative only.
The percentages of pollen types are calculated on the Total Land Pollen (TLP); in this case, Cyperaceae (max one or two grains per sample) were included in the TLP and they likely represent non-hygro-hydrophilous species of Carex. The percentages of the NPP morphotypes are calculated on their total sum (Innes and Blackford, 2003). The pollen diagram was drawn with TILIA software (version 3.0.3. ©1991–2022 Eric C. Grimm).
Pedoanthracology
The pedoanthracological analysis was conducted at the Laboratory of Bioarchaeology at the Catalan Institute of Classical Archeology (ICAC) following the standard methods (Carcaillet and Thinon, 1996; Cunill et al., 2013; Saulnier et al., 2020; Talon et al., 2002). Soil samples were air dried and then sieved to remove rock fragments larger than 5 mm; the remaining material was then weighed to calculate the anthracomass (charcoal amount) and quantitatively express the anthracological data. The dried samples were sieved with water using a stack of sieves with mesh sizes of 4, 2, and 0.8 mm. Charcoal sorting was performed using a binocular microscope (ZEISS Stemi 508) at a magnification of 5.0–0.63×. The anthracomass was calculated based on the weight of charcoal pieces larger than 0.8 mm. The results are presented in terms of the total weight of charcoal per pit expressed in mg per kg of dry sediment (AP, anthracomass per pit), of charcoal per level (AL, anthracomass per level) and of charcoal per level and per taxon (ALT, anthracomass per level and taxon). Following Saulnier et al. (2020) and the references therein, up to 50 charcoal remains were randomly selected from each size fraction in the sieve for each level, where the charcoal quantity allowed. Identification was performed using reflected light microscopy (Olympus BX51) with both bright- and dark-field settings at magnifications of 50×, 100×, 200×, and 500×. Charcoal identification was based on anatomical features described in wood anatomy atlases (Greguss, 1959; Schweingruber, 1990; Schweingruber and Berge, 2017a, 2017b, 2017c) as well as online databases (Wood Anatomy, InsideWood Database, Xylem Database, and Digital Plant Atlas). Botanical nomenclature follows The Plant List (TPL). When identification was only possible at the genus level, the abbreviation ‘sp’. was used (e.g. Pinus sp.). The designation ‘type’ reflects anatomical uncertainty among several species. If anatomical diagnostic features were too ambiguous for precise genus identification, the abbreviation ‘cf’. was prefixed to the suggested genus (e.g. cf. Fagus). Fragments that were too damaged to allow anatomical analysis (e.g. ‘vitrified’ charcoals deriving from the fusion and homogenization of cell walls, Courty et al., 2020; McParland et al., 2010) were classified as ‘indeterminate’. The term ‘unidentified’ was used for remains with largely intact anatomical structures but uncertain taxonomic classification. The term ‘other Angiosperm’ refers to angiosperm different from Fagus sylvatica. The charcoal diagram was drawn with TILIA software (version 3.0.3. ©1991–2022 Eric C. Grimm).
Archaeobotany, archaeoentomology, and malacology
During pedoanthracological sorting, other organic remains such as charred and modern seeds, insects, and terrestrial mollusks were also collected when present, and analyzed.
Archaeobotanical identifications were conducted with a binocular microscope (ZEISS Stemi 508) at a magnification from 5.0× to 0.63× and with the aid of relevant seed atlases (Cappers et al., 2012) and archaeobotanical publications (e.g. Jacomet, 2006); Latin nomenclature for all plant remains follows Guest (1966–1988); Zohary et al. (2012) for wild species.
Insect remains were analyzed at the TRACES Laboratory of the University of Toulouse Jean Jaurès II (France). Identification was carried out using a Leica MZ 125 microscope and standard reference works. The minimum number of individuals (MNI) was determined through the quantification of lateralized anatomical elements (Schafstall et al., 2020). Particular attention was paid to the state of preservation of insect cuticle, using the categorization system (Time since death index (TSD index)), based on previous studies (Kenward and Large, 1998; Morlan and Matthews, 1983) and developed in Parrilla et al. (2026). This index allows the identification of potentially more recent remains introduced through bioturbation, a common phenomenon in soils. Terrestrial mollusk identifications were conducted with a Wild stereo-microscope at magnification from 6× to 50×, with the aid of relevant atlases (Welter-Schultes, 2012) and many other publications (Gavetti et al., 2008; Kerney et al., 1983). The nomenclature is in accordance with the last checklist of Italian molluscan fauna (Bodon et al., 2021). Ecological characters were based mostly on Falkner et al. (2001). The minimum number of individuals (MNI) was calculated based on the presence of specific conchological elements (e.g. protoconch for young shells, peristome for mature specimens); many small unidentified fragments were not counted.
Lipid extraction, analysis, and quantification
Lipid extraction, identification and quantification was conducted at the AMBI Lab (Universidad de La Laguna, Spain). About 5 g of each sediment sample was dried for 48 h at 40°C and then homogenized using an agate mortar. Before extractions, all glassware and glass wool were placed in a muffle oven at 550°C for 3 h to eliminate any possible organic compounds. Protocols for the total lipid extract (TLE) and fraction separation of n-alkanes, aromatics and fecal sterols compounds via silica gel column chromatography were completed following Jambrina-Enríquez et al. (2018) and Pescini et al. (2024). Common indexes developed for aiding in the palaeoenvironmental interpretation of occurring wax n-alkane abundances were used: the odd-over-even predominance (OEP), average chain length (ACL), and P-aqueous (Paq). OEP measures the relative abundance of odd over even carbon for the evaluation of the degradation effect, whereas ACL is the weighted average of the various carbon chain lengths reflecting changes in vegetation structure through a sedimentary sequence (Bush and McInerney, 2015; Connolly et al., 2019). Paq was first proposed by Ficken et al. (2000) to separate submerged/floating aquatic macrophyte inputs from emergent and terrestrial plant inputs to sediments.
The odd-over-even predominance (OEP) of the n-alkanes patterns was applied following the Zech et al. (2010) formula: OEP = (C27 + C29 + C31 + C33)/(C26 + C28 + C30 + C32), and the Paq is defined as: Paq = (C23 + C25)/(C23 + C25 + C29 + C31). The average chain length (ACL) was calculated following Freeman and Pancost (2014) using the C21–C35 interval: ACL21–35total = Σ(Ci × [Ci])/Σ[Ci]; 21 ⩽ i ⩽ 35 (The value [Ci] is the concentration of the n-alkane with i carbon atoms).
Additionally, the distribution of n-alkanes was also evaluated using the long chain n-alkane ratio (nC31 + nC33)/(nC27 + nC29 + nC31 + nC33) to determine the percentage contribution of grasses and herbs versus trees and shrubs, to the total content of fossil plant alkanes (Schatz et al., 2011; Zech et al., 2010).
Results
Soil features and classification
T1S1 (Coordinates WGS84: Lat
44,150,221°N; Long. 7,776,903°E; Alt. 1530 m a.s.l.) – Cambic Calcisol (Siltic; Thapto-Kastanozems).
The profile is located on a slightly depressed karst plateau due to the presence of a deep sinkhole (Figure 1).
The field data defines a complex pedosedimentary sequence greater than 1 m thick (Figure 2); unfortunately, the pit was not deep enough to allow us to reach the bedrock. From a stratigraphic perspective, the sequence can be divided into two parts, or pedostratigraphic units (hereafter PSU). In fact, a main lithological discontinuity has been identified. At the bottom, a buried organic paleosol with a silty loam texture and well-developed fine prismatic structure, is observed likely due to vertic properties (PSU1; see Supplemental Material 3, available online for the main field features of this profile). It consists of an 2ABb horizon showing medium values of CaCO3 and high pH (alkaline). The 2ABb horizon shows a high O.M. and N content while P content is scarce. The CEC value is high, mostly due to the higher OC and clay content (Table 2).

Sketch of the investigated soil profiles (T1S1, T1S2, and T1S3) and related photo. The showed radiocarbon dates are calibrated.
The major physical and chemical features of the profile.
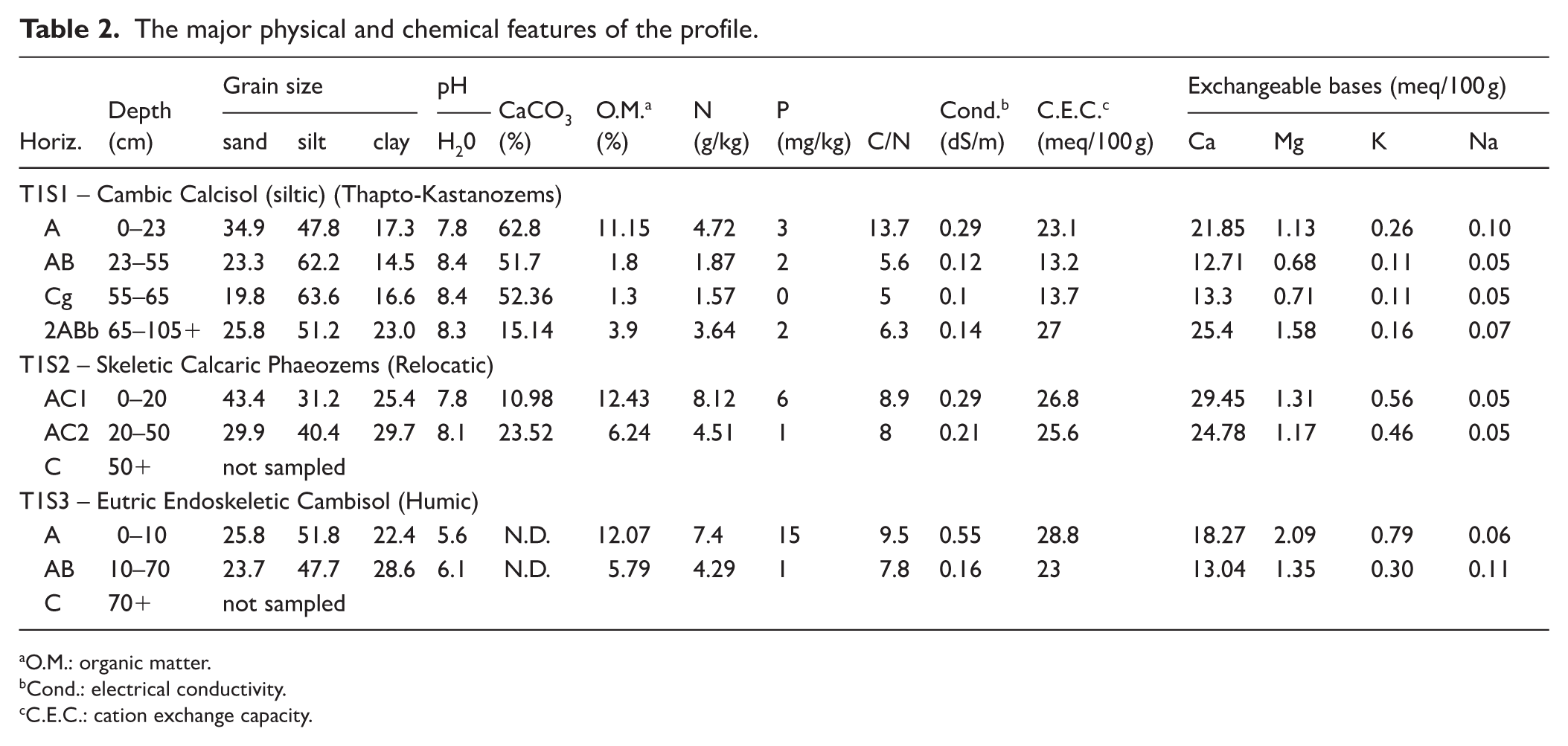
O.M.: organic matter.
