
Abstract
Background:
Biomechanical data and contributions to ankle joint stability have been previously reported for the individual distal tibiofibular ligaments. These results have not yet been validated based on recent anatomic descriptions or using current biomechanical testing devices.
Methods:
Eight matched-pair, lower leg specimens were tested using a dynamic, biaxial testing machine. The proximal tibiofibular joint and the medial and lateral ankle ligaments were left intact. After fixation, specimens were preconditioned and then biomechanically tested following sequential cutting of the tibiofibular ligaments to assess the individual ligamentous contributions to syndesmotic stability. Matched paired specimens were randomly divided into 1 of 2 cutting sequences: (1) anterior-to-posterior: intact, anterior inferior tibiofibular ligament (AITFL), interosseous tibiofibular ligament (ITFL), deep posterior inferior tibiofibular ligament (PITFL), superficial PITFL, and complete interosseous membrane; (2) posterior-to-anterior: intact, superficial PITFL, deep PITFL, ITFL, AITFL, and complete interosseous membrane. While under a 750-N axial compressive load, the foot was rotated to 15 degrees of external rotation and 10 degrees of internal rotation for each sectioned state. Torque (Nm), rotational position (degrees), and 3-dimensional data were recorded continuously throughout testing.
Results:
Testing of the intact ankle syndesmosis under simulated physiologic conditions revealed 4.3 degrees of fibular rotation in the axial plane and 3.3 mm of fibular translation in the sagittal plane. Significant increases in fibular sagittal translation and axial rotation were observed after syndesmotic injury, particularly after sectioning of the AITFL and superficial PITFL. Sequential sectioning of the syndesmotic ligaments resulted in significant reductions in resistance to both internal and external rotation. Isolated injuries to the AITFL resulted in the most substantial reduction of resistance to external rotation (average of 24%). However, resistance to internal rotation was not significantly diminished until the majority of the syndesmotic structures had been sectioned.
Conclusion:
The ligaments of the syndesmosis provide significant contributions to rotary stability of the distal tibiofibular joint within the physiologic range of motion.
Clinical Relevance:
This study defined normal motion of the syndesmosis and the biomechanical consequences of injury. The degree of instability was increased with each additional injured structure; however, isolated injuries to the AITFL alone may lead to significant external rotary instability.
Keywords
Introduction
Injuries to the ligaments of the ankle syndesmosis are being recognized with increasing frequency. This has ranged from a reported incidence of 1% of ankle injuries in the West Point study of 1990 16 to more recent studies with an incidence rate of 7% to 25% of all ankle injuries.14,19,35 In spite of improved diagnostic methods, diagnosis of ankle syndesmotic injuries remains difficult by physical examination and radiographic methods.3,6,9,24,30,31 Furthermore, controversy continues regarding the ideal operative treatment method of syndesmosis injury, with techniques including screws, suture-buttons, and various combinations of the two. * A comprehensive understanding of the biomechanical properties of the normal and injured syndesmosis could provide an objective foundation for better diagnosis, treatment, and rehabilitation.
The stability of the weight-bearing ankle joint and syndesmosis is attributable in large part to the bony anatomy of the distal tibia and fibula. The ligamentous stability of the syndesmosis relies on the anterior inferior tibiofibular ligament (AITFL), interosseous tibiofibular ligament (ITFL), and the superficial and deep fibers of the posterior inferior tibiofibular ligament (PITFL). Recently, the anatomy of the syndesmosis has been thoroughly described both qualitatively and quantitatively with respect to surrounding bony anatomy.1,10,36,37 Previous biomechanical studies have provided information about individual ligament properties and their contributions to joint stability including resistance to syndesmotic diastasis (via lateral traction) and external rotation, as well as the effects of injury on tibiotalar joint contact pressures and mechanics.8,22,25,27-29,39 The research of Ogilvie-Harris et al is frequently referenced, in which diastasis of the distal tibiofibular joint was assessed following sectioning of individual ligaments. However, it should be noted that these specimens were disarticulated at the tibiotalar joint, and diastasis was elicited by an isolated lateral traction force until 2 mm of diastasis was achieved. 25 Although valuable and clinically applicable, these results have not been sufficiently investigated and validated based on more detailed anatomic descriptions or through the use of current biomechanical testing devices and more physiological protocols. 36
Therefore, the purpose of the current investigation was to evaluate the contributions of the individual syndesmotic ligaments to rotational stability of the syndesmosis using a modern and reproducible testing method. It was hypothesized that the stabilizing roles of the 3 syndesmotic ligaments, including the AITFL, ITFL, and PITFL, could be quantified within the context of a clinically relevant injury mechanism and biomechanical testing protocol. The study was conducted with the intention that the results may help improve the overall clinical treatment of these injuries, from initial detection and diagnosis through operative treatment and postoperative management.
Materials and Methods
Specimen Preparation
Eight matched-pair, lower leg specimens (median age 52 years, range 38-64 years; 8 male), obtained from the tibial plateau through toe tip, with no history of previous foot and ankle injury, surgery, or other confounding pathology were utilized for testing. Specimens were dissected to identify and isolate the ligaments of the syndesmosis, including the AITFL, ITFL, and PITFL, including its superficial and deep fibers. Given that the purpose of this investigation was to quantify the contributions of the static stabilizers of the syndesmosis, all overlying soft tissue was removed. All other ligamentous restraints were left intact, including the structures of the proximal tibiofibular joint, and the medial and lateral ankle ligaments.
Specimens were then prepared for biomechanical testing. To facilitate mounting to the dynamic, biaxial testing machine (ElectroPuls E10000; Instron Systems, Norwood, MA), 2 anterior-posterior–directed bicortical pins were used to secure the proximal tibia to a custom-loading fixture mounted to the actuator of the testing machine. The distal pin was secured in the proximal portion of the tibial diaphysis to ensure rigid fixation throughout testing. Distally, the foot was rigidly mounted to a custom steel fixture via three 6-mm pins. Two were placed through the side of the fixture into the lateral calcaneus, and a single pin was inserted posteriorly. To isolate measurements due to the motion at the tibiotalar joint, the subtalar joint was rigidly fixed using 2 superior-inferior–directed screws placed through the talar neck and posterior aspect of the talar dome into the calcaneus. Finally, a steel strap was placed over the dorsal aspect of the foot and secured to the base of the testing fixture. This fixture was then clamped to the testing machine such that the specimens were oriented in neutral plantar and dorsiflexion and neutral internal/external rotation with respect to the vertical tibia (Figure 1).
