
Abstract
In selective ulnar and median nerve transfers, donor nerve fascicles should be harvested in an area where motor and sensory fascicles are intermingled to minimize motor or sensory deficits. We aimed to define such an area for ulnar and median nerve harvesting through microanatomical dissection and histology in 12 fresh adult cadaveric upper extremities. Anatomically, we studied the arrangement, localization, and histological features of fascicle groups in two nerves at eight segments of the upper arms. Histological sections were examined to confirm the findings of the anatomical dissections. We found that sensory and motor fascicles were mixed proximally to the third most distal segment of the ulnar nerve and to the fourth most distal segment of the median nerve. We conclude that harvesting a part of the ulnar or median nerve proximal to these levels minimizes donor nerve deficits.
Introduction
Nerve transfer is one of the surgical treatment possibilities for brachial plexus injury. Its clinical prognosis depends on the donor–recipient nerve–fibre match, the long period required for nerve reinnervation, the mismatching of motor and sensory nerves, and the complexity of the donor nerve. Close attention should be paid to protecting residual function and reducing additional damage during surgery. Oberlin et al. (1994) reported ulnar nerve transfer to the biceps branch of the musculocutaneous nerve to restore elbow flexion, which is named Oberlin’s procedure. Later, the brachial triceps branch and the median nerve were transferred to the axillary nerve to restore shoulder abduction using modifications of Oberlin’s procedure (Leechavengvongs et al., 2003; Mackinnon et al., 2005). Transfer of the phrenic nerve or the motor branch of the accessory nerve to the suprascapular nerve has been used to restore shoulder abduction (Li and Gu, 2002). To reconstruct hand function, nerve transfer was used to restore intrinsic muscle function in the hand (Mackinnon, 2016). Furthermore, selective nerve transfers can be used in targeted muscle reinnervation to control prostheses (Takagi et al., 2019).
Harvesting nerves for transfer may cause functional deficits, and the localization of the harvested bundle may influence the prognosis. The purpose of this study was to investigate the optimal level for harvesting the ulnar and median nerves for transfer.
Methods
Microanatomical studies