Cond.: electrical conductivity.
C.E.C.: cation exchange capacity.
The paleosol is covered by a colluvial deposit (PSU2), weakly pedogenized, which is composed of silty loam reworked material from the alluvial fan and in which there is substantial secondary accumulation of CaCO3 and signs of hydromorphism (Cg) due to the presence of more clay and organic material in the buried paleosol at the base, slowing the downward flow. These horizons have medium to high available OM and N content, whereas the P content is always low (Table 2).
The micromorphological analysis of the soil horizons revealed several features that are not always recognizable at the field scale. Among the main micromorphological characteristics identified in the A horizon, clusters of micro-charcoals (Figure 3(c)) represent one of the most significant features. Particular attention was devoted to the micromorphological characteristics at the transition to the 2ABb horizon to obtain information on the nature of palaeoenvironmental changes. The Cg horizon is characterized by a chaotic mixture of reworked and rounded soil aggregates (pedorelicts) and well-sorted calcareous silty grains, likely resulting from sedimentary phases related to the alluvial fan (Figure 3(b)). Near the buried 2ABb horizon, a complex surface sedimentary crust was also identified. This crust appears locally fragmented and incorporated into the groundmass and is characterized by lamination and vesicular voids (Figure 3(a)). In some cases, scattered charcoal fragments occur within the groundmass of the 2ABb horizon (Figure 3(d)), which commonly shows channel and chamber void systems produced by soil fauna activity. This biological activity results in strongly bioturbated soil material. In particular, earthworm and enchytraeid excrements and infillings (Figure 3(l)) are present throughout all horizons of the profile. The particular environmental condition (alkalinity and moisture availability) may have favored the activity of earthworms.
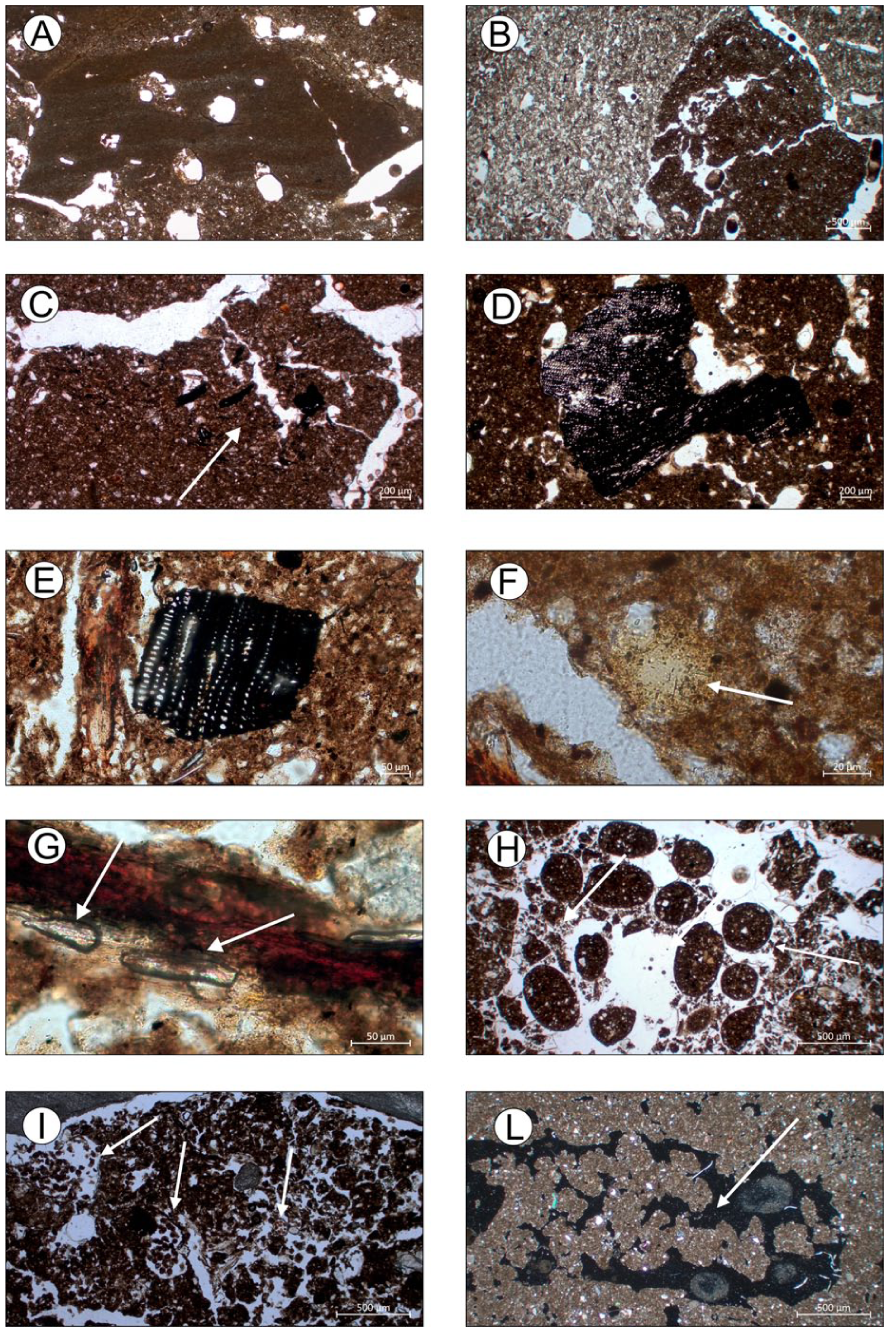
Relevant micromorphological features of pedo-sedimentary sequences of T1S1, T1S2 and T1S3. (a) Complex fragment of surface crust formed by several sedimentary microlayers (silt and clay) and some vesicles, integrated in colluvial deposit. PPL (2ABb/Cg, T1S1); (b) Chaotic mix of reworked and rounded soil aggregate (pedorelic) and well sorted calcareous silty grains, PPL (Cg, T1S1); (c) Cluster of micro-charcoals, PPL ((a) T1S1.); (d) Large fragment of charcoal (2ABb, T1S1); (e) Charcoal fragment, PPL (AC1, T1S2); (f) Small phosphatic nodule (apatite), PPL (AC2, T1S2); (g) Suspected elongate phytoliths in strong humified organic tissues, PPL (AC1, T1S2.); (h) Earthworm fecal pellets, PPL ((a) T1S3); (i) Ellipsoidal organic excrements of mites in decomposing roots and modified voids, PPL (AC1, T1S2); (l) Infillings with coalesced earthworm excrements, XPL (AB, T1S1).
T1S2 (Coordinates WGS84: Lat
44,155,278°N; Long. 7,766,774°E; Alt. 1578 m a.s.l.) – Skeletic Calcaric Phaeozems (Relocatic).
The profile is located on the apex alluvial fan characterized by terraces (Figure 1). It is 80 cm deep and exhibits horizons that are extremely gravelly with significant textural differences, but that are also rich in organic matter (Figure 2). In particular, the upper horizon is also characterized by high N content but low available P content. The pH values are slightly alkaline due to the influence of carbonates, while the high CEC values of these soils appear to be directly correlated with the organic matter content (see Table 2). These horizons are characterized by intense biological activity. Observation of thin sections revealed a very dark micromass and a granular microstructure. The aggregates are moderately to highly separated, mostly rounded. They can coalesce to create a spongy-like structure with various types of voids. Frequent plant residues and root tissues are often observed in channels. The pedofeatures observed in thin section include abundant excrement, infillings and passage features due to soil fauna activity. It is possible to recognize excrements attributable to different species of invertebrates: mites, enchytraeids and endogeic earthworms (Figure 3(i)). Excrement infillings are often the predominant organic soil matter in agricultural or cultivated soil (Kooistra and Pulleman, 2020) that is well populated with earthworms, specifically in the unploughed near surface horizon (Stolt Mark and Lindbo David, 2010). It is also possible to distinguish remains of incompletely burned vegetal tissue, phytoliths (Figure 3(g)), fine charcoal fragments (Figure 3(e)), and secondary phosphates, commonly occur as small nodules or insect remains (Figure 3(f)).
T1S3 (Coordinates WGS84: Lat
44,153,721°N; Long. 7,775,621°E; Alt. 1741 m a.s.l.) – Eutric Endoskeletic Cambisol (Humic).