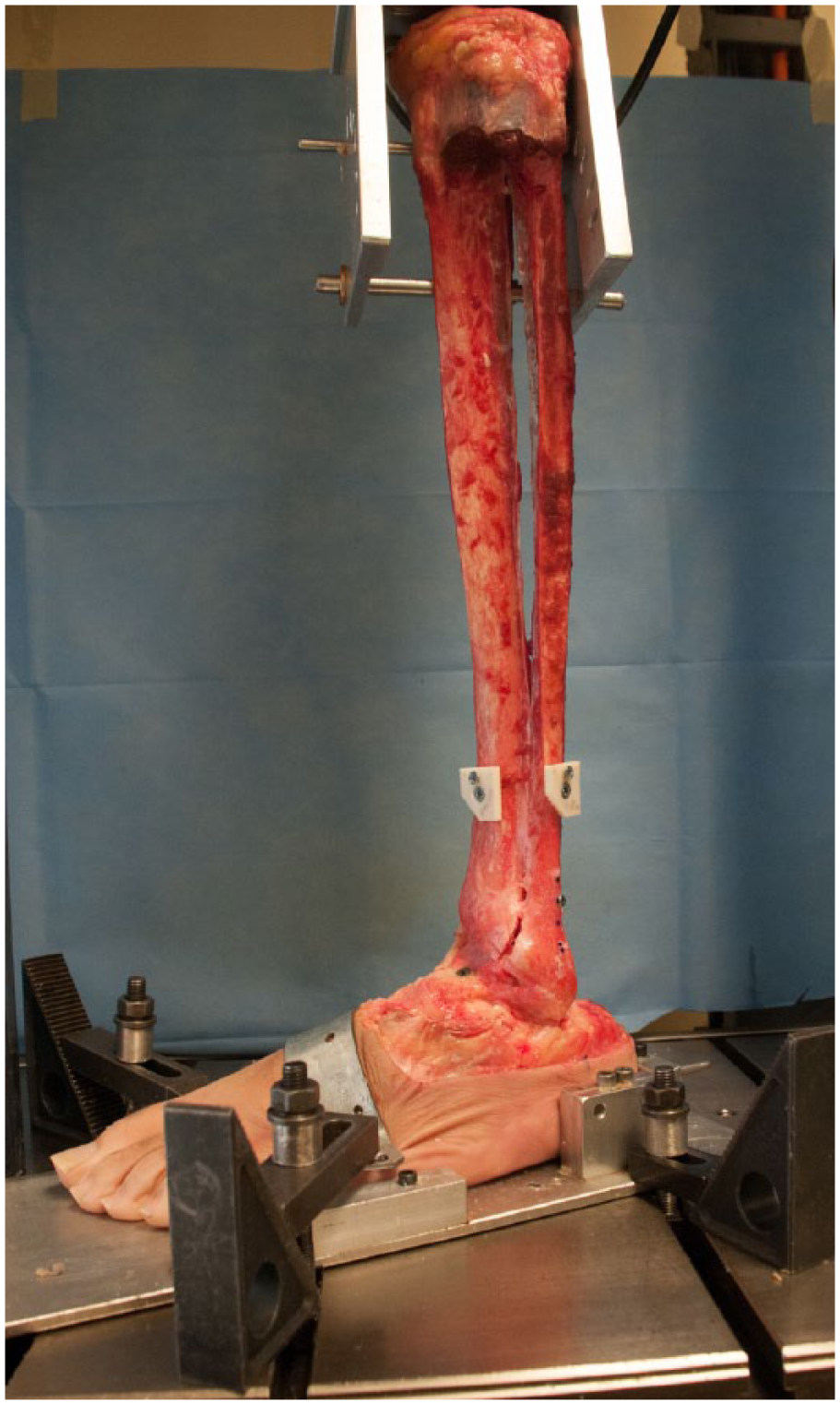
Left lower leg specimen secured in the dynamic, biaxial testing machine. All syndesmotic structures have been divided.
Anatomic Coordinate System
Prior to biomechanical testing, an anatomic coordinate system for the syndesmosis was established based on the International Society of Biomechanics (ISB) recommendations and previous anatomic literature. 38 The medial-lateral axis was established by the line connecting the medial and lateral malleolus. The superior-inferior axis was coincident with the vertical axis of the tibia, which was established by 3 series of 4 circumferentially collected points about the tibial diaphysis. The anterior-posterior axis was then defined as the calculated line mutually perpendicular to the superior-inferior and medial-lateral axes. All measurements, including tibial and fibular translations and rotations, were reported with respect to this anatomic coordinate frame. In addition, at the time this coordinate frame was established, coordinate frame blocks were attached to the distal tibia and fibula to allow for 3-dimensional (3-D) positional data collection during biomechanical testing.
Biomechanical Testing
Specimens were tested biomechanically following sequential cutting of the ligaments of the syndesmosis to assess the individual ligamentous contributions to syndesmotic stability. Matched paired specimens were split and assigned to 1 of 2 cutting sequences. The first pair was split randomly while subsequent pairs were assigned alternately to ensure an even number of left and right specimens in each group. The first cutting sequence was an anterior-posterior–directed sequential sectioning of the syndesmosis: intact, AITFL, ITFL, deep PITFL, superficial PITFL, and the complete interosseous membrane (Figure 2). The second cutting order reversed the order of the first, sequentially sectioning the syndesmosis from posterior-to-anterior: intact, superficial PITFL, deep PITFL, ITFL, AITFL, and complete interosseous membrane. Two sectioning orders were chosen to minimize directional bias and aid in determining which ligament contributed the most to syndesmotic stability. The cutting states are defined in Table 1.