Twelve formalin-fixed upper extremities were used in this study. The ulnar and median nerves and their branches were dissected. The ulnar and median nerves in the forearm (from the styloid process of the radius to the medial epicondyle of the humerus) were evenly divided into 16 segments (Figure 1). The most distal nerve segment of the forearm was marked as the 1/16 segment, and the most proximal nerve segment of the forearm was marked as the 16/16 segment. For the definition of the level, the 0/16 level meant the distal transverse section of the 1/16 segment of nerve, while the 1/16 level meant the proximal transverse section of the 1/16 segment of nerve. Similarly, nerves in the upper arm were evenly divided into eight segments from the medial epicondyle of the humerus to the inferior boarder of the teres major muscle. For example, the 3/8 level of the ulnar nerve in the upper arm meant the proximal transverse section of the 3/8 segment of the nerve. The arrangement, localization, and merging of fascicle groups of the ulnar and median nerves were recorded.
The marking of segments and levels. Both the ulnar and median nerves in the forearm (from the styloid process of the radius to the medial epicondyle of the humerus) were evenly divided into 16 segments. Nerves in the upper arm were evenly divided into eight segments from the medial epicondyle of the humerus to the inferior boarder of the teres major muscle.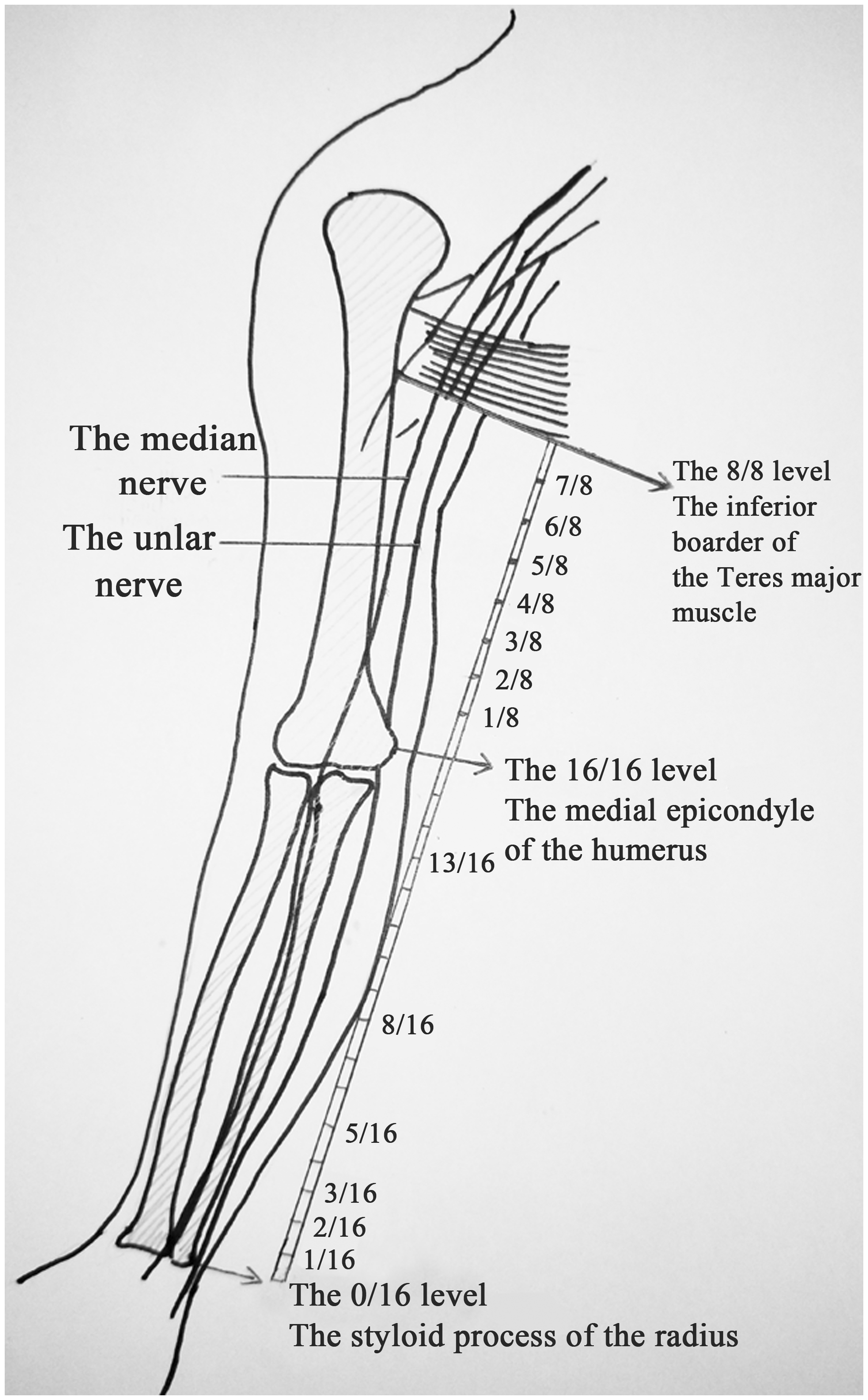
Histological studies
Twelve fresh adult cadaverous specimens were used. The ulnar and median nerves and their branches were exposed and divided according to the same standards as in the microanatomical studies.
All sections were stained using Karnovsky–Roots acetylcholinesterase histochemical stain to distinguish motor fibres from sensory fibres. A section of fresh nerve tissue specimen was placed in formalin (pH 7.4) prepared with phosphate buffer at 4℃ and fixed for 1 hour, then washed with phosphate buffered saline (PBS) at 4℃ (PBS, pH = 7.2) three to five times and stored in PBS overnight in refrigerator at 4℃. The nerve was embedded with optimal cutting temperature compound. After hardening of the embedding agent, the sections were cut into 15 -µm-thick cross-sectioned slices by the freezing slicer and incubated in the incubation solution at 4℃ for 24 hours. Then the excess sediment was washed off in 4℃ PBS. The slides were dried naturally and sealed with neutral gum. The incubation solution contained: 12.5 mg acetylthiocholine iodide, 16 ml 0.1 mol/L phosphate buffer, 1 ml 0.1 mol/L sodium citrate, 2.5 ml 30 mmol/L copper sulphate, 2.5 ml 5 mmol/L potassium ferricyanide, and 2 ml distilled water. For the negative control group, an incubation solution without iodized acetylthiocholine and 2 ml 10−4 mol/LEserin were used.
The distribution of functional fascicle groups in the transverse sections were examined under the microscope with 15 power and recorded in combination with the microdissection results. Another set of specimens were cut transversely into 7 -µm-thick sections and stained using solochrome cyanine. They were examined under the microscope with 200 and 400 power for counting fibres.