The profile is 70 cm deep, and it is located on upper part of a talus slope (Figure 1) used as a pasture, characterized by the alternation of clast-supported layers and matrix-supported layers constituted of fine pre-weathered soil material.
The field data characterizes well-structured soil, with a silty loam texture and well-developed sub angular blocky structure (Figure 2). In particular, the upper horizons are characterized by complete carbonate leaching, acid reaction and high available P and N contents (see Table 2).
The results of the micromorphological analysis performed on the horizons allow us to observe a strong biogenic soil microstructure, dominated by spheroidal earthworm excrements (Figure 3(g)) and channels. Plant residues, phytoliths and roots are common. The occurrence of elongated phytoliths, very common in grasses (Poaceae), or dung accumulation together with the micromorphological and chemical features of the soil may also be indicative of a grazing area under humid conditions (Kooistra and Pulleman, 2020).
Bioarchaeological analysis
Only the results of the T1S1 survey are presented as the bioarchaeological analyses of the T1S2 and T1S3 surveys are still ongoing.
Pollen and NPPs
Overall, pollen grain preservation was poor and high percentages of indeterminable pollen grains were present, possibly due to the calcareous nature of the soil (see section on issue of taphonomy and preservation), furthermore, the dominance of Cichorioideae likely reflects the greater resistance of their pollen to degradation, which may have led to an overrepresentation in the assemblage. The percentages of pollen taxa are shown in the diagram, but due to the low TLP values these percentages should be considered indicative only. The synanthropic vegetation indicators include: Amaranthaceae, Polygonum aviculare type, Urticaceae, Sambucus ebulus type and Artemisia (Figure 4(a)).
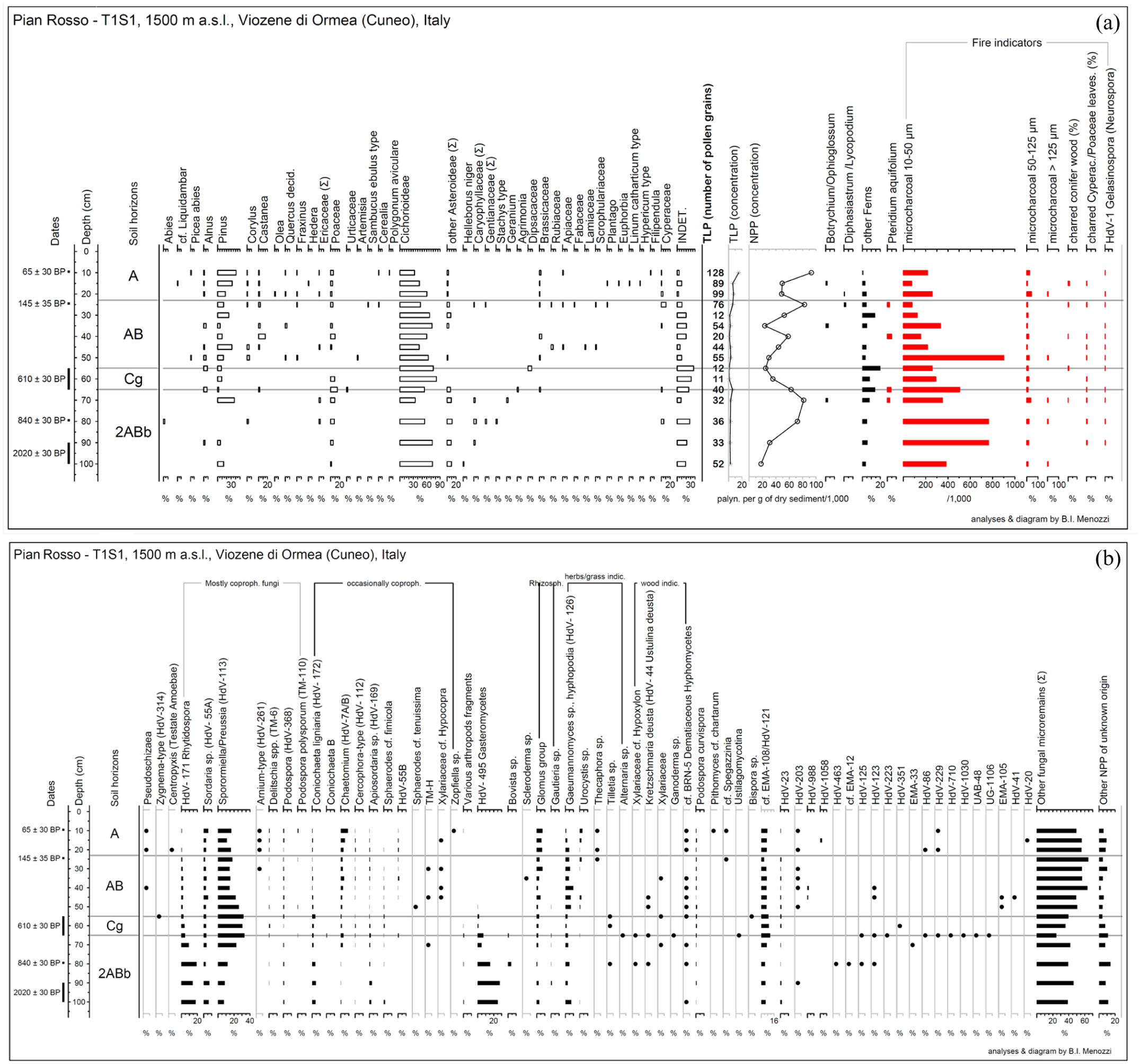
(a) Pollen diagram of T1S1; (b) non-pollen palynomorphs (NPPs) percentages diagram of T1S1.
More evidence for the history of the site are provided by the non-pollen palynomorphs, whose concentration was adequate (Figure 4(b)) with the number of palynomorphs counted ranging from 352 at 100 cm to more than a 1000 at 40 and 25 cm. The number of NPP types categorized during the counting varies between 31 (at 90 cm) and 46 (at 80 cm) and in total about 120 types have been differentiated. To facilitate the reading of the diagram the types with known ecological features and taphonomy are grouped together: (1) soil surface indicators: HdV-495 Gasteromycetes; Bovista sp.; Scleroderma sp.; Arthropods fragmented exoskeleton parts; (2) rhizosphere indicators: Glomus group and spores of the hypogeal fungal genus Gautieria (Montecchi and Sarasini, 2000; Sarasini et al., 2015); (3) herbaceous plant indicators: Tilletia type, HdV-126 Clasterosporium caricinum (Pals et al., 1980; van Geel and Aptroot, 2006); UG-1079 Urocystis; Thecaphora sp.; Alternaria sp.; (4) Wood indicators (trees and/or decaying wood): Xylariaceae cf. Hypoxylon; HdV-44 Ustulina (=Kretzschmaria) deusta (Van Geel, 1978); Ganoderma sp.; (5) the coprophilous fungi are divided in two main groups, the primarily coprophilous (Arnium, Delitschia spp. (Cugny et al., 2010; Shumilovskikh and van Geel, 2020; van Geel et al., 2003), HdV-55A, HdV-171 Rhytidospora (Doveri, 2014; van Geel and Aptroot, 2006), Podospora spp., Sporormiella/Preussia (HdV-113), TM-110 Podospora polysporum), and all the others (Coniochaeta B, HdV-7A/B Chaetomium, HdV-112 Cercophora, HdV-172 Coniochaeta; HdV-169 Apiosordaria; HdV-55B; Sphaerodes cf tenuissima; Sphaerodes cf. fimicola; TM-H; Xylariaceae cf. Hypocopra; Zopfiella).
Pteridium, HdV-1 Gelasinospora (now included in the genus Neurospora), charred fragments of monocot leaves and of conifer wood are present throughout the sequence but in minimal quantities.
Among the coprophilous fungi spores, the most abundant are ones of the type Sporormiella/Preussia (HdV-113), followed by HdV-171 Rhytidospora that occur more abundantly in the deepest levels (horizon 2ABb). The fungi known to be mostly fimicolous are more abundant in the horizon Cg and then in 2ABb.
A somewhat curious trend is shown by the soil surface indicators whose highest percentages (around 20–27%) are in the three lowest samples (horizon 2ABb, 80–100 cm); percentages that decline going up in the sequence. These spores are produced above the surface of the soil by gasteroid fungi (mainly puffballs).
Conversely, Glomus group and Gautieria sp. (rhizosphere indicators) are present mostly in the AB and A horizons.
Wood indicators reach very low percentages and are present between 35 and 80 cm, while herbaceous plant indicators appear constantly with percentages ranging between 3 and 9%.
Soil charcoal and seeds
The total anthracomass of S1 is 385,168 mg/kg corresponding to 1.226 charcoals. Charcoal fragments were found at all levels of the test pit, though with noticeable variations (Figure 5). The anthracomass reached a maximum of 329,294 mg/kg at the most superficial level (Level 1, 10–20 cm deep). From Level 2 (25–35 cm deep) onward, charcoal remains decreased significantly, except at Level 5 (55–65 cm deep), where there was a slight increase. The charcoal content then declined further, reaching a minimum of 1557 mg/kg at Level 7 (80–90 cm deep) before slightly increasing again at the deepest level (Level 8, 90–100 cm deep). The 422 charcoals identified show a very poor variety of taxa: Pinus sylvestris type, Fagus sylvatica, undetermined gymnosperms and undetermined angiosperms. Pine dominates all the analyzed samples. Within the framework of this study, this taxon may correspond to P. uncinata Ramond ex DC. and/or P. sylvestris L. Among the fragments belonging to this category, nine charred pine needles (Figure 6(a) and (b)) were also included and found in Level 1 (10–20 cm deep; n = 5), Level 2 (25–35 cm deep; n = 2) and Level 5 (55–65 cm deep; n = 2). The category of ‘unidentified’ includes charcoals whose anatomy is visible (although in some cases partially deformed i.e. through vitrification) but it has not been possible to attribute any specific taxon. Nonetheless, certain anatomical elements allow us to link some of these fragments to stems (Figure 6(c)–(f)) and rhizomes (Figure 6(g) and (h)) of herbaceous species.