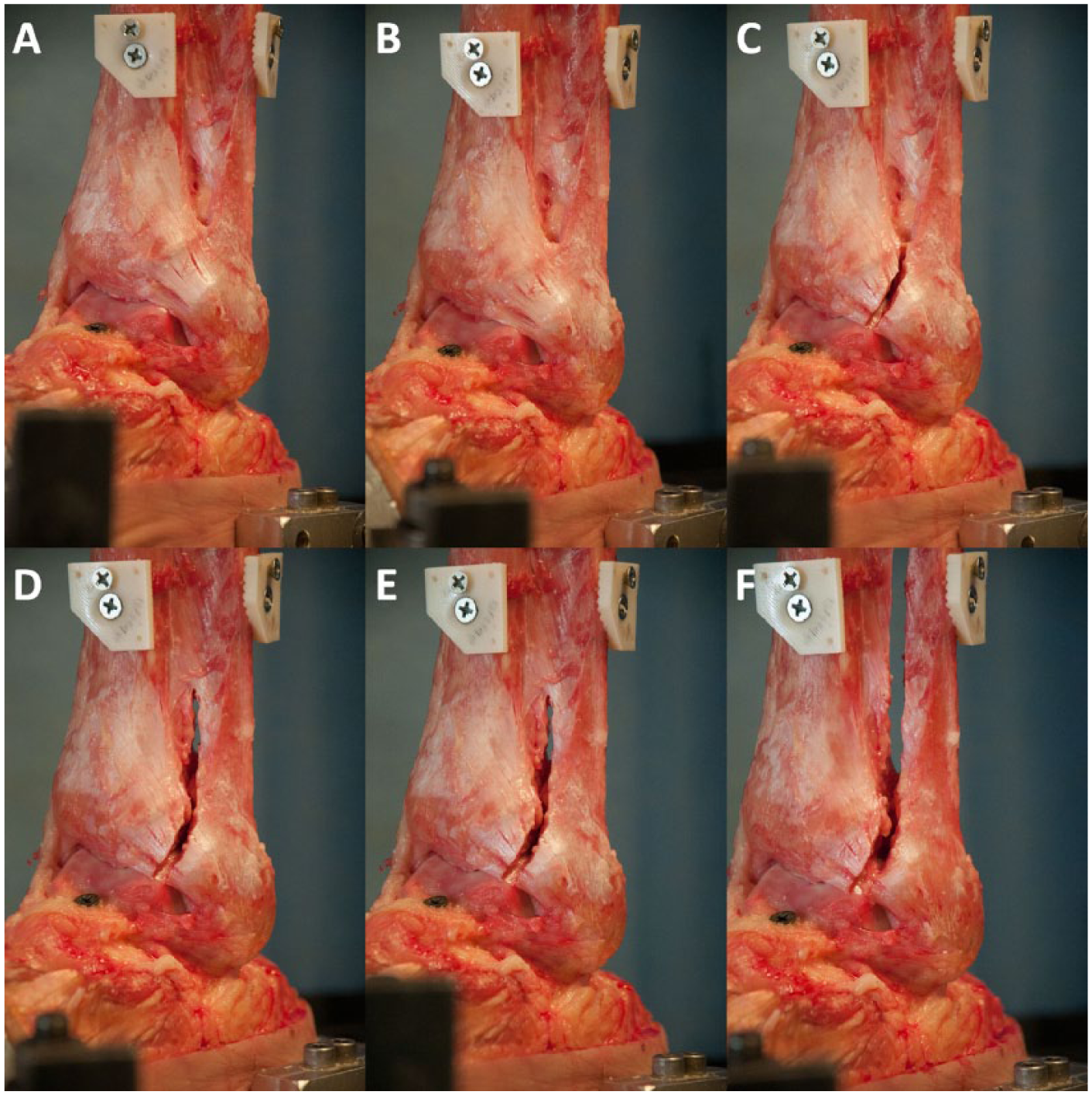
Photographs of a left lower leg specimen following sequential anterior-to-posterior sectioning. Neutral rotation: (A) intact syndesmosis. 15° external rotation: (B) intact syndesmosis, (C) AITFL sectioned (state 1AP), (D) AITFL + ITFL sectioned (state 2AP), (E) AITFL, ITFL, dPITFL and sPITFL sectioned (state 4AP), (F) IOM sectioned (all cut).
Description and Abbreviation Key for Cutting States. a
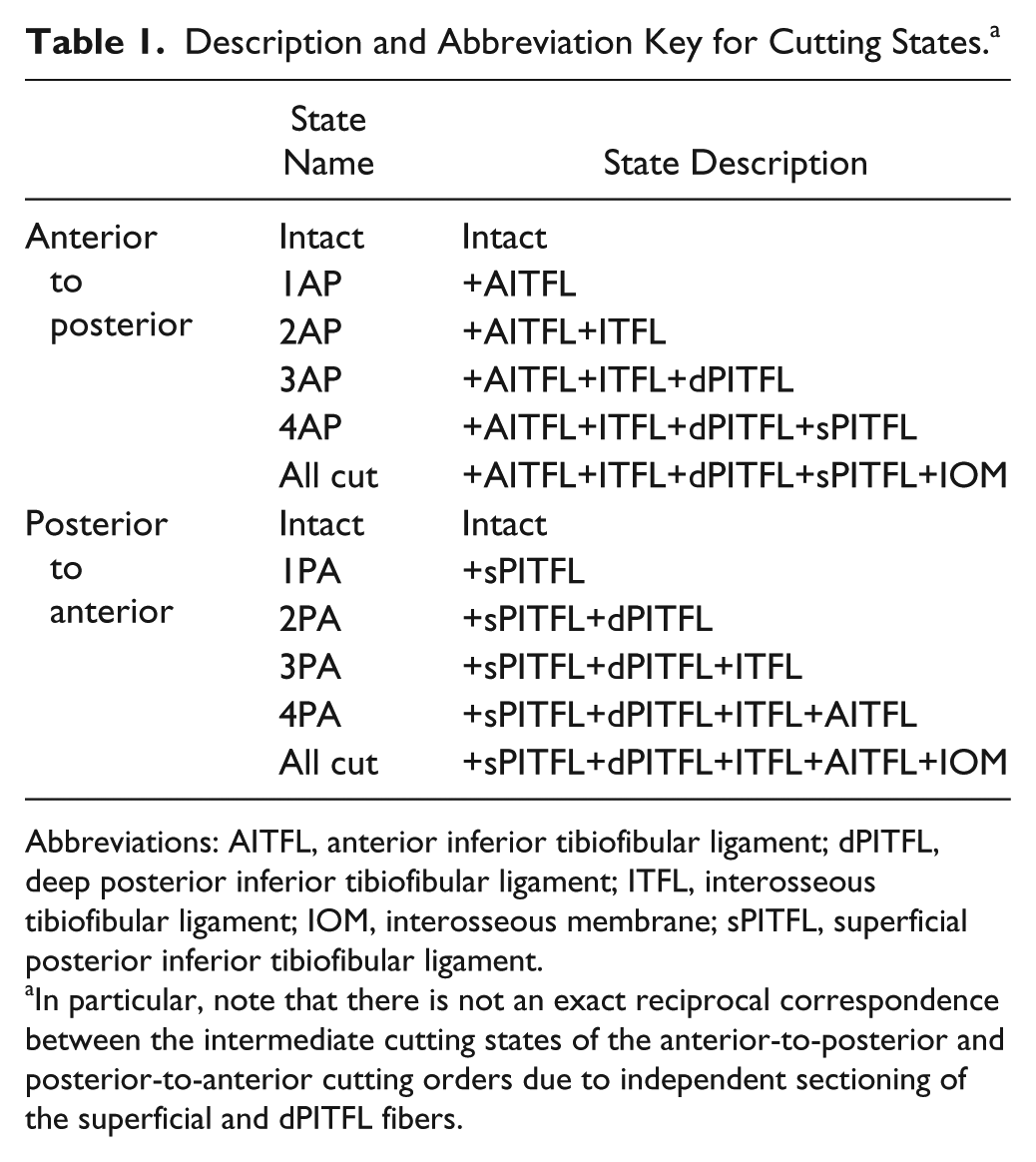
Abbreviations: AITFL, anterior inferior tibiofibular ligament; dPITFL, deep posterior inferior tibiofibular ligament; ITFL, interosseous tibiofibular ligament; IOM, interosseous membrane; sPITFL, superficial posterior inferior tibiofibular ligament.