These histological sections were then analysed by image analysis software (Image-Pro Plus, Media Cybernetics, Rockville, MD, USA). Using Karnovsky–Roots acetylcholinesterase staining, the unmyelinated sensory nerve fibres exhibited a dark-brown strong positive staining with indistinct borders, while the myelinated motor nerve fibres, as well as axons, were moderately positive with clear borders. The staining of the myelin sheath was negative (Figure 2). Solochrome cyanine staining showed a clear sheath border of the myelinated nerve fibres in light blue, while the connective tissue stained red (Figure 3). For each histological section we calculated the fibre density and the number of myelinated fibres and studied the distribution of nerve branches and the merging of fascicle groups.
The acetylcholinesterase staining of the ulnar nerve in the 1/16 level of the forearm. The left part of the fascicular group was the deep branch to the hand, mainly including the motor fibres with moderate positive staining and clear border. The right part of the fascicular group was the superficial branch to the hand and mainly included the sensory fibres with dark-brown strong positive staining and indistinct border. The solochrome cyanine staining of the ulnar nerve in the 3/8 level of the upper arm. We used the magnified image for counting fibre numbers.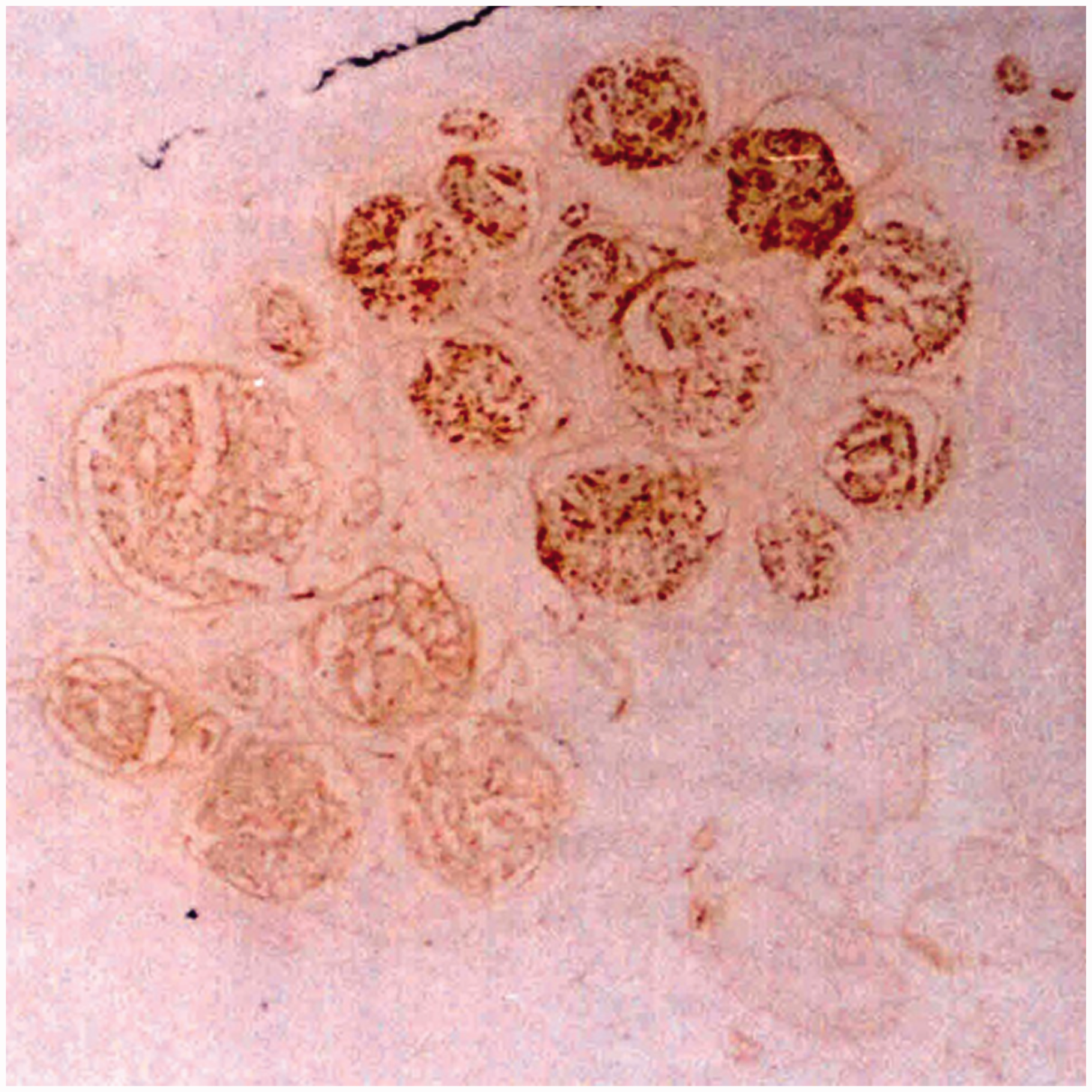

Statistical methods
The fibre density at different levels was compared using a one-way analysis of variance. The number of fibres in adjacent levels were compared using paired t-tests. The results are shown as the mean (standard deviation). A p-value < 0.05 was considered statistically significant.