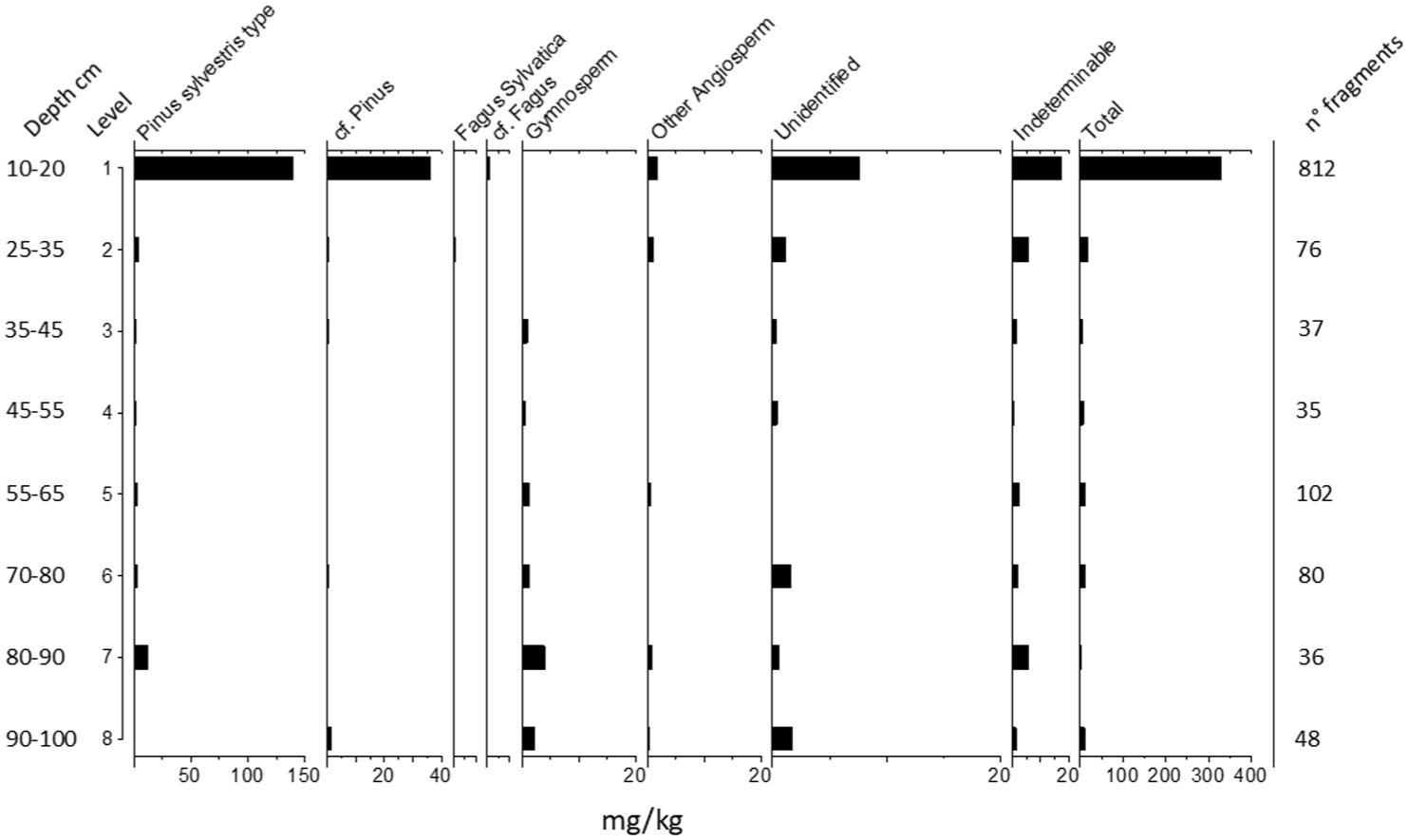
Pedoanthracological diagram, showing the anthracomass per Level and identified taxa.

Charcoal fragments recovered from T1S1 and examined using reflected light microscopy and a stereomicroscope at different magnifications. (a and b) Cross section of a charred pine needle; (c–f) ‘Unidentified’ charcoal fragments tentatively interpreted as grass culms: left, lateral view under stereomicroscope; right, transverse section observed under reflected light microscopy; (g and h) ‘Unidentified’ charcoal fragment possibly corresponding to roots or rhizomes of herbaceous plants. High-definition photos are also added in Supplemental Material 5, available online.
Overall, a total of 17 charred plant items and 75 modern plant items were recovered from the test pit samples. Modern plant remains were not all identified to the species level, but are consistent with a ruderal, pastureland environment. It is also notable that modern plant remains were only recovered from the upper 65 cm of the profile, indicating a moderate level of bioturbation in these top strata, by modern plant roots, burrowing snails and mammals. Of the 17 charred plant items recovered, seven were identified as indeterminate cereal remains and two as cf. Triticum sp. Other items were either wild/weed taxa indicative of ruderal environments or unidentifiable charred fragments (see Table 3). The preservation and overall recovery of archaeobotanical material from the test pit was poor. In the majority of cases, grains and seeds were broken and/or charred at very high temperatures (>400°C – Charles et al., 2015; Stroud et al., 2023). Charred plant remains were only recovered from the lower 45 cm of the test pit. This fact, coupled with the lack of modern plant remains found in these levels would indicate that taphonomic processes such as bioturbation and soil erosion were less of a factor in seed preservation within the deeper levels of the test pit. Finally, radiocarbon dating of the cf. Triticum grain from Level 5 (55–65 cm) indicated a date range within the 14th century AD. This date seems to fit with the results of other radiocarbon dating completed on charcoal from other levels of the test pit, with the age of remains increasing with soil depth. This could suggests that charred remains were not subject to high post-depositional soil mixing and may provide a possible representation of local vegetation through time, in this area of Monte Mongioie.
Summary of all archaeobotanical and modern plant remains recovered from profile T1S1.

Entomofauna and malacofauna
A total of 23 sclerites were found throughout the entire profile, indicating a sporadic preservation pattern (Table 4). The sclerites appear shiny and show no significant taphonomic alterations, being mostly categorized as stage 1 of TSD (Parrilla et al., 2026), except for two slightly tarnished pupae from Level 5 (classified in stage 2) and a complete Megalonotus specimen (classified in stage 0) found in Level 1. In Level 8 (90–100) the head of Myrmica sabuleti was well preserved, with both eyes and bristles (stage 1). Several remains, however, were too fragmented and lacked diagnostic features, preventing any reliable taxonomic identification.
List of entomofauna (Coleoptera and Hymenoptera) and terrestrial mollusk identified at Pian Rosso along the profile T1S1.
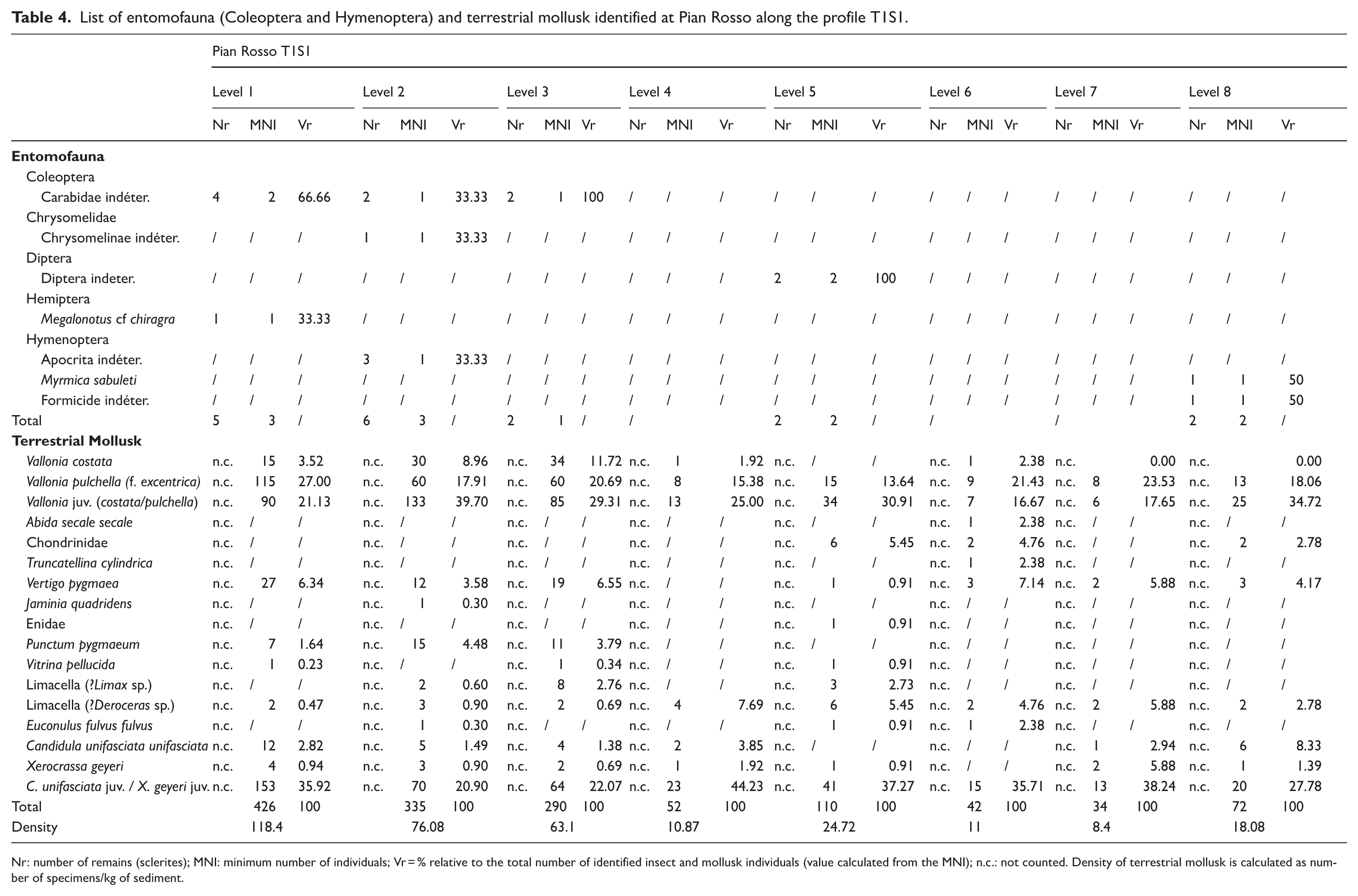
Nr: number of remains (sclerites); MNI: minimum number of individuals; Vr = % relative to the total number of identified insect and mollusk individuals (value calculated from the MNI); n.c.: not counted. Density of terrestrial mollusk is calculated as number of specimens/kg of sediment.