In particular, note that there is not an exact reciprocal correspondence between the intermediate cutting states of the anterior-to-posterior and posterior-to-anterior cutting orders due to independent sectioning of the superficial and dPITFL fibers.
After fixation in the testing apparatus, specimens were preconditioned for 10 cycles to a maximum applied torque of ±7.5 Nm while a 750-N axial compressive load was applied and maintained. Specimens were then tested biomechanically under several conditions to assess the rotational stability of the syndesmosis at each sectioned state (Instron Systems, Norwood). With each testing condition, the relative positions and rotations of the tibia and fibula were recorded via the distally transfixed coordinate frame blocks and a 3-D coordinate measuring device (7315 Romer Absolute Arm, Hexagon Metrology; manufacturer-reported point repeatability of 0.025 mm). These positions were later used to calculate changes in tibiofibular rotation and translation. The first condition was a neutrally positioned orientation under a 0-N axial load intended to represent a non-weightbearing state. Next, a 750-N axial force was applied to simulate a weightbearing load. While maintaining the weightbearing axial load, the foot was externally rotated to 15 degrees and internally rotated to 10 degrees. The order of internal and external rotation was reversed for left and right specimens. Torque (Nm) and rotational position (degrees) were recorded continuously throughout testing. Additionally, the relative translation and rotation of the fibula were recorded at the extreme of each testing condition via the coordinate frame blocks and 3-D coordinate measuring device. The above-described protocol was then repeated for each sectioned state. Note that all 3-D movement data presented herein are of the fibula with respect to the tibia relative to the neutrally oriented, axially loaded intact syndesmosis state.
Statistical Analysis
To address the primary purpose of evaluating the contributions of the individual syndesmotic ligaments to rotational stability, comparisons were made between cutting states for each measurement type (torque, translation, or rotation), experimental condition (ER or IR), and cutting order (AP or PA). The intact condition was compared to each of the subsequent cut states, and 4 additional comparisons were made comparing between sequential cutting states. Equal variance among groups was not assumed, thus paired t tests were used and the Holm method was used to control the family-wise type I error rate among the 9 comparisons. The statistical computing software R was used for all analysis and figures (R Development Core Team, Vienna, Austria) and Holm-adjusted P values smaller than .05 were deemed significant.
Results
Torque
Intact Specimens
In intact specimens assigned to the anterior-to-posterior cutting order, the mean ± SD torque was 10.5 ± 6.2 Nm in external rotation and 14.4 ± 4.2 Nm in internal rotation. Similarly, intact specimens assigned to the posterior-to-anterior cutting order had a mean torque of 11.6 ± 5.3 Nm and 13.2 ± 3.8 Nm for external and internal rotation, respectively. The intact torque values were not statistically different between treatment groups.
Effect of Loading and External Rotation on Sectioned Specimens
While sectioning of the ligaments in the anterior-to-posterior direction, cutting the AITFL (state 1AP) resulted in the largest percentage reduction in resistance to external rotation (24%, 95% CI 14.1, 34.0). Similarly, when the AITFL was sectioned in the posterior-to-anterior cutting direction (state 4PA), a 24.3% (95% CI 19.2, 29.4) reduction in resistance to external rotation was noted from the previous state (Table 2, Figure 3). Resistance to external rotation significantly decreased at each additional cutting state regardless of cutting order (P < 0.05) with the exception of cutting state 4AP in the anterior-to-posterior direction specimens (Table 2, Figure 3).
External and Internal Rotation Torques Relative to the Intact State for Sequential Anterior-to-Posterior and Posterior-to-Anterior Ligament Sectioning Directions.
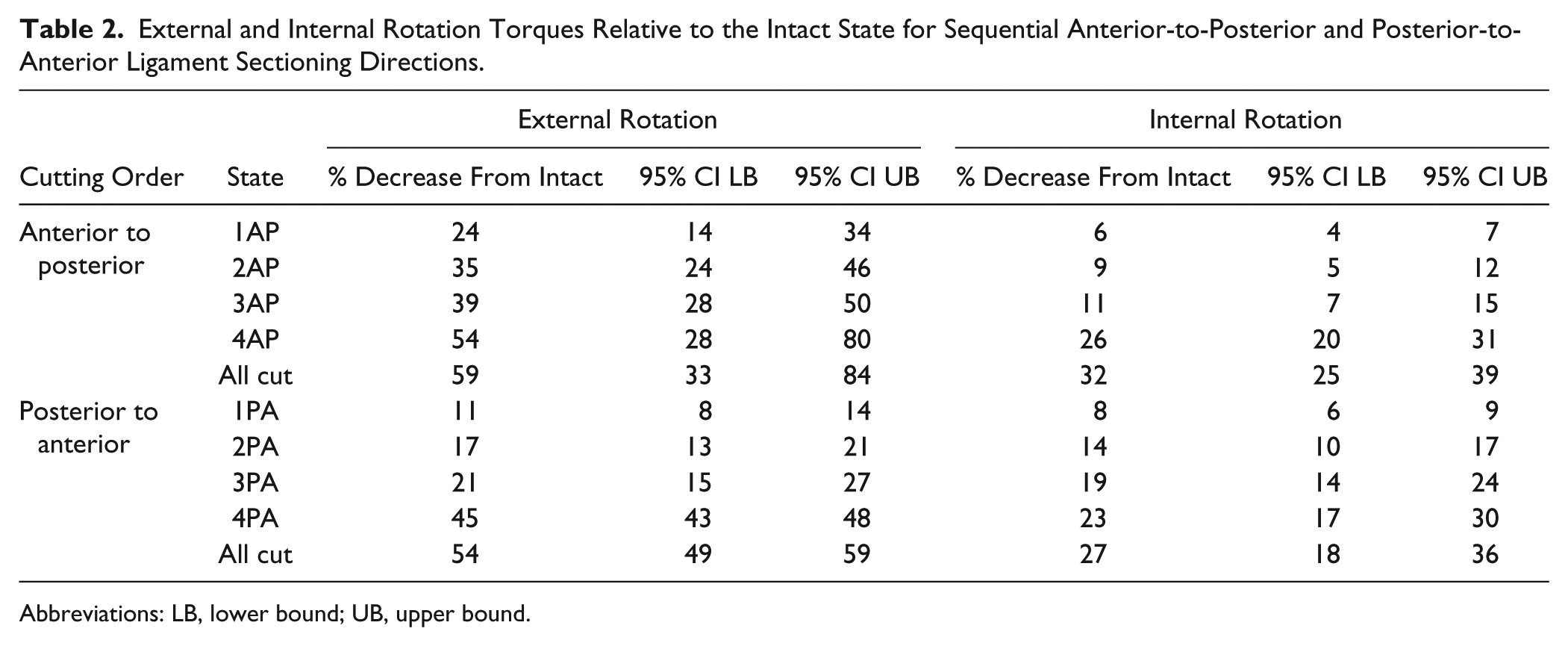
Abbreviations: LB, lower bound; UB, upper bound.