Results
Distribution of the ulnar fascicle nerve
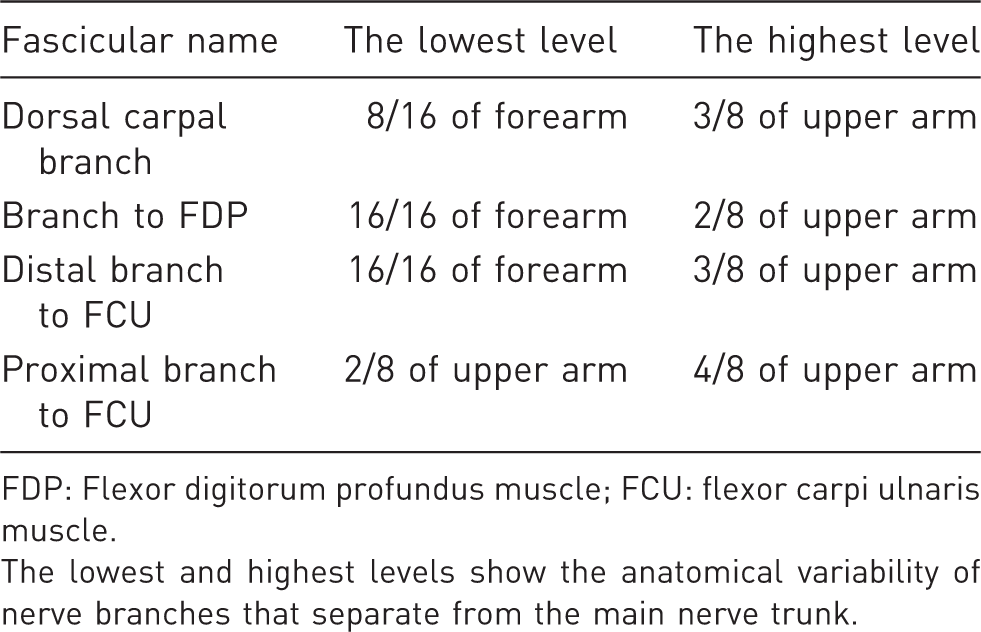
In the distal part of the forearm, the ulnar nerve consisted of three fascicle groups: the deep branch of the hand, the superficial branch of the hand, and the dorsal carpal branch. The deep and superficial branches of the hand composed the ulnar nerve trunk. The dorsal carpal branch accompanied the medial side of the nerve trunk and did not merge into it until the 15/16 level of the forearm. In the 10/16 to 12/16 levels of the forearm, the ulnar arterial fascicle merged into the trunk from the posterolateral side. In the 14/16 to 16/16 levels of the forearm, fascicles of the distal nerve branch to the flexor carpi ulnaris (FCU), flexor digitorum profundus, and proximal branch of the FCU gradually merged into the posteromedial side. The definite separate levels of several main nerve branches were marked (Table 1). It showed the variability of nerve branches separating from the main nerve trunk. Proximal to the 3/8 level of the upper arm, the histochemical staining showed no distinct border between the motor fibres and the sensory fibres. These fascicle groups showed characteristics of both sensory and motor fibres. These nerves were merged consistently (Figure 4).