A total of 1361 terrestrial mollusk specimens were recovered (Table 4 and Supplemental Material 4, available online). The most abundant taxa belong to the genera Vallonia (V. costata (Müller, 1774), V. pulchella (Müller, 1774)), Vertigo (V. pygmaea (Draparnaud, 1801)), Candidula (C. unifasciata unifasciata (Poiret, 1801)), and Xerocrassa (X. geyeri (Soós, 1926)). Other taxa, including Limax sp., Deroceras sp., Punctum pygmaeum (Draparnaud, 1801), Truncatellina cylindrica (Férussac, 1807)), Jaminia quadridens (Müller, 1774), Abida secale secale (Draparnaud, 1801), Vitrina pellucida (Müller, 1774), and Euconulus fulvus fulvus (Müller, 1774), were relatively localized, recovered only sporadically or in very low numbers (Table 4). The specimen count, normalized to the excavated volume for each stratigraphic layer, indicates a general decrease in abundance in deeper layers. Nevertheless, the overall malacofaunal composition remains relatively stable throughout the profile. Some discontinuities are observable: the total number of remains declines steadily through Level 4, increases in Level 5, then decreases again, with a slight rise in the deepest level (Level 8). The assemblage is dominated by species typical of open environments, with the exception of Punctum, which occurs exclusively in the upper levels (1–3). Most taxa are mesophilic or xerophilic (Vallonia, Xerocrassa, Candidula), yet they co-occur with distinctly hygrophilic species (e.g. Vertigo pygmaea), a feature consistent across the profile. All taxa are surface dwellers; no endogean species were recorded. Only Limax and Deroceras exhibit limited burrowing behavior, typically confined to the upper soil horizons during periods of drought. The taxocenosis includes exclusively small-sized species. The absence of medium-sized taxa is likely due to the limited extent of the excavation. Although the identified species span a broad altitudinal range, high-elevation specialists are generally absent, with the notable exception of X. geyeri, a species currently documented only above 1500 m a.s.l. in the Western Alps (Birindelli et al., 2020; Gavetti et al., 2008). All other species are widespread and commonly found in the Ligurian Alps (Birindelli et al., 2020; Boato et al., 1985). The apparent rarity of X. geyeri may be due to underreporting, as it is often misidentified because of its morphological similarity to Candidula unifasciata.
Lipid fractions
A total of 19 sediment samples from the T1S1 profile were processed. Distribution and abundance of n-alkanes for each sample recovered are reported in Figure 7, and n-alkane ratios (i.e. OEP, ACL, Paq, and percentage of grasses over woody fossil organic matter) is shown in Figure 8.
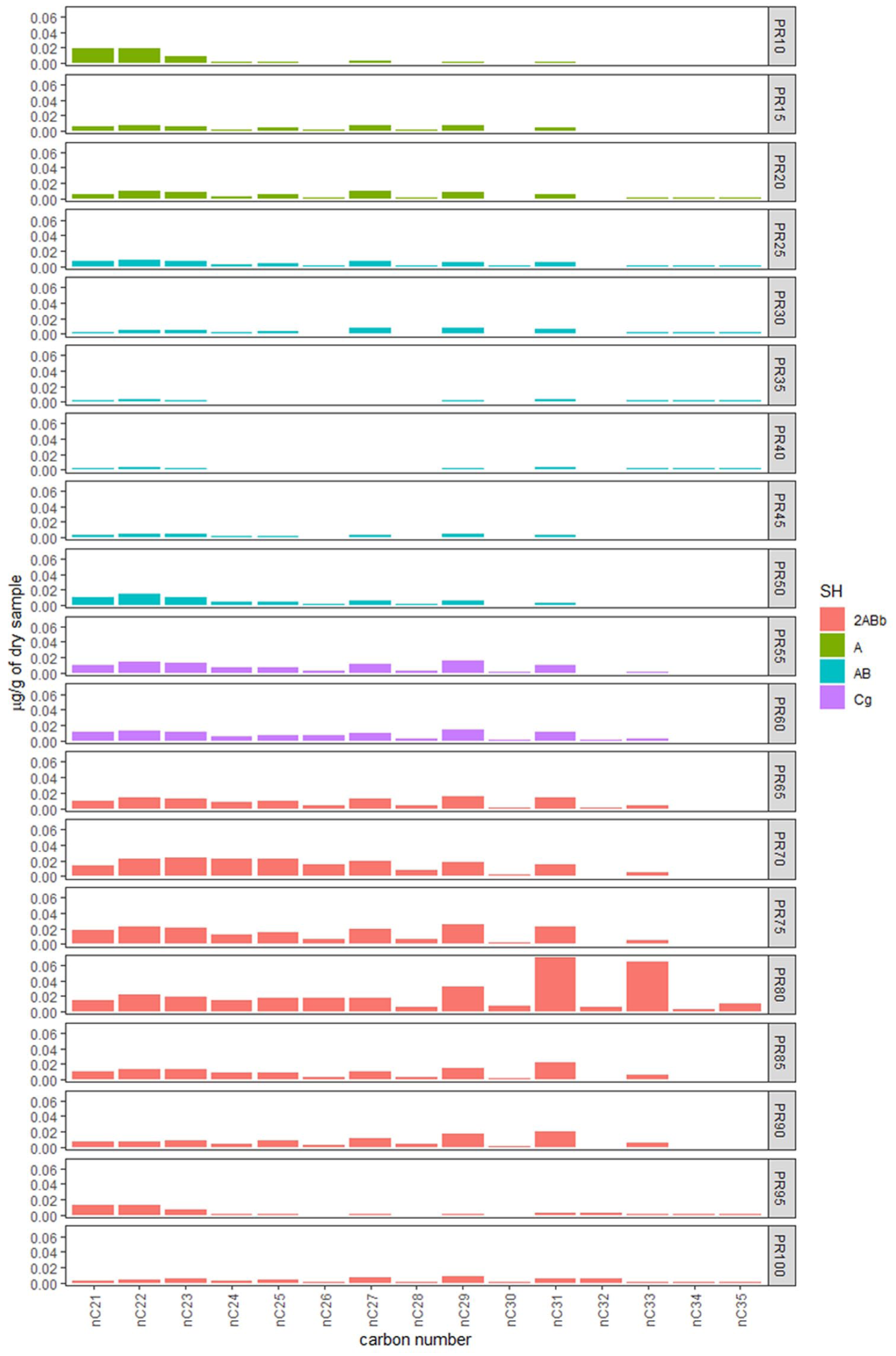
n-Alkanes concentration and distribution patterns for each sample and soil horizons (SH) indicated by color. Concentrations are expressed as μg of individual compound per gram of dry sample.
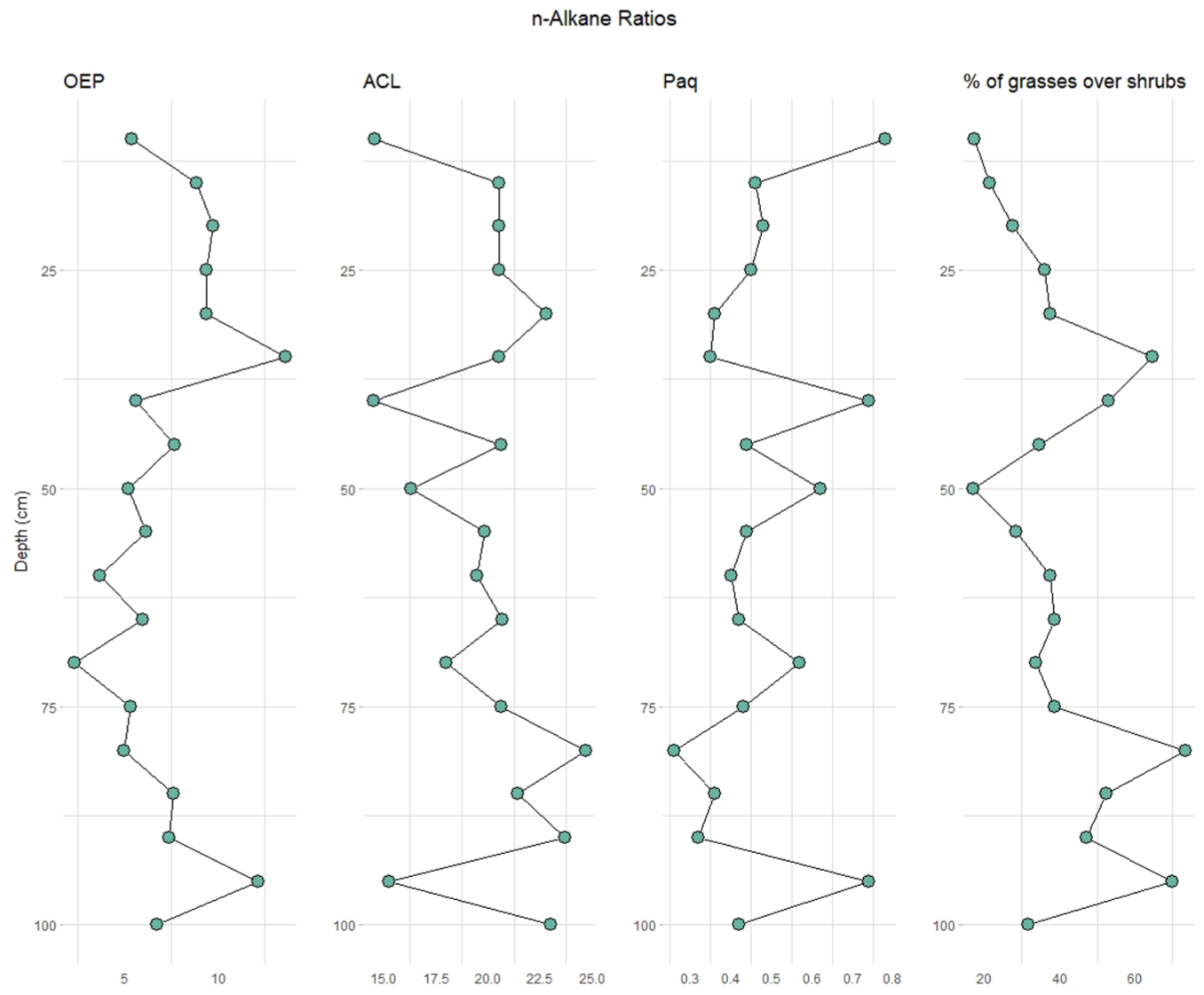
n-Alkane OEP, ACL, Paq, and % ratio (nC31 + nC33)/(nC27 + nC29 + nC31 + nC33) for each sample following depth (in cm).