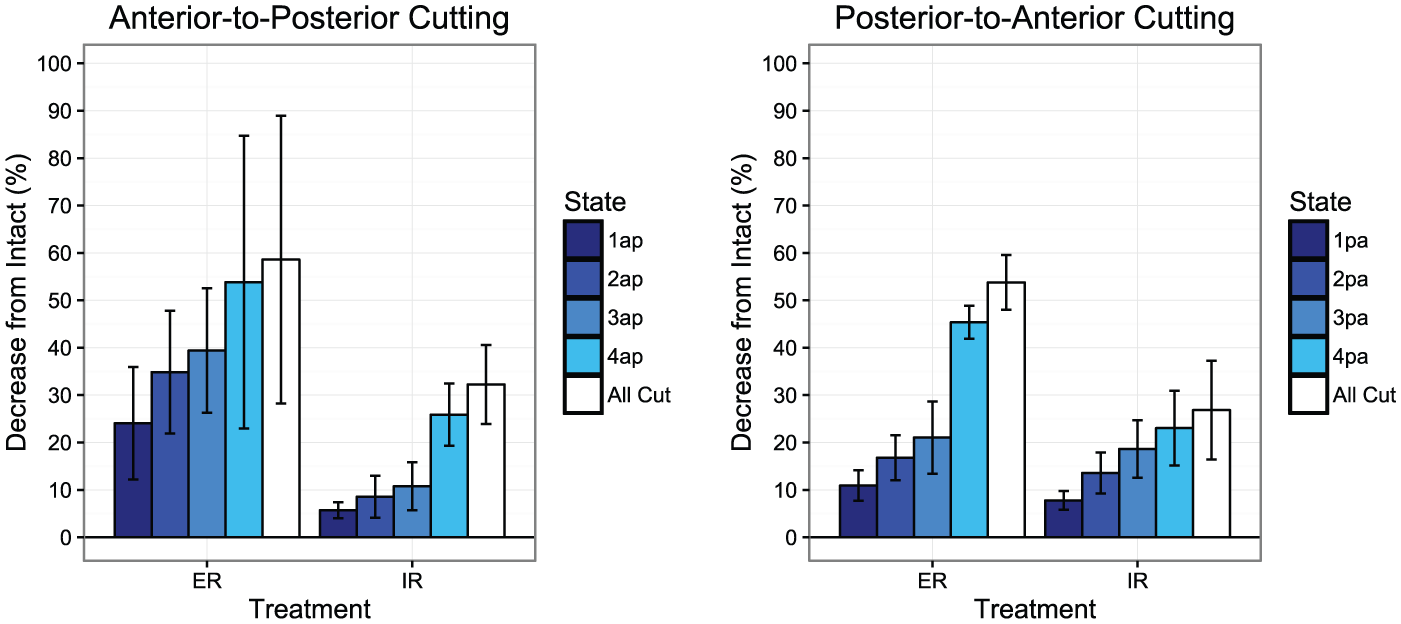
Graphical representations of reductions in torque in subsequent cutting states (defined in Table 1) for both anterior-to-posterior and posterior-to-anterior cutting orders. Error bars represent +/- SD.
Effect of Loading and Internal Rotation on Sectioned Specimens
Sectioning of the superficial PITFL resulted in the largest incremental percentage torque reduction in internal rotation, regardless of cutting order. When cutting structures from anterior-to-posterior, cutting the superficial PITFL resulted in an average incremental decrease of 15.1% (95% CI 10.8, 19.5) in torque. In the posterior-to-anterior cutting specimens, torque decreased an average of 7.8% (95% CI 6.1, 9.4) with internal rotation. Resistance to internal rotation significantly decreased at each additional cutting state regardless of cutting order (P < 0.05, Table 2, Figure 3).
Three-Dimensional Movement of the Fibula
Intact Specimens
For intact ankles, 3-D movement data of the fibula relative to the tibia revealed an average of 4.3 degrees of total rotation in the axial plane, and 3.3 mm of total translation in the sagittal plane (Figures 4 and 5, Table 3). When pooling intact data from both groups, the fibula was found to externally rotate 2.8 ± 1.2 degrees and translate 2.6 ± 1.5 mm posteriorly when the foot was loaded and externally rotated. Likewise, when loading and internally rotating the foot, the intact fibula internally rotated 1.5 ± 0.7 degrees and translated 0.7 ± 1.7 mm anteriorly (Figures 4 and 5, Table 3).