The acetylcholinesterase staining of the ulnar nerve. The transverse section of the 3/8 level of the upper arm. The merged fascicles showed characteristics of both motor and sensory fibres. The lowest and highest levels at which the fascicles are definitely separated from the main nerve trunk. FDP: Flexor digitorum profundus muscle; FCU: flexor carpi ulnaris muscle. The lowest and highest levels show the anatomical variability of nerve branches that separate from the main nerve trunk.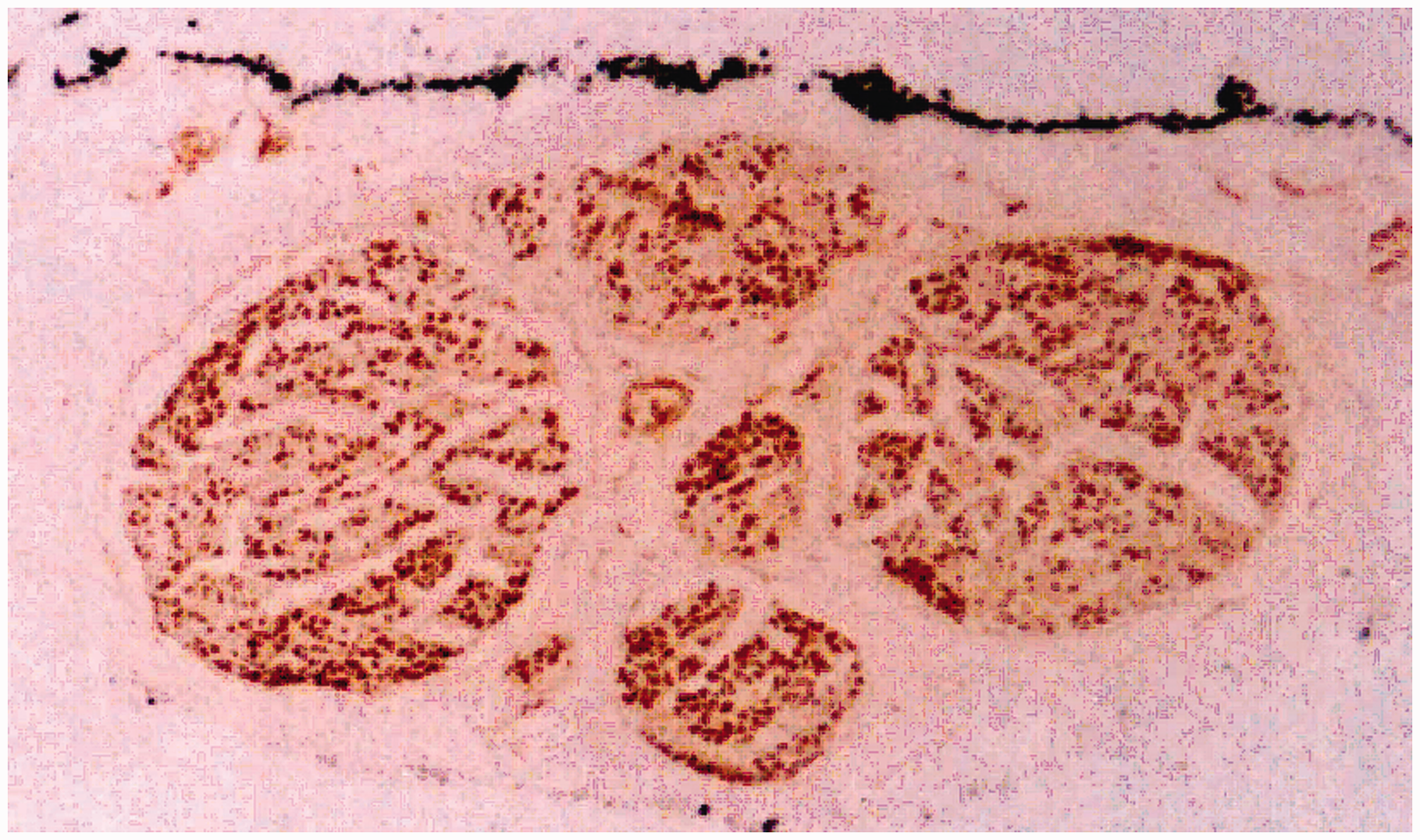
Distribution of the median fascicle nerve
The median nerve consisted of the intrinsic branch and palmer cutaneous branch of the hand in the distal part of the forearm. In the 11/16 to 14/16 levels of the forearm, the anterior interosseous nerve accompanied the posteromedial side of the nerve trunk. Merging into the trunk, the series of branches from the pronator quadrates, palmaris longus, flexor digitorum profundus, and superficialis and flexor pollicis longus varied individually. They could merge into the trunk separately or merge into each other and then into the trunk. These branches composed the posterior fascicle group around the 14/16 level of the median nerve. The branches of the flexor carpi radialis and pronator teres accompanied the lateral side of the trunk from the 11/16 to 14/16 levels. These branches gradually turned to the anterior side of the nerve trunk and composed the anterior fascicle. Proximal to the 1/8 level of the upper arm, there were three main fascicles with clear borders, including the posterior, anterior, and middle fascicles. In the 2/8 to 3/8 levels of the upper arm, the middle fascicle merged into the posterior fascicle. In the 3/8 to 4/8 levels of the upper arm, the anterior fascicle merged into the trunk. Proximal to the 4/8 level, the clear border of three distinct fascicle groups disappeared. Only a higher sensory fibre density could be observed in centre (Figure 5). Proximal to the 8/8 level, the median nerve divided into medial and lateral cords. The lateral cord had more motor fibres than the medial cord.