All samples show mid- and long-chain n-alkanes (>nC21) with odd-over-even predominance ranging from nC21 to nC35 with a total n-alkane concentration of 1.67 µg/gds (µg/g of dry sample) and unimodal distributions. Following the depth of the sedimentary record, we distinguish three different phases. First, between 10 and 25 cm from the surface, we observe a predominance of wetland and/or meadow cover, followed by a phase of n-alkanes with Cmax at nC21, nC22, and nC29, probably indicating a transition of the previous more humid meadow to tree-shrub vegetation (30–40 cm). Between 45 and 75 cm, there is a predominance of mountain tree-shrub-type vegetation (Cmax at nC29), followed by a phase of open grasslands (80–90 cm), shown by the presence of n-alkanes with Cmax at nC31. The last sample at 100 cm deep has a predominance of tree-shrub-type vegetation with n-alkanes maximizing at nC29.
Accordingly, when the percentage contribution of grasses and herbs versus trees and shrubs is applied, we observe that the samples with the highest values are between 80 and 95 cm deep (with a mean of 60.6%). The Paq index results show values between ca. 0.7–0.2, indicating a predominance of emergent and terrestrial plants in all samples. Interestingly, the most superficial sample associated with meadow type vegetation has the highest values, thus indicating more input from submerged freshwater plants (Mead et al., 2005). OEP values range between 2.3 and 13.6, with lower values (<5) characteristic of topsoil or organic matter degradation, and higher values being typical of fresh plant material, indicating a good preservation of plant-derived organic matter. ACL ranges from 14.5 to 24.6, and correlates well with the % n-alkane ratio, indicating that the higher ACL values reflect grass abundance and warmer periods (Bush and McInerney, 2013; Tipple and Pagani, 2013).
Although the polycyclic aromatic hydrocarbons (PAHs) and steroid fractions were analyzed, no detectable peaks were observed. Instead, our sediment samples revealed the presence of long-chain trimethylsilyl ether derivatives of fatty alcohols (FA-OH), including 1-hexadecanol (C16) and 1-octadecanol (C18), at depths of 20, 30, 40, 50, 60, 70, and 100 cm. In addition, docosanol (C22), 1-tetracosanol (C24), and 1-hexacosanol (C26) were identified specifically at depths of 20 and 100 cm. Furthermore, saturated fatty acids (FA), namely hexadecanoic acid methyl ester (C16:0) and octadecanoic acid methyl ester (C18:0), were detected in samples collected from depths of 50 and 100 cm. The origins and presence of FA-OH in sediments are primarily associated with the hydrolysis of plant wax esters, especially from epicuticular waxes of terrestrial vegetation (Beeston et al., 2006). By contrast, fatty acids can typically originate from the hydrolysis of membrane lipids, as well as from microbial activity or the degradation of plant and animal organic matter (Heron et al., 1991).
Discussion
Issues of taphonomy and preservation
Among the main risks of studying dry off-site contexts are those linked to soil formation and in particular, to the multiple pedoturbation processes that may have damaged any organic remains (e.g. physical alteration of pollen grains, charcoals, seeds, etc. affecting their preservation), altered their distribution along the pedological profile (e.g. soil mixing due to bioturbation, see also Davidson et al., 1999) and modified the chemistry of the horizons. The combination of these processes may therefore compromise the representational accuracy of the recovered archeological evidence. At Pian Rosso, some of these pedoturbation processes have been documented.
The calcareous nature of the area, along with the physico-chemical variations characterizing the soil profile (i.e. pH, CEC, and water presence), appear to have significantly affected the preservation of some of the bioarchaeological remains recovered.
The poor preservation of both pollen and insect remains at Pian Rosso can largely be attributed to the calcareous nature of the soil, which significantly affects taphonomic conditions at both chemical and biological levels. Calcareous substrates are typically associated with alkaline pH values and enhanced biological activity, factors known to accelerate the degradation of organic records (Davidson et al., 1999; Dimbleby, 1957, 1984, 1985). In palynological terms, these conditions promote the action of bacteria, fungi, and soil-dwelling invertebrates, which contribute to both mechanical and biochemical breakdown of pollen grains and non-pollen palynomorphs (Elsik, 1966; Havinga, 1984). For example, pollen grains with a thick wall are more resistant; likewise, the thickness and chemical nature of the wall of the fungi spores/conidia (e.g. abundance of fungal melanins in several coprophilous fungi spores) protect them from degradation.
Similarly, the insect record is extremely limited and discontinuous, likely due to the same alkaline soil conditions, combined with the absence of permanently waterlogged or anaerobic layers (conditions known to favor the long-term preservation of insect remains (Elias, 2010; Kenward, 2009). The exceptional preservation observed in a few isolated specimens, such as the complete Megalonotus, is more likely due to recent contamination, either via bioturbation or during the sampling process. The occurrence, however, of a Myrmica sabuleti head (stage 1 of TSD) in the deepest stratigraphic layer is most likely attributable to the burrowing behavior and subterranean nesting habits characteristic of ants.
While calcareous sediments do not directly degrade biomarkers, they can create conditions that influence their preservation or degradation by affecting microbial activity, mineral interactions, and the overall diagenetic environment. The detection of long-chain n-alkanes, fatty alcohols and very few fatty acids, combined with the absence of polycyclic aromatic hydrocarbons (PAHs) and fecal esterols, suggests a selective preservation of chemically stable, non-polar lipids under early diagenetic conditions that are oxidative and alkaline in nature. This pattern is characteristic of calcareous sediments, which typically provide poor protection for labile or aromatic organic compounds (Summons et al., 2008). Fatty acids are rapidly lost via microbial respiration and hydrolysis, especially under a mild alkaline pH, which can destabilize ester bonds and promote lipid breakdown (March et al., 2014); while PAHs, especially those of low molecular weight or alkylated forms, and steroids, are vulnerable to oxidation and microbial degradation in oxic environments (Haritash and Kaushik, 2009; Misra et al., 2021). Moreover, without mineral surfaces like clays or metal oxides to adsorb and protect those, lipids are less likely to persist over time (Keil and Mayer, 2014; Zhao et al., 2022). The observations of bioturbation are attributable solely to the activity of soil fauna. Among the possible effects of this phenomenon could be the presence of intrusive insects, the transport and redistribution of both organic and inorganic soil constituents, and the redistribution of microcharcoal particles (10–50 μm size range) along the profile. Bioturbation, however, does not appear to have had significant effects (mechanical disturbance) on the soil horizonation or on the distribution of the carbonized macro-remains (macro-charcoal and seeds).
Despite these preservation issues that have affected some bioarchaeological remains, some interesting features and specific patterns could be analyzed together with documentary sources to characterize the dynamics of the soils and vegetation cover present at Pian Rosso, as well as the agro-sylvo-pastoral practices used for the management of environmental, and in particular, pastoral resources.
History of the soil archive
The pedological and microstratigraphic analysis of the three profiles excavated along the altitudinal transect has enabled the reconstruction of local soil dynamics on the southern slope of Monte Mongioie. A complex interplay between pedogenetic development, episodic colluvial inputs and agro-sylvo-pastoral practices has been identified.
Profile 1 (T1S1), located on the Pian Rosso plateau, provides the most informative sequence, serving as the key reference for understanding the local pedogenesis and its chronological framework. In this profile, the PSU1 (2ABb, 65–100 cm depth) correspond to a paleosol as indicated by elevated concentrations of organic matter (ranging 3.9 g/kg) and total nitrogen (3.64 g/kg), in contrast to the overlying layers. This paleosol also contains a greater abundance of lipid compounds, including long-chain n-alkanes and fatty alcohols, suggesting enhanced preservation and accumulation of organic residues in this layer. The combination of increased total organic content and lipid biomarkers support the interpretation of this unit as a former stable soil surface, likely characterized by vegetation cover and reduced post-depositional oxidation, which together favored biomolecular preservation. Bioarchaeological indicators such as pollen, non-pollen palynomorphs (NPPs), macrocharcoal, terrestrial mollusk remains, and charred seeds also show an increase in both abundance and concentration (Figures 4a, b, and Figures 5–7; Tables 3 and 4). Radiocarbon dating suggests that the deepest part of the paleosol (90–100 cm) may fall in the Roman period (date Poz-179505: 98 cal. BC–106 cal. AD), whilst the upper portion (from 80 cm) seems to dates to the Middle Ages (date LuS18320: 1162–1267 cal. AD). The preservation of the buried soil can plausibly be attributed to the overlying alluvial sediment (Cg – PSU2), which likely represents a rapid depositional event from the alluvial fan that sealed the underlying soil features and biotic assemblages, reducing subsequent disturbance. Radiocarbon data are compatible with a Late Middle Ages attribution for this horizon (date Poz-179504: 1299–1404 cal. AD). This event may have facilitated the preservation of specific micromorphological features characteristic of the soil surface, such as the sedimentary crust fragments and specific NPPs (e.g. Gasteromycetes, Scleroderma, and Bovista types). Following this intense alluvial event, a phase of continued but less intense sedimentation appears to have occurred, possibly related to the alluvial fan system above. This low sedimentation rate would have favored the processes of soil profile formation (humus accumulation and weathering) with A and AB horizons formation.