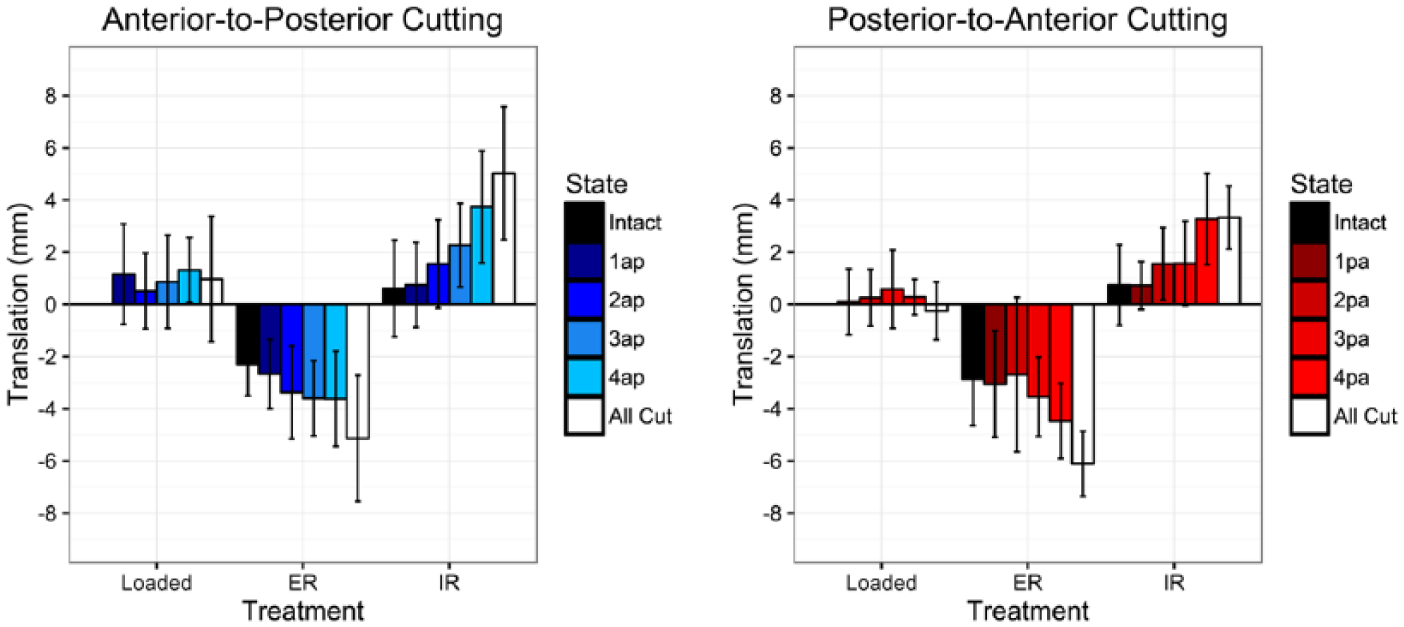
Sagittal motion of the fibula with loading, external, and internal rotation of the foot at each cutting state for each cutting order. A positive value refers to anterior translation; a negative value refers to posterior translation. Values are reported in millimeters relative to loaded neutral intact controls. Error bars represent +/- SD.
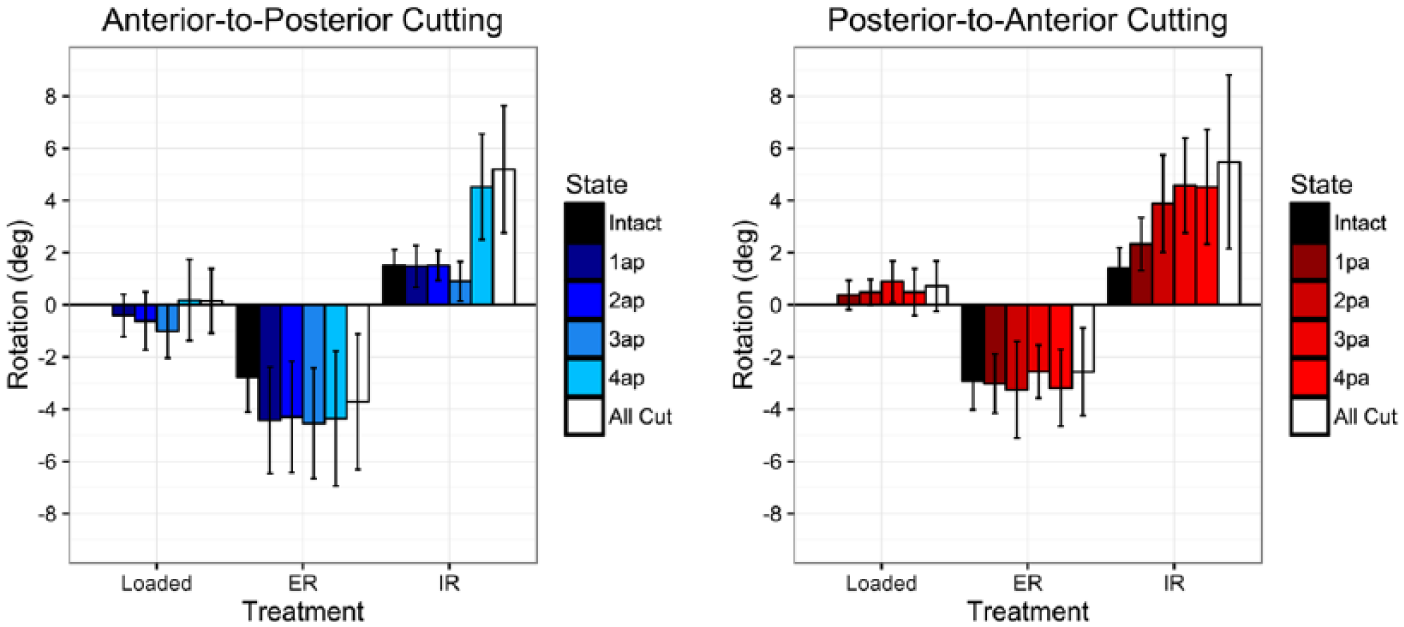
Axial rotation of the fibula with loading, external, and internal rotation of the foot at each cutting state for each cutting order. A positive value refers to internal rotation; a negative value refers to external rotation. Values are reported in degrees relative to loaded neutral intact controls. Error bars represent +/- SD.