The acetylcholinesterase staining of the median nerve. The transverse section of 4/8 level of the upper arm. The merged fascicles showed characteristics of both motor and sensory fibres.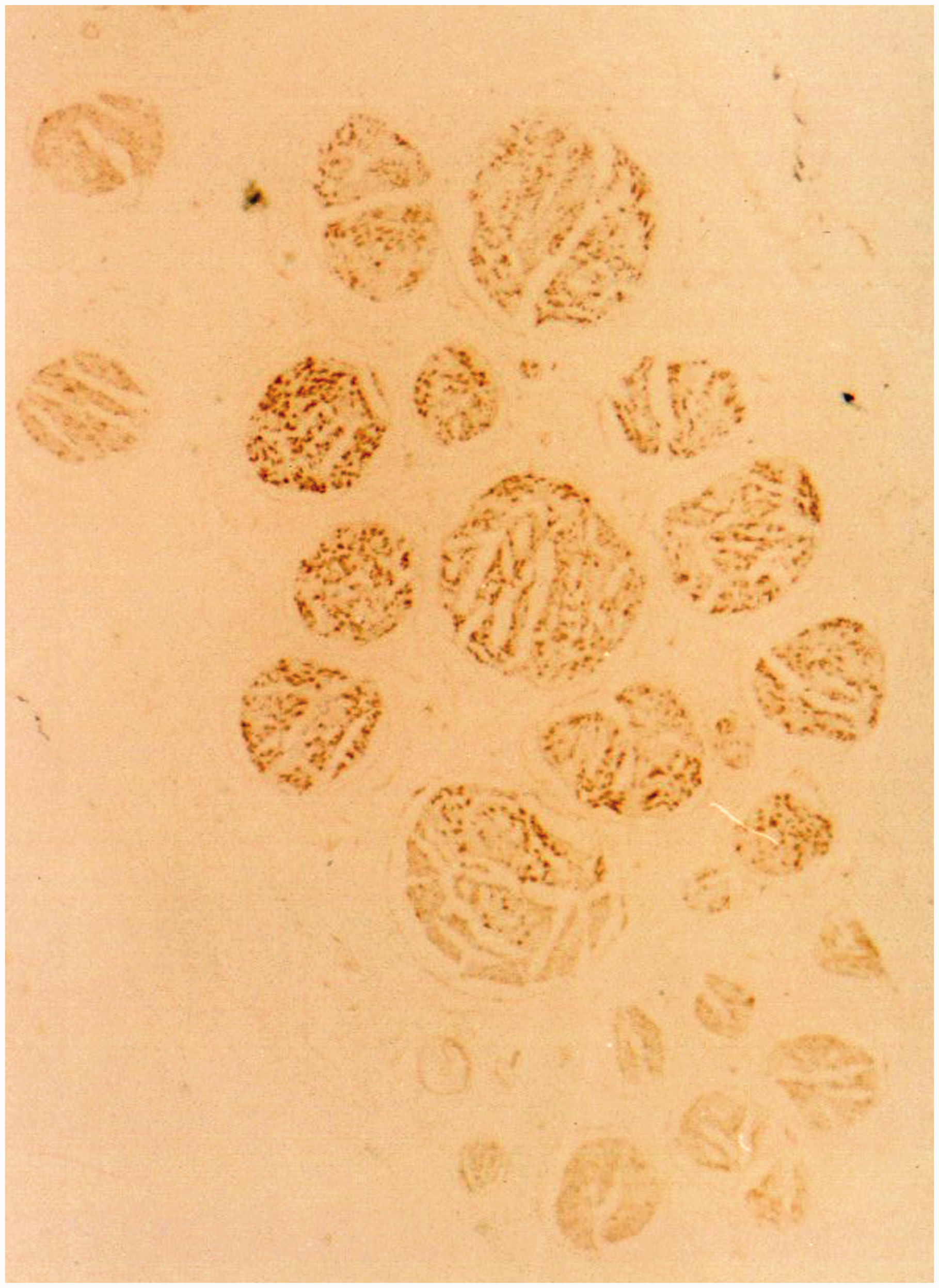
Staining results
Using solochrome cyanine staining, we determined the number of myelinated fibres in the ulnar and median nerves in the 8/8 level of the upper arm to be 14,234 (SD 1021) and 21,079 (SD 852), respectively (supplementary online Table S1). Several adjacent parts of the level indicated significant differences when compared using a paired t-test (p < 0.05). A one-way analysis of variance indicated no significant differences in fibre density between adjacent parts.