Radiocarbon dates performed at 10 and 25 cm deep are broadly coeval, spanning from the 17th to 20th century (date LuS18319: 1669-modern cal. AD and date LuS18318: 1693-1918 cal. AD). Both horizons appear to result from a combination of pedogenesis (humification) and slope-derived sedimentation. Evidence of colluvial processes is also observed in profile 3 (T1S3), located on the upper part of the talus slope. Radiocarbon dating from this profile yielded exclusively Roman-age results (date Poz-168001: 194 cal. BC–5 cal. AD and date Poz-168572: 172 cal. BC–8 cal. AD), with an apparent absence of Medieval and later phases. This chronological gap could reflect erosive events that removed subsequent deposits. The displaced colluvium was subsequently transported downslope, potentially contributing fine sediment to the Pian Rosso plateau, as suggested by the emplacement of sediments in the PSU2 of T1S1. By contrast, profile 2 (T1S2) is directly set on the apex of the alluvial fan and corresponds to terracing used for agricultural purposes. It demonstrates horizons that are extremely gravelly and exhibit significant textural differences. Surprisingly, radiocarbon dating performed at 45 cm yielded a Late Bronze Age/Early Iron Age date (date Poz-168000: 1203–941 cal. BC) despite the fact that the construction of this terracing (as well as the terraces present on the slopes surrounding the Pian Rosso area) was likely performed at a relatively recent date. Indeed, historical data suggests that intensive slope management practices, (possible terracing and structure construction) became viable only from the 19th century onward. Prior to this, regulations embedded in the local commons system restricted permanent modifications to the land. The demographic and socio-economic changes of the mid 19th century, including the establishment of Viozene parish and land privatization, likely marked the beginning of systematic terracing activity in the area (see the section on historical background). Thus, the Late Bronze Age/Early Iron Age dating is possibly related to soil mixing that occurred during the construction of terracing and cannot be related to the building of the terracing itself.
Pastureland and vegetation dynamics at Pian Rosso
Geo and bio-archeological markers provide a clear picture of the ecological characteristics and land use of the study area over the past centuries. Despite minor discontinuities, evidence suggests that the southern slope of Monte Mongioie has been dominated by open environments consisting of grasslands and pastures, since at least the Roman period.
Indicators of grasses and herbaceous taxa prevail throughout the soil profile T1S1, as demonstrated by the consistent trends in Cichorioideae pollen (Figure 4(a)), n-alkane biomarkers (Figure 7), and the presence of terrestrial mollusks typical of open habitats (Table 4). The general scarcity of macrocharcoal remains, except in the uppermost stratigraphic layer, which is likely linked to a specific event or land-use practice (see Section on agro-sylvo-pastoral evidence), also supports the hypothesis of limited biomass. This aligns with expectations for areas lacking dense arboreal or shrubby vegetation, which typically generate greater combustible material. Strong peaks in the presence of grassland markers were also observed along the profile at depths of 35, 80, and 90 cm. Cichorioideae and n-alkanes corresponding to grasses increase while pine decreases or even disappears completely (see at 80 cm deep), as well as other deciduous trees (which notably are practically absent in the paleosol). An increase in microcharcoal is also registered at the same time. From at least the Middle Ages, and more markedly during the post-medieval period, this interpretation is corroborated by cartographic and textual sources. These documents describe Pian Rosso as a highly valued grazing area, characterized by meadows, seasonal pastures, intense fodder production (from silver fir and possibly pine) and temporary cultivation (e.g. cereals, legumes; Quaini, 1986).
Nevertheless, some notable discontinuities suggest changes in the vegetation cover surrounding the Pian Rosso plateau. In particular, the palynological record from the most superficial stratigraphic horizon indicates an increase in arboreal taxa (notably pine) alongside a decline in herbaceous indicators such as Cichorioideae (Figure 4(a)). This rise in tree pollen, however, should not be interpreted as a reforestation event or the development of a dense woodland on the Pian Rosso plateau. Instead, it likely reflects an increase in surrounding arboreal populations. This is supported by n-alkane data, which continue to indicate a local dominance of humid meadows over woody vegetation, although this phase shows the lowest meadow signal across the entire sedimentary profile (Figure 8). The most prominent arboreal taxon is pine, an anemophilous species capable of long-distance pollen dispersal, suggesting that these stands were not locally dense but rather situated at a distance from the sampling site. This interpretation is further reinforced by iconographic sources (e.g. historical photographs, postcards, aerial imagery) dating from the late 19th century onward (Montevecchi, 2001). These sources reveal a marked expansion of forested areas (particularly along the watersheds bordering Pian Rosso and down to the Tanaro River valley) beginning only in the first half of the 20th century. This landscape transformation coincides with the progressive depopulation of the parish of Viozene and the subsequent decline in commons agro-sylvo-pastoral practices and agricultural production. Further signals of the increase in arboreal vegetation can be found in other parts of the profile, although these are more sporadic. Between 45 and 55 cm in depth a decrease of Cichorioideae coincides with an increase in pine pollen and in n-alkanes corresponding to woody and shrub-like plants (nC27 and nC29). An increase in microcharcoal is also registered at the same time. At 70 and 100 cm deep a similar pattern is registered by pollens and n-alkanes. Despite these variations the value of the bioarchaeological evidence is not consistent with the presence of a woodland in the Pian Rosso plateau and slopes. Instead, this evidence may refer to a sporadic increase in arboreal and shrub populations, consistent with the management of a tree-bearing landscape but within a wider, open environment.
Evidence for agro-sylvo-pastoral practices
The presence and patterns of some geo and bio-archeological evidence recorded along the profile T1S1, when compared with historical sources, can reveal the occurrence of specific agro-sylvo-pastoral practices, particularly those related to the use of fire, the cultivation of specific crops, and pastoral activities.
Fire use
The trends identified from the microcharcoal are interesting and contextually significant, recording higher peaks both at levels where there is an increase in tree-shrub and herbaceous populations. Although it is commonly assumed (especially in mountain regions) that an increase in microcharcoal particles coinciding with a decline in arboreal taxa and a rise in herbaceous taxa reflects phases of woodland clearance for cultivation or grazing (i.e. slash-and-burn practices), such an interpretation does not apply to the case presented here. In the Pian Rosso sequence, microcharcoal is consistently present even during periods of maximum representation of herbaceous taxa and well-established open environments (see the section on pastureland and vegetation dynamics). This could simply be a result of bioturbation or could be related to specific agro-sylvo-pastoral practices. Indeed, historical sources document fire-related practices associated with herbaceous vegetation (see the section on historical background), particularly the use of fire for fertilization and rejuvenation of the grass sward, both to improve pasture quality and to prepare land for temporary cultivation. It is in this context that the charcoal remains recovered at Pian Rosso may be interpreted. Given that the microcharcoal particles observed fall within the 10–50 μm size range (thus highly volatile and subject to long-distance transport), however, it is not possible to pinpoint the exact location of these fire events. Nevertheless, it is reasonable to assume that such practices took place in the surrounding area. Evidence for these practices is instead provided by macrocharcoal (some of them vitrified, thus heavier), which is more representative of local fires. The increase of fragments in the paleosol levels attributable to stems and rhizomes of herbaceous plants (see ‘unidentified’ taxa in pedoanthracological diagram) supports this hypothesis. At the same time, however, this raises questions about the characteristics of fire (e.g. temperatures and speeds reached by the fire) that allowed the carbonization of anatomical elements and species not generally recorded in anthracological and pedoanthracological records as they tend to be reduced to ashes. Fire temperature and rate of spread appear to be the main factors responsible for the production and preservation of this type of charcoal. In this sense, some of the techniques described above may have created a suitable situation for such high temperatures to be reached (archaeobotanical analysis suggests a possible burning temperature in excess of 400°C), in a rapid and inhomogeneous way.
Instead, the peak of macrocharcoal present in the most superficial level (Level 1 between 10 and 20 cm) dated to the 19th century, would be attributable to a different practice. In this case, the evidence can likely be attributed to an intentional deposition of charred material, produced not on site but rather transported from another location. The clear increase in macrocharcoal occurs in an environment still characterized by large open spaces, meadows and crops, therefore in a general scarcity of flammable biomass. The presence, only in this horizon, of clusters of micro-charcoal visible in the microstratigraphy (see the section on soil features and classification) support this hypothesis of an intentional addition of charcoal and ash residues (e.g. from domestic fires and/or local cheese making activities) to the soil. Furthermore, this may also indicate a possible soil fertilization practice, but using a different technique from previous systems.