Fibular Sagittal Translation and Axial Rotation With Loading, External Rotation, and Internal Rotation of the Foot at Each Cutting State for Each Cutting Order. a
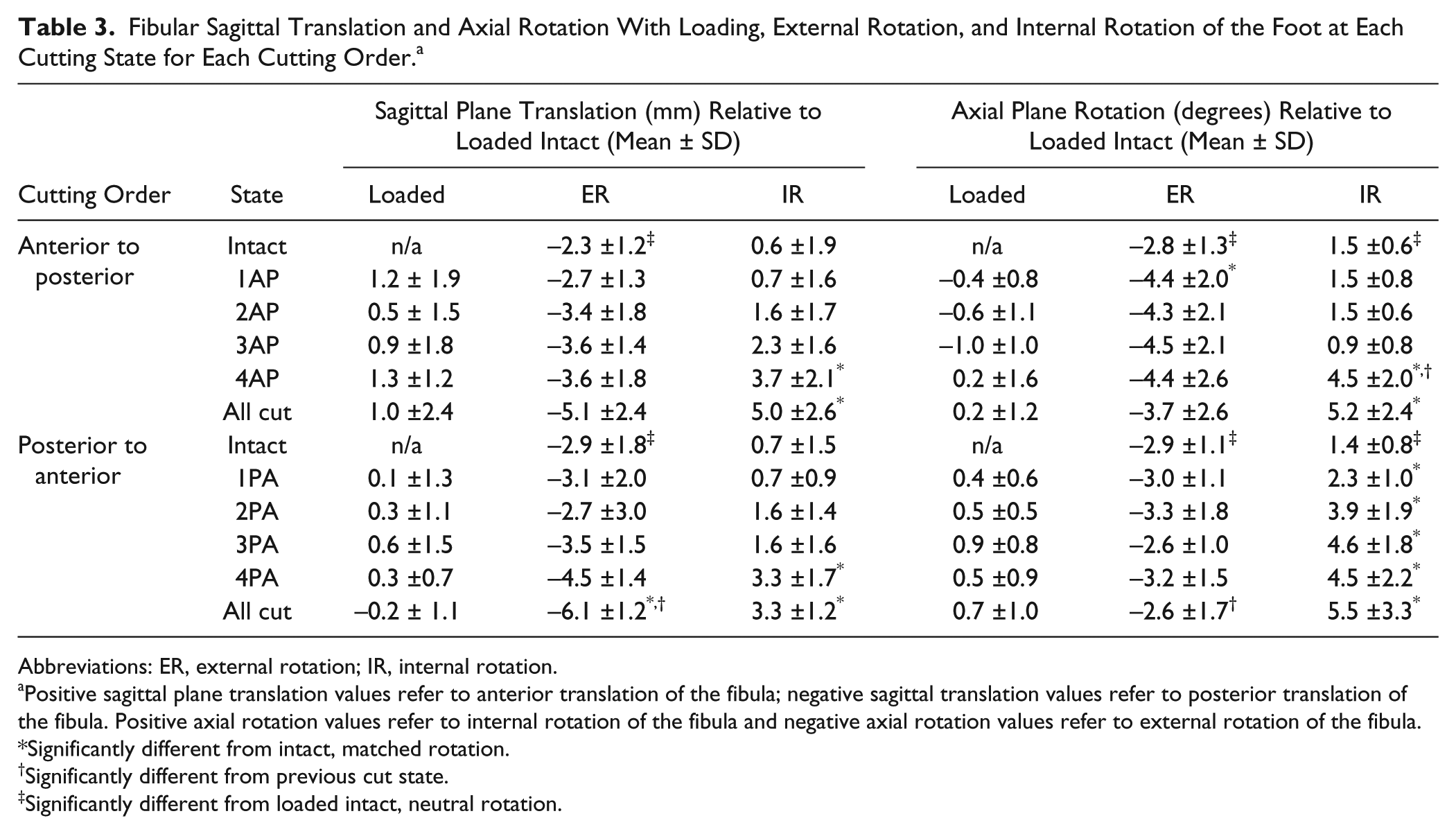
Abbreviations: ER, external rotation; IR, internal rotation.
Positive sagittal plane translation values refer to anterior translation of the fibula; negative sagittal translation values refer to posterior translation of the fibula. Positive axial rotation values refer to internal rotation of the fibula and negative axial rotation values refer to external rotation of the fibula.
Significantly different from intact, matched rotation.
Significantly different from previous cut state.
Significantly different from loaded intact, neutral rotation.
Loaded Specimens
Compared to the unloaded state, no significant changes were noted in fibular translation or rotation in any plane when specimens were loaded but not rotated regardless of cutting state or cutting order.
Effect of Loading and External Rotation on Sectioned Specimens
In both the anterior-to-posterior and posterior-to-anterior cutting orders, there was a trend toward increased posterior fibular translation with each sectioned structure; however, this was not significant until all of the structures were sectioned in the posterior-to-anterior cutting order (P < .001, Table 3, Figure 4). Rotationally, a 1.7-degree increase in external rotation was observed following sectioning of the AITFL in the anterior-to-posterior cutting order (state 1AP, P < .05).
Effect of Loading and Internal Rotation on Sectioned Specimens
In both cutting orders, there was a stepwise increase in anterior fibular translation at each cutting state (Table 3, Figure 4.). In the anterior-to-posterior cutting order, a significant increase in anterior fibular translation was only noted following sectioning of the superficial PITFL (state 4AP, P < .05). In the posterior-to-anterior cutting order, a significant 2.5-mm increase in anterior fibular translation compared to the intact state was observed following sectioning of the AITFL (state 4PA, P < .01).
In anterior-to-posterior sectioning, a significant increase was noted in fibular internal rotation following sectioning of the superficial PITFL when compared to intact specimens (state 4AP, P < .05). This was an incremental increase of 3.6 degrees compared to the prior cutting state (P < .01). In the posterior-to-anterior cutting order specimens, there was a stepwise increase in internal fibular rotation following each sectioned state. Each cutting state displayed a significant increase in internal rotation when compared to intact specimens (P < .05); however, the cutting states were not significantly different from previous cut states (Table 3, Figure 5).
Additional Rotational and Translation Data
The fibula translated in the medial-lateral direction with large variability independent of loading, foot rotation, cutting state, or cutting order. Negligible fibular movement (<1 mm) was observed in superior-inferior translation regardless of loading or cutting state. Rotation in the sagittal and coronal planes were also negligible secondary to bony constraints of the distal and proximal tibiofibular articulations as well as the ligamentous and capsular constraints of the proximal tibiofibular joint (see appendices A and B).
Discussion
The data presented herein support prior literature in that the AITFL provides clincally significant stability to the syndesmosis, specifically providing resistance to posterior fibular translation and external rotation of the lateral malleolus when an external rotational force is applied. The PITFL also provides a significant, but smaller, contribution to stability, primarily providing resistance to internal rotation. The results of the present study provide detailed quantitative and statistical analysis of the contributions of each of the syndesmosis ligaments using a reproducible biomechanical model and measurement technique.