Discussion
We found that proximally to the 3/8 level of the ulnar nerve and the 4/8 level of the median nerve in the upper arm, sensory and motor nerve fibres were consistently merged. If donor nerve fibres are transferred proximal to these levels, motor dysfunction and paraesthesia of the donor nerve may be minimized. The 3/8 level of the ulnar nerve and the 4/8 level of the median nerve were localized at 80 mm (SD 1) and 107 mm (SD 1) proximal to the medial epicondyle of the humerus, respectively. These are the average data for adult Asian males and can be referenced for surgical planning.
Considering the functional compensation, transferring a small number of nerve fascicles from the intermingled fascicles would not lead to obvious donor nerve dysfunction (Sunderland, 1945). In most of the early published studies, the level at which the donor nerve was harvested was unclear. Oberlin merely recommended to make an 8–10 cm incision 4 cm distally to the lateral border of the pectoralis major muscle. It has been shown that the anteromedial branches to the intrinsic hand muscles may contribute to better innervation for active movement because of supply from a larger cerebral cortex area (Bertelli et al., 2004). However, this procedure may contribute to donor nerve deficits, which were transient or spontaneously recovered (Carlsen et al., 2011; Cho et al., 2014). The anterolateral branches of the FCU may cause less damage of the donor nerve (Suzuki et al., 2011). Paraesthesia or motor dysfunction of the donor nerve can last more than 3 months (Estrella, 2011; Nath et al., 2006). Therefore, it is important to choose a transfer level that can both reduce potential donor nerve dysfunction and restore function in the paralysed extremity.
We consider that if we harvested donor nerves with relatively consistent intermingling fascicles and sufficiently large fibres remaining to provide continued function, the dysfunction of the donor nerve could be minimized. Sections proximal to the 3/8 level of the ulnar nerve and the 4/8 level of the median nerve in the upper arm showed relatively consistent mixed motor and sensory nerve fascicles. In clinical application, the key point is that the donor nerve fascicle should be harvested in this area to avoid purely motor or sensory deficit will result from the nerve transfer. These areas represent the safe levels for these two donor nerves.
Limitations to our study include that the sample size was only 12 adult cadaverous specimens. Zhao et al. (2001) reported that definite separate levels of the anterior and posterior fascicle groups were located in the 4.8/8 to 6.6/8 levels and 2.6/8 to 3.9/8 levels of the upper arm, respectively, while our research showed that nine of 12 samples had definite separate levels of the anterior fascicle group in the 5/8 level and that eight of 12 samples had definite separate levels of the posterior fascicle group in the 3/8 level. Different results may come from different choices of definite separate points and sample size.
Supplemental Material
JHS901441 Supplemental Table - Supplemental material for Safe level for harvesting for ulnar and median nerve transfers: a microanatomical and histological study
Supplemental material, JHS901441 Supplemental Table for Safe level for harvesting for ulnar and median nerve transfers: a microanatomical and histological study by Wen-ting He, Shao-guang Li, Yan Shao, Jian-tao Yang, Yi Yang, Ben-gang Qin and Li-qiang Gu in Journal of Hand Surgery (European Volume)
Footnotes
Acknowledgement
Wen-ting He and Shao-guang Li contributed equally to this study.
Declaration of conflicting interests
The authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: we acknowledge the support to this research work from the National Nature Science Foundation of China (grant numbers 81572130, 81601057) and the Natural Science Foundation of Guangdong Province (grant numbers 2018A030310254).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.