Cultivation practices
Written and cartographic sources extensively document the presence of cultivated fields on the Pian Rosso plateau and the immediately adjacent slopes since the Medieval period. Some geo and bio-archeological evidence can be traced back to these practices: (1) Cichorioideae have already been reported as possible indicators of arable land (permanent and temporary) for coastal and mountainous Liguria (Cevasco et al., 2015; Guido et al., 2013; Menozzi et al., 2007); (2) The micromorphological analysis of the paleosol (T1S1–PSU1) allowed the identification of a very significant anthropogenic feature, that is buried depositional crusts. They are well preserved and characterized by rounded vesicles and well laminated (Figure 3). These crusts are formed as a result of the lateral transport of fine particles at the soil surface. They can be localized, for example in furrows or rill channels, and occur over large areas such as irrigation basins or lower slopes that are subject to sheet erosion providing surface runoff rich in suspended matter. In cultivated soils, sedimentary seal is very common, especially in micro-depressions. Fine soil particles that derive from destruction of soil aggregates accumulate in small depressions, forming a seal or a crust. Furthermore, the vesicles often concentrate below the clay layer that inhibits the escape of air. These relict features as well as indicating a rapid burial following a flood event clearly suggest tillage activities and continuous cultivation (Pagliai and Stoops, 2010). Overgrazing may also be one of the causes of sedimentary crust formation. In this case, however, this hypothesis seems less likely given the absence of micromorphological features characteristic of overgrazing (i.e. the presence of fecal spherulites, traces of microlayered trampled surfaces). Cultivation practices are additionally confirmed by (3) the presence of charred cereal grains, albeit in minimal quantities, along the T1S1 profile and specifically in correspondence with the paleosoil (Table 3). A domestic origin for these cereals (i.e. the result of kitchen waste or cereal processing) is excluded, since documentary sources describe Pian Rosso as common land. This system of managing space and resources effectively prevented the construction of permanent structures, as these were seen as possessory actions and attempts to appropriate communal areas. Such system was active between 14th and 18th centuries at least. Written documentation of 16th–18th centuries clearly reports the presence of rye, oat and legumes among the main crops cultivated in Pian Rosso, although none of these species were found along the profile. This discrepancy may suggest a possible difference in the taxa selected for cultivation between the Medieval and post-Medieval periods, although further research is needed to clarify this aspect. Moreover, the abundance of biogenic channels and excremental pedofeatures, together with secondary phosphate accumulations and dispersed microcharcoal observed in the T1S2 profile, may be interpreted as evidence of manuring practices related to the cultivation of the terraced soils (Davidson, 2002; French, 2003).
Grazing activity
The rather high percentages of predominantly coprophilous fungi suggest the use of the area for farming related activities. A shift in the main livestock raised could have occurred between the PSU1 and PSU2 (see the trend of spores of Rhytidospora and Sporormiella/Preussia). Doveri (2004, 2014) reports that these species were recorded on the dung of both domestic (cattle, sheep, donkeys, horses, goats) and wild animals (wild boars, wild rabbits, mouflons, hares, fallow deer). The lack of preservation of fecal biomarkers prevents us from making hypotheses about the type of animals grazed in this area (see section on issue of taphonomy and preservation). Nevertheless, historical sources are quite clear about the animals present in Pian Rosso, at least since the Medieval period. Until the end of the 18th century sheep and goats were the most common animals with only sporadic cattle breeding. Only in the last few decades has there been a reversal, with an increase in cattle to the detriment of sheep and goats. Supporting the interpretation of this side of Monte Mongioie (particularly its higher-altitude sector) as a grazing area is the presence of elongated phytoliths, likely derived from grasses or dung accumulation, along with its biogenic microstructure and specific chemical characteristics of the soil identified in T1S3.
Conclusion
This research highlights the high potential of dry off-sites as both archeological and environmental archives. The multi-proxy approach adopted, combined with a regressive and high-resolution analysis of the unearthed evidence, has proven effective to overcome the intrinsic issues posed by such contexts related to pedogenetic and post-depositional processes. Moreover, this study provides a valuable reference for future investigations of calcareous mountain environments.
The integrated analysis of geo- and bio-archeological evidence from the three soil profiles, supported by historical texts and maps, has revealed a complex interplay between pedogenetic development, episodic colluvial inputs, local environmental sources, and agro-sylvo-pastoral practices.
The clearest example of this dynamic is offered by Profile 1 (T1S1), on the Pian Rosso plateau. Here, a well-preserved paleosol that may date between the Roman period and the Middle Ages was identified, marked by high organic content, lipid biomarkers, and well-preserved bio-archeological assemblages. This surface was rapidly buried by a colluvial event likely in the Late Middle Ages, which sealed and protected the underlying soil features. Subsequent slower sedimentation allowed renewed pedogenetic activity, forming the AB and A horizons possibly dated between the Early Modern period and the present. Additional data from the upslope Profile 3 (T1S3) suggest sediment removal due to erosion, while Profile 2 (T1S2), located on a man-made terrace, reflects later land-use interventions likely from the 19th century. A Bronze Age radiocarbon date from this profile is interpreted as a result of soil mixing during terrace construction, rather than evidence of early occupation.
Environmental archeological data indicate the persistence of an open landscape with grassland cover from at least the Roman period onward. While some discontinuities appear, the general trend remains stable until a progressive increase in tree vegetation between the 19th and 20th centuries. This ‘renaturalisation’ is likely linked to the gradual abandonment of local agro-sylvo-pastoral practices. Such practices consisted of sheep and goat grazing, temporary cultivation, and the use of fire as a tool for turf/herb layer fertilization and rejuvenation.
These were part of a broader multiple land-use system long managed through collective rules tied to the commons regime.
Together, these findings contribute to a more nuanced understanding of long-term historical and environmental interactions in the uplands, and underline the historical role of land-use practices and related environmental effects in shaping mountain landscapes.
Supplemental Material
sj-docx-1-hol-10.1177_09596836261450787 – Supplemental material for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie
Supplemental material, sj-docx-1-hol-10.1177_09596836261450787 for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie by Valentina Pescini, Marco Bodon, Roberta Cevasco, Charlotte Diffey, Natalia Egüez, Bruna Ilde Menozzi, Diego Moreno, Sarah Parrilla and Ivano Rellini in The Holocene
Supplemental Material
sj-docx-3-hol-10.1177_09596836261450787 – Supplemental material for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie
Supplemental material, sj-docx-3-hol-10.1177_09596836261450787 for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie by Valentina Pescini, Marco Bodon, Roberta Cevasco, Charlotte Diffey, Natalia Egüez, Bruna Ilde Menozzi, Diego Moreno, Sarah Parrilla and Ivano Rellini in The Holocene
Supplemental Material
sj-docx-4-hol-10.1177_09596836261450787 – Supplemental material for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie
Supplemental material, sj-docx-4-hol-10.1177_09596836261450787 for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie by Valentina Pescini, Marco Bodon, Roberta Cevasco, Charlotte Diffey, Natalia Egüez, Bruna Ilde Menozzi, Diego Moreno, Sarah Parrilla and Ivano Rellini in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_09596836261450787 – Supplemental material for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie
Supplemental material, sj-pdf-2-hol-10.1177_09596836261450787 for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie by Valentina Pescini, Marco Bodon, Roberta Cevasco, Charlotte Diffey, Natalia Egüez, Bruna Ilde Menozzi, Diego Moreno, Sarah Parrilla and Ivano Rellini in The Holocene
Supplemental Material
sj-pdf-5-hol-10.1177_09596836261450787 – Supplemental material for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie
Supplemental material, sj-pdf-5-hol-10.1177_09596836261450787 for Long-term environmental dynamics and agro-sylvo-pastoral practices in the SW Alps (Italy): Interdisciplinary perspectives from Monte Mongioie by Valentina Pescini, Marco Bodon, Roberta Cevasco, Charlotte Diffey, Natalia Egüez, Bruna Ilde Menozzi, Diego Moreno, Sarah Parrilla and Ivano Rellini in The Holocene
Footnotes
Acknowledgements
The authors would like to express their sincere gratitude to the Municipality of Ormea for the logistical support during the fieldwork activities and for granting access to archival materials. We are also indebted to Sebastiano Carrara, Alessia Castagnino and Beatrice Palmero for their invaluable contribution in fostering local networking and for sharing historical information about the Alta Val Tanaro area. Special thanks are extended to the Municipality of Pieve di Teco for allowing consultation of the municipal archival collections. We are equally grateful to the Soprintendenza Archeologia, Belle Arti e Paesaggio for the Provinces of Alessandria, Asti, and Cuneo, particularly for authorizing the paleoenvironmental sampling activities. A very special acknowledgment goes to Fabio Negrino, whose assistance was essential throughout the entire development of the Transeant project and in fostering collaboration with the ArcheoAlps project. We also thank Anna Stagno, within the framework of the agreements between the Catalan Institute of Classical Archeology, the University of Genoa, and the Laboratory of Archeology and Environmental History (LASA). Finally, we thank Vittorio Tigrino for the support during the historical and archival research.
ORCID iDs
Ethical considerations
This article does not contain any studies with human or animal participants. There are no human participants in this article and informed consent is not required. Sampling and fieldwork have follow the Ethical Chart of the Catalan Institute of Classical Archeology (ICAC) for the prevention, detection and action against discrimination during fieldwork.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Author contributions
Valentina Pescini: Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Resources; Supervision; Validation; Visualization; Writing – original draft; Writing – review & editing.
Marco Bodon: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Roberta Cevasco: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Charlotte Diffey: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Natalia Egüez: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Bruna Ilde Menozzi: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Diego Moreno: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Sarah Parrilla: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Ivano Rellini: Data curation; Investigation; Writing – original draft; Writing – review & editing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study is part of the larger multi-scale and interdisciplinary project “Transeant - Landscape of Transhumance. Environmental Archeology research between eastern Pyrenees (Spain) and Maritime/Ligurian Alps (Italy)”, funded through the Ramón y Cajal grant program of the Agencia Estatal de Investigación (AEI) of Spain (RYC2021-034621-I). N.E. acknowledges funding from the Ramón y Cajal Programme (no. RYC2022-036901-I, MCIN/AEI/10.13039/501100011033 and FSE+). S.P. was financially supported by a Vinci grant (2020-2023) and the JCJC ANR project “BENDYS” (2019-2024, ANR-19-CE03-0010). C.D. was funded by a Marie Sklodowska Curie Action (MSCA, H2020 Marie Skłodowska-Curie Actions-MSCA-IF-2020-101024917). This research have also received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement No 853539), ANTIGONE ERC StG 2019 project (![]() ) and from the Ministero dell’Universitá e della Ricerca under the FARE 2020 programme (KORE project – CUP D33C21000240001).
) and from the Ministero dell’Universitá e della Ricerca under the FARE 2020 programme (KORE project – CUP D33C21000240001).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Data will be made available on request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.