There has been increasing interest in the study of syndesmotic injury and repair in recent years, as there has yet to be consensus data regarding the diagnosis and management of these debilitating and persistent injuries.12,20,34 Many reference the Ogilvie-Harris et al study, in which medial to lateral diastasis of the distal tibiofibular joint was assessed with 8 transtibial cadaveric specimens through sequential sectioning of each element of the syndesmosis. 25 It should be noted that those specimens were disarticulated at the tibiotalar joint prior to testing using lateral traction rather than a potentially more physiologic rotational model. The results of the study determined that the AITFL contributed the most to stability of the joint (35%), followed closely by the deep PITFL (33%). Although this was a landmark study, it lacked anatomic and biomechanical analysis of the role of each individual element of the syndesmosis before and after sectioning. Additionally, specimens were not tested in a manner in which they would be stressed naturally.
The syndesmosis is most commonly injured following an external rotation and hyper-dorsiflexion moment about the ankle, with the AITFL injured first. 5 Our results demonstrate that the syndesmosis, and particularly the AITFL, is a primary stabilizer to external rotation of the foot. Our torque data suggest the AITFL is responsible for a 24% reduction in resistance to external rotation of the foot when injured first, with each additional injured syndesmotic element leading to further instability. The superficial PITFL also appears to be an important structure in restricting internal rotation of the fibula, and sectioning of this ligament yielded a significant 15.1% and 7.8% reduction in torque in the anterior-to-posterior and posterior-to-anterior cutting order specimens, respectively.
Comparisons of loaded to unloaded states revealed very little fibular motion in any direction, including rotation; however, there was normal motion of the fibula when the foot was internally and externally rotated. The data reported here suggest approximately 3.3 mm of total anterior-to-posterior translation (sagittal motion) of the fibula with respect to the tibia in normal physiologic motion. We observed 2.6 mm of posterior fibular translation with external rotation and 0.7 mm anterior translation with internal rotation. These results are consistent with previously reported data.2,18 With each injured state, this motion increased with both internal and external rotation of the foot to a total of 9.4- and 10.2-mm of translation in the sagittal plane for the posterior-to-anterior and anterior-to-posterior groups, respectively. This is in contrast to motion in the coronal plane (medial-lateral), which was present in all cut states but not in the intact specimen. Unfortunately, these data were highly variable and not statistically significant and could not be compared to previous research.
There was no significant increase in internal rotation of the fibula with internal foot rotation until complete sectioning of the superficial PITFL, at which point internal rotation increased from 1.5 (intact) to 4.5 degrees. This dramatic increase occurred in the anterior-to-posterior cutting specimens, but not in the posterior-to-anterior cutting specimens. Rather, there were smaller stepwise increases in internal rotation at every sectioning state in the posterior-to-anterior cutting order specimens.
Fibular motion in the intact and injured state has not been sufficiently characterized with modern biomechanical techniques. The data presented herein suggests that there is physiologic translation and rotation of the fibula that significantly increases with injury to the syndesmosis, particularly the AITFL and superficial PITFL. More research is necessary to understand how these data correspond to plain and stress radiography as well as different fixation techniques. Fibular translation in the sagittal plane may be readily visualized on lateral radiographs as demonstrated by Xenos et al. 39 Compared to mortise stress radiographs, lateral projection stress radiograph demonstrated both greater interobserver reliability and higher correlations with measurements of diastasis. 39 Yet, the use of plain radiographs may be less important as MRI prevalence and its quality continues to improve. 7
In a previous cadaveric model, Hunt et al reported a significant increase in posterior translation of the fibula occurred only after interosseous ligament release, yet no syndesmotic widening was observed. 18 They also found that mean tibiotalar contact pressure increased significantly after interosseous ligament release, and the center of pressure shifted posterolaterally. This contact pressure was increased with loading and external rotation of the foot. The data in the present study demonstrated that the most significant translation and rotation of the fibula occurred with loading and external rotation. This increased fibular movement demonstrated in the present investigation could be a contributing cause of the increased tibiotalar contact pressures noted in the study by Hunt et al. 18 In treating syndesmotic injuries, it is unclear which fixation technique would successfully decrease these contact pressures yet allow for physiologic fibular motion. Regardless of fixation technique, this study suggests that repair of the AITFL may be necessary to augment centrally placed transosseous fixation to help control for rotational instability that may persist after syndesmotic fixation. More information is needed to determine the clinical ramifications of screw versus suture-button fixation in this regard.
The authors acknowledge limitations of the present study. Foremost, this study utilized a cadaveric biomechanical model that may not be completely reflective of the in vivo kinematics or mechanism of injury. In order to ensure correct identification and accurate creation of individual ligament injuries, all overlying soft tissue were removed prior to testing. The authors acknowledge that this tissue may provide additional static and dynamic stability to the syndesmosis in vivo, and the data and conclusions must be interpreted accordingly. Furthermore, this biomechanical model implemented a simplified loading protocol using uniform axial loads and torques that do not completely replicate the complex in vivo kinematics of the syndesmosis during both daily and athletic activities. Additionally, the design of this study did not allow for direct statistical comparison between anterior-to-posterior and posterior-to-anterior groups. Therefore, this study cannot provide definitive information that would determine, for instance, whether the AITFL or PITFL provides the most stability to the syndesmosis. The authors believe that the comprehensive data set provides the opportunity for such relative comparisons; however, further study is needed in order to answer this question formally. Finally, it is possible that relaxation of tissues and ligamentous restraints could potentially contribute to decreased torque values and increased fibular motion observed with subsequent sectioned states. Regardless of the inherent limitations of the given cadaveric model, the authors believe that the trends established by the data provide valuable and clinically relevant information for both the diagnosis and treatment of syndesmosis injuries.
In conclusion, the ligaments of the syndesmosis provide significant contributions to the rotary stability of the distal tibiofibular joint within the physiologic range of motion. Among these structures, the AITFL is very important in providing resistance to external rotation and posterior translation of the fibula whereas the PITFL is an important structure involved in controlling internal rotation.
Footnotes
Acknowledgements
The authors thank Angelica Wedell for medical photography and Mary T. Goldsmith, MS, for assistance with data processing.
Authors’ Note
The investigation was performed at the Department of BioMedical Engineering, Steadman Philippon Research Institute, Vail, Colorado.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Thomas O. Clanton, MD, is a consultant and receives royalties from Arthrex. He is also a consultant for Wright Medical, Stryker, and Smith & Nephew. Robert F. LaPrade, MD, PhD, is a consultant and receives royalties from Arthrex, Inc, Ossur, and Smith & Nephew.